
Abstract
Abstract
Purpose:
To explore the inhibitory effects of antivascular endothelial growth factor C (VEGF-C) therapy on corneal lymphangiogenesis and allograft rejection in rats.
Methods:
Fischer 344 rat corneas were transplanted into Lewis rat eyes. After corneal transplantation, Lewis rats (the recipients) were randomly and equally divided into 2 groups: anti-VEGF-C treatment (group A) and control (group B). Corneal hemangiogenesis and lymphangiogenesis were characterized using whole-mount immunofluorescence, and the immune rejection of the grafts was examined using a slit lamp and evaluated by scoring the rejection index (RI). In addition, the expression of VEGF-C was examined by immunohistochemistry and real-time polymerase chain reaction. The association of corneal lymphangiogenesis and hemangiogenesis with VEGF-C in transplanted corneas was also characterized.
Results:
VEGF-C expression was markedly downregulated after anti-VEGF-C therapy. The outgrowth of corneal lymphangiogenesis dramatically decreased in group A. There was a significant relationship between VEGF-C reduction and the decrease in the lymphatic vessel area (r=0.55, P<0.05), whereas the relationship between the reduction of VEGF-C and the decrease in blood vessel area was not significant (r=0.11, P>0.05). In addition, the RI scores were significantly lower in group A compared with group B at 7, 10, and 14 days after transplantation. The graft survival time in group A rats (20.33±1.37 days) was significantly longer than that in group B rats (12.83±1.47 days; P<0.05).
Conclusions:
The results suggested that VEGF-C blockade had a significant role in preventing corneal lymphangiogenesis in corneal beds, which resulted in higher allograft survival rates.
Introduction
T
Although additional unknown factors may be involved in the establishment and maintenance of the lymphatic vasculature, vascular endothelial growth factor C (VEGF-C) has been described as a predominantly lymphangiogenic factor.7,8 Direct evidence for the role of VEGF-C in promoting lymphangiogenesis comes from studies of transgenic mice overexpressing VEGF-C under the control of the keratin14 (k14) promoter. K14-VEGF-C mice displayed pronounced hyperplasia of the cutaneous lymphatic vessels, whereas blood vessel growth was not affected. 9 Conversely, lymphatic vessels transiently regressed in the skin of the inner organ of k14-VEGFR3-Ig mice. 10 In a previous study, we examined transplanted rat corneas and found that VEGF-C expression dramatically increased after keratoplasty. 11 Our results also revealed a significant relationship between corneal lymphangiogenesis and VEGF-C mRNA levels, indicating that corneal lymphangiogenesis correlated closely with VEGF-C expression after keratoplasty. Therefore, targeting VEGF-C might be useful to control the development of corneal lymphangiogenesis and inhibit later allograft failure.
Based on these findings, the present study targeted VEGF-C in transplanted corneas through the intraperitoneal injection of the monoclonal anti-VEGF-C antibody and investigated whether anti-VEGF-C therapy had an inhibitory role in corneal lymphangiogenesis and allograft rejection. Together, these findings may broaden our understanding of the mechanisms involved in corneal lymphangiogenesis and corneal transplant rejection.
Methods
Animals
We used 132 of 139 Lewis rats (RT1l) as recipients, with the remaining 7 rats serving as normal controls. In addition, 66 Fischer 344 rats (RT1lv1) were used as donors for corneal transplantation. All rats were male and aged 8–10 weeks, with weights ranging from 180 to 200 g. All rats were provided by the Animal Center of Sun Yat-Sen University, Guangdong, China. The experimental conditions used in the study conformed to good laboratory practices (National Research Council, 1996), and all animals involved in the study were handled in accordance with the ARVO Statement for the Use of Animals in Ophthalmic and Vision Research. Before treatment, we confirmed that all rats were free of ocular disease. The experimental protocol was approved by the Sun Yat-Sen University/Institutional Animal Care and Use Committee. All the surgeries were performed under chloral hydrate anesthesia, and all efforts were made to minimize suffering.
Orthotopic corneal transplantation and VEGF-C blockade
The procedure for corneal transplantation was adapted from the technique described previously by Liu et al. 12 Briefly, rats were anesthetized with chloral hydrate (300 mg/kg) through the abdominal cavity, and 1% atropine was administered 20 min before the operation. Donor corneas (3.5 mm in diameter) were removed and placed in Optisol solution. After the recipient's right cornea was marked with a 3.0-mm trephine and excised under the operating microscope, the donor graft was sutured into the recipient's bed with 8 interrupted sutures (10-0 nylon). The anterior chamber of the eye was reformed by injecting a balanced salt solution. The graft sutures were removed 2 weeks after the operation.
After corneal transplantation, the recipient rats were randomly and equally divided into 2 groups: anti-VEGF-C treatment (group A) and control (group B). To block VEGF-C, 20 mg/kg (in a total 0.1 mL volume) of neutralizing goat anti-rat VEGF-C monoclonal antibody (Boshide Bio-Project Co., Wuhan, China) was injected intraperitoneally the day after surgery and subsequently on alternate days for up to 2 weeks. Control rats received 0.1 mL of normal saline, intraperitoneally.
In each group, 5 corneas (the right cornea in Lewis rats, including both donor and recipient parts) were examined by immunohistochemistry, real-time polymerase chain reaction (PCR), and whole-mount immunofluorescence at 3, 7, 10, and 14 days after transplantation; 6 rats were used for slit-lamp observation to evaluate the state of corneal graft rejection. In addition, 7 right corneas from normal Lewis rats were used as normal controls for immunohistochemistry (2 corneas), immunofluorescence (2 corneas), and real-time PCR (3 corneas).
Immunohistochemistry
Corneas were fixed in 10% neutral formalin for 24 h, embedded in paraffin, serially sectioned into 4-μm sections, rehydrated using a graded ethanol/water series, and washed with distilled water. The endogenous peroxidase activity was blocked by incubating in 30 mg/L hydrogen peroxide for 20 min. Tissue sections were then autoclaved at 121°C in a 10 mM citrate buffer (pH 6.0) for 10 min to retrieve antigens and then cooled at room temperature for 30 min. Corneal sections were then incubated with the goat anti-rat VEGF-C monoclonal antibody (Boshide Bio-Project Co.) for 3 h, followed by biotin-conjugated rabbit anti-goat immunoglobulin as the secondary antibody. The streptavidin–biotin complex peroxidase was used as the immune check system. The slides were visualized using diaminobenzidine and counterstained with hematoxylin.
For VEGF-C staining, the brown cells were considered VEGF-C positive. VEGF-C expression was determined according to Zhang et al. 13 with modifications. Briefly, the immunostained sections were scanned on a light microscope, and all of the cells were evaluated using the following criteria: −, negative; +, focal expression in <5% of the cells; ++, focal expression in 5%–20% of the cells; and +++, diffuse expression in >20% of the cells. Tissue with +++ staining for VEGF-C was classified as “high,” tissue with ++staining was classified as “middle,” and tissues with − and + staining were classified as “low.”
RNA isolation and purification
Total RNA was isolated from the samples using the TRIzol Reagent (GIBCO-BRL Life Technologies, Gaithersburg, MD). RNA was prepared according to the manufacturer's protocol. The RNA pellets were washed with 75% ethanol, centrifuged, and dried. Residual DNA was removed by treatment with DNase I. Pellets were resuspended in 30 μL of diethylpyrocarbonate-treated water followed by the addition of 50 mM Tris, pH 7.5, 10 mM MgCl2, 20 U of RNase-free DNase I, and 20 U of RNase in a total volume of 60 μL. Samples were incubated at 37°C for 25 min. The RNA was then cleaned using RNeasy Mini Kits (Qiagen, Valencia, CA) following the manufacturer's protocol. RNA concentration and purity were determined by measuring the optical density at 260 and 280 nm on a spectrophotometer.
Real-time reverse transcription-polymerase chain reaction
cDNA was generated from total RNA samples using the TaqMan Reverse Transcription Reagents Kit (Applied Biosystems, Foster City, CA). To generate cDNA, total RNA from each sample was first incubated at 25°C for 10 min and then reverse transcribed at 48°C for 30 min. Real-time reverse transcription-polymerase chain reaction (RT-PCR) was performed using SYBR green dye (Applied Biosystems) with the ABI PRISM 7900HT instrument. The VEGF-C primers were 5′-TGTAAAACTTGCTGCTGCACATT-3′ (sense) and 5′-GAACGTCTAATAATTGAATGAACTTGTCT-3′ (antisense) (GenBank accession No. AY032729). The VEGFR2 primers were 5′-ATCGGTGAGAAAGCCTTGATCTC-3′ (sense) and 5′-TTCTAGCTGCCAGTACCATTGGA-3′ (antisense) (GenBank accession No. NM_013062). The VEGFR3 primers were 5′-CTGAGGCAGAATATCAGTCTGGAG-3′ (sense) and 5′-AGATGCTCATACGTGTAGTTGTCC-3′ (antisense) (GenBank accession No. NM_053652.1). DNA polymerase was first activated at 95°C for 10 min, then denatured at 95°C for 15 s, and finally annealed/extended at 60°C for 1 min for 40 cycles, according to the manufacturer's protocol. The products were sequenced to ensure that the correct gene sequence was being amplified. All PCRs were performed in triplicate. Relative quantitation of gene expression was performed using the standard curve method (User Bulletin number 2, ABI PRISM 7700 Sequence Detection System; Applied Biosystems). To compare the transcript levels between samples, standard curves were prepared for both the target gene and the endogenous reference (18S ribosomal RNA). For each experimental sample, the amounts of transcript for the target and endogenous reference were determined from the appropriate standard curves. Then, the target amount was divided by the endogenous reference amount to obtain a normalized target value. Each of the experimentally normalized sample values was divided by the normalized control sample value to generate the relative expression levels.
Immunofluorescence of whole-mount rat corneas
The method was modified from a previous protocol.14,15 Briefly, 10 mL of fluorescein isothiocyanate (FITC)-lectin from Lycopersicon esculentum (Sigma-Aldrich, St. Louis, MO) (20 μg/mL) was injected intracardially and allowed to perfuse for 2 min. The cornea was incubated in phycoerythrin (PE)-labeled anti-rat PECAM1 (CD31-PECAM) (BD Biosciences, San Diego, CA) (1:50) in the TNB blocking buffer (TSA Biotin System; NEN Life Science, Boston, MA) overnight at 4°C after being dissected and fixed in acetone for 15 min at −20°C. Corneas were digitally photographed on a confocal fluorescent microscope (Zeiss LSM 510 Meta; Carl Zeiss, Jena, Germany). The lymphatic vessel area (LVA) was defined as the total lectin−/PECAM+ area, and the blood vessel area (BVA) was defined as the total lectin+/PECAM+ area. The images were further analyzed at low magnification (×2), and the LVA and BVA were quantified using the AxioVision 4.7.2 (Carl Zeiss) digital image analysis software.
Slit-lamp microscopy
The corneal grafts were examined by a blinded observer every day following transplantation. The clinical appearance of each graft was scored according to the protocol established by Holland et al. 16 Briefly, a rejection index (RI) of 0–12 was generated based on the sum of 3 graft components: clarity, edema, and neovascularization (NV) (0–4 for each component). The following clarity scoring system was used: 0, clear cornea; 1, slight haze; 2, increased haze, but anterior chamber structures still clear; 3, advanced haze with a difficult view of the anterior chamber; and 4, opaque cornea with no view of the anterior chamber. The following edema scoring system was used: 0, no stromal or epithelial edema; 1, slight stromal thickness; 2, diffuse stromal edema; 3, diffuse stromal edema with microcystic edema of the epithelium; and 4, bullous keratopathy. NV was scored as follows: 0, no NV; 1, NV of the peripheral cornea; 2, NV of the corneal wound; 3, NV of the peripheral graft; and 4, NV of the entire graft. According to Zhang et al. 17 a clarity score of ≥3.0 and a total score>6 were recorded as rejected.
Statistical analysis
Graft survival was analyzed using the Kaplan–Meier survival method. 18 The Mantel–Cox log-rank test was used to determine significant differences between the groups. Significant differences between groups were analyzed using a paired t-test (SPSS 16.0 statistical software; SPSS, Inc., Chicago, IL). Pearson's analysis was used to determine correlations among BVA, LVA, and VEGF-C mRNA. Values are presented as the mean±standard deviation. Differences were accepted as significant at P<0.05.
Results
Intraperitoneal injection of monoclonal anti-VEGF-C antibody significantly reduced the expression of VEGF-C after keratoplasty
To investigate whether the intraperitoneal injection of a neutralizing monoclonal antibody played a role in VEGF-C blockade, we examined the expression of VEGF-C by immunohistochemistry and real-time PCR at different time points after keratoplasty. Immunohistochemical analysis demonstrated that VEGF-C proteins were absent in normal corneal stromas, but were expressed to some extent after transplantation. In group B, VEGF-C immunoreactivity dramatically increased on day 3, decreased gradually until day 7, and then peaked on day 14, with the majority of VEGF-C proteins localizing to invading inflammatory cells and vessel surroundings. However, group A displayed only mild immunoreactivity to VEGF-C on day 3 after transplantation. The expression of VEGF-C was faint on both days 7 and 14 after transplantation (Fig. 1).
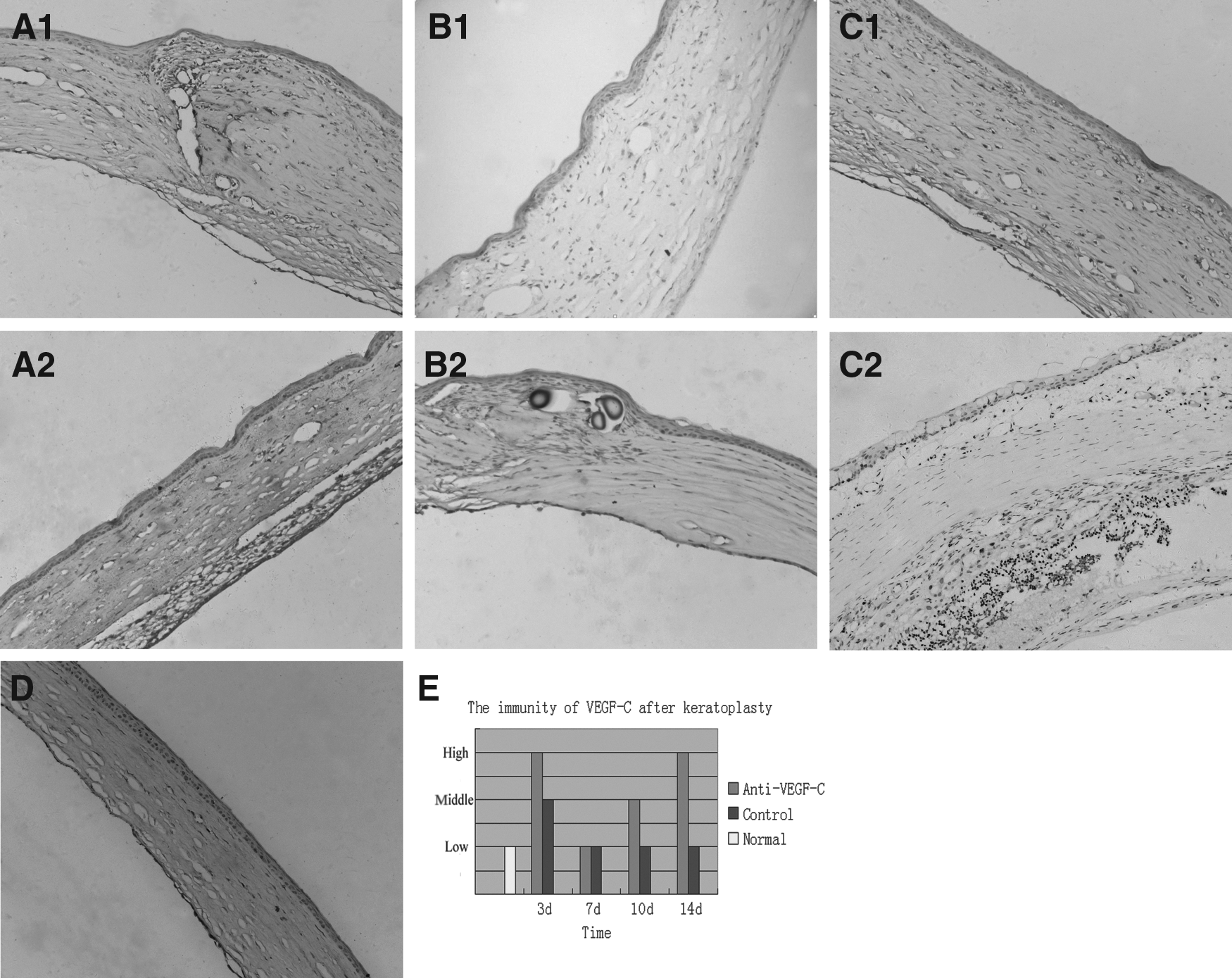
Immunohistochemistry of vascular endothelial growth factor C (VEGF-C) in transplanted rat corneas. In the control group, a large number of VEGF-C+ cells were observed invading the stroma on day 3 after transplantation
Real-time PCR revealed similar trends in VEGF-C expression between groups A and B after corneal transplantation (Fig. 2A, B). In both groups, VEGF-C mRNA levels were the highest on day 14 and the lowest on day 7 after keratoplasty. However, compared with group B, VEGF-C mRNA expression was significantly decreased at 3, 10, and 14 days after corneal transplantation in group A (Table 1). In addition, group A exhibited decreased VEGFR2 (0.407-fold decrease on day 3 compared with group B, P<0.05) and VEGFR3 (0.683-fold decrease on day 3, 0.615-fold decrease on day 10, and 0.708-fold decrease on day 14) expression compared with the control group. The downregulation on all days, but day 7, was significant (P<0.05) after anti-VEGF-C antibody injection (Fig. 3). We therefore concluded that the expression of VEGF-C was successfully inhibited following injection of the neutralizing monoclonal antibody.

The BVA and LVA trends were in parallel in the control group

The levels of VEGF-C, VEGFR2, and VEGFR3 mRNA after keratoplasty in each group. VEGF-C and VEGFR3 expression levels were markedly decreased in the corneas of anti-VEGF-C-group rats compared with the control group rat corneas on days 3, 10, and 14 after keratoplasty. VEGFR2 expression was only mildly reduced on day 3 after keratoplasty in the anti-VEGF-C group. The difference in VEGFR2 expression was not significant between the 2 groups on days 7, 10, and 14 after keratoplasty. Data are expressed as the fold changes compared with normal nontransplanted corneas. Graphs represent the mean±SEM (standard error of the mean) of 5 samples per group (*P<0.05 vs. control group).
Data are expressed as the mean±standard deviation.
P<0.05, compared with group B.
P>0.05, compared with group B.
VEGF-C, vascular endothelial growth factor C.
VEGF-C blockade effectively inhibited corneal lymphangiogenesis after keratoplasty
To visualize corneal lymphatic and blood vessels, we labeled blood vessels green through intravenous injection of FITC-conjugated lectin and labeled all vessels red by incubating the whole cornea with PE-conjugated anti-CD31. After the green and red fluorescent images were merged, blood vessels appeared yellow and lymphatic vessels appeared red. Using this method, we observed that the growth of new corneal blood vessels and lymphatic vessels was much faster in group B rats than in group A rats (Fig. 4A). The BVA was 3.3±0.42, 9.86±0.97, 12.44±1.29, and 14.72±0.98 mm2 at 3, 7, 10, and 14 days, respectively, after keratoplasty in group B, whereas the BVA in group A was 2.98±0.33, 8.56±0.58, 11.58±0.77, and 11.96±1.27 mm2, respectively. The difference in BVA between groups A and B was significant on days 7 and 14 after keratoplasty (both P<0.05). Compared with the BVA, the LVA was lower in group A. Whereas the LVA was 0.52±0.13, 2.34±0.34, 4.04±0.29, and 5.40±0.91 mm2 at 3, 7, 10, and 14 days, respectively, after keratoplasty in group B, it was 0.22±0.05, 0.94±0.17, 1.42±0.35, and 1.28±0.33 mm2 at the same time points in group A. The difference in LVA between the 2 groups was significant at all time points after keratoplasty (Fig. 3b) (all P<0.05). Although there was no significant relationship between the decrease in BVA and the reduction in VEGF-C expression (r=0.11, P>0.05), the relationship between the decreased LVA and the reduction in VEGF-C expression was significant (r=0.55, P<0.05) (Fig. 4C, D), suggesting that anti-VEGF-C therapy effectively inhibited corneal lymphangiogenesis after keratoplasty.
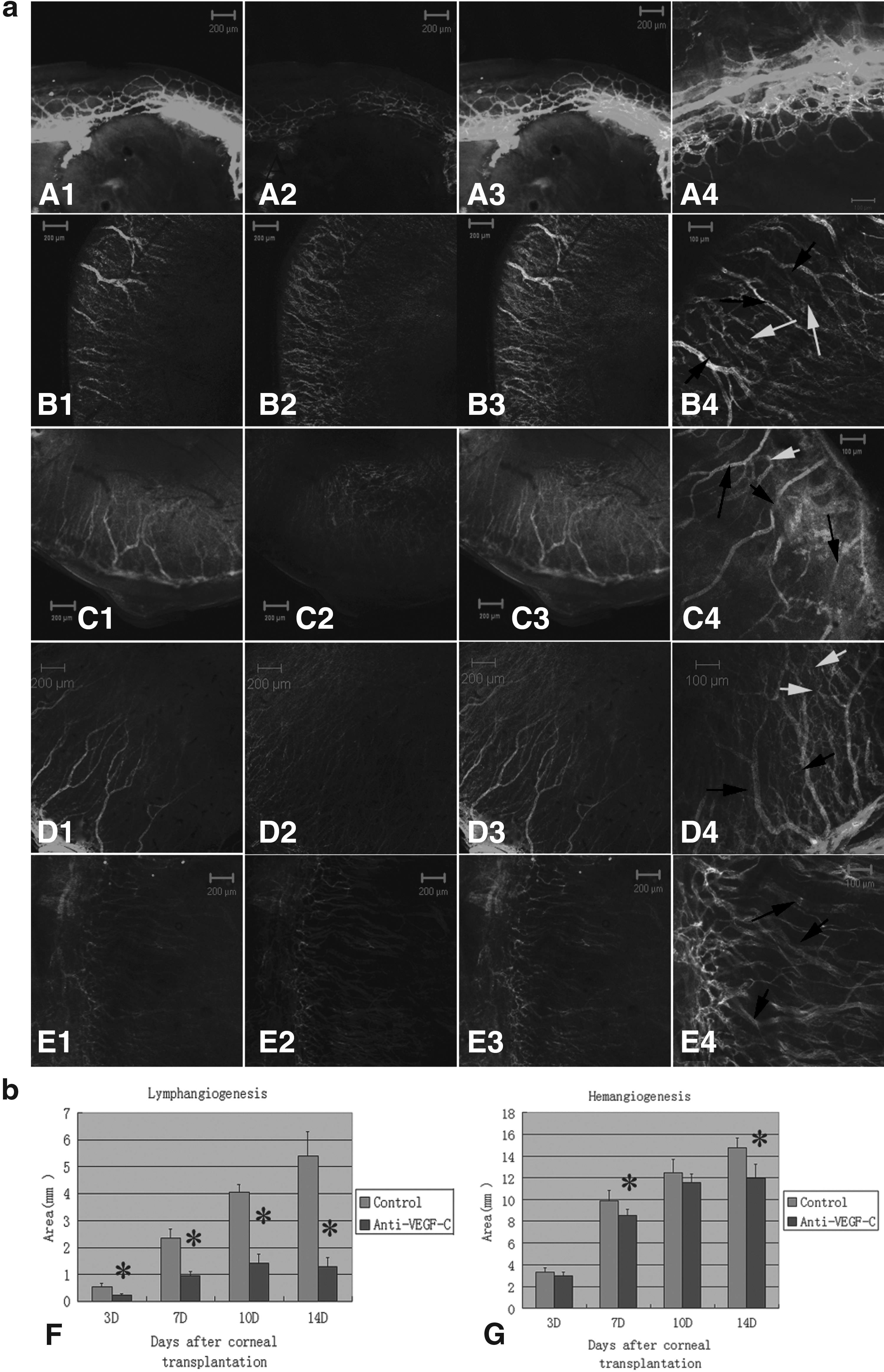
Whole-mount corneal immunofluorescence
Anti-VEGF-C therapy improved corneal graft survival
Although corneal NV (hemangiogenesis and lymphangiogenesis) was present in both group A and group B grafts, the growth of blood vessels in group B was more rapid than in group A on day 7 after corneal transplantation (Fig. 5A, C). On day 14 after transplantation, most grafts in group B failed, with abundant blood and lymphatic vessels invading into the center of the cornea. However, corneal NV occurred only at the edges of the grafts in group A, and most of the grafts survived (Fig. 5B, D). The RI and NV scores in group A were significantly lower than in group B at 7, 10, and 14 days after transplantation (Table 2). Compared with the graft survival time for group B (12.83±1.47 days), the graft survival time for group A (20.33±1.37 days) was significantly higher (P<0.05, Fig. 6). Together, these results strongly suggested that intraperitoneal injection of a monoclonal anti-VEGF-C antibody was effective in preventing corneal allograft rejection.
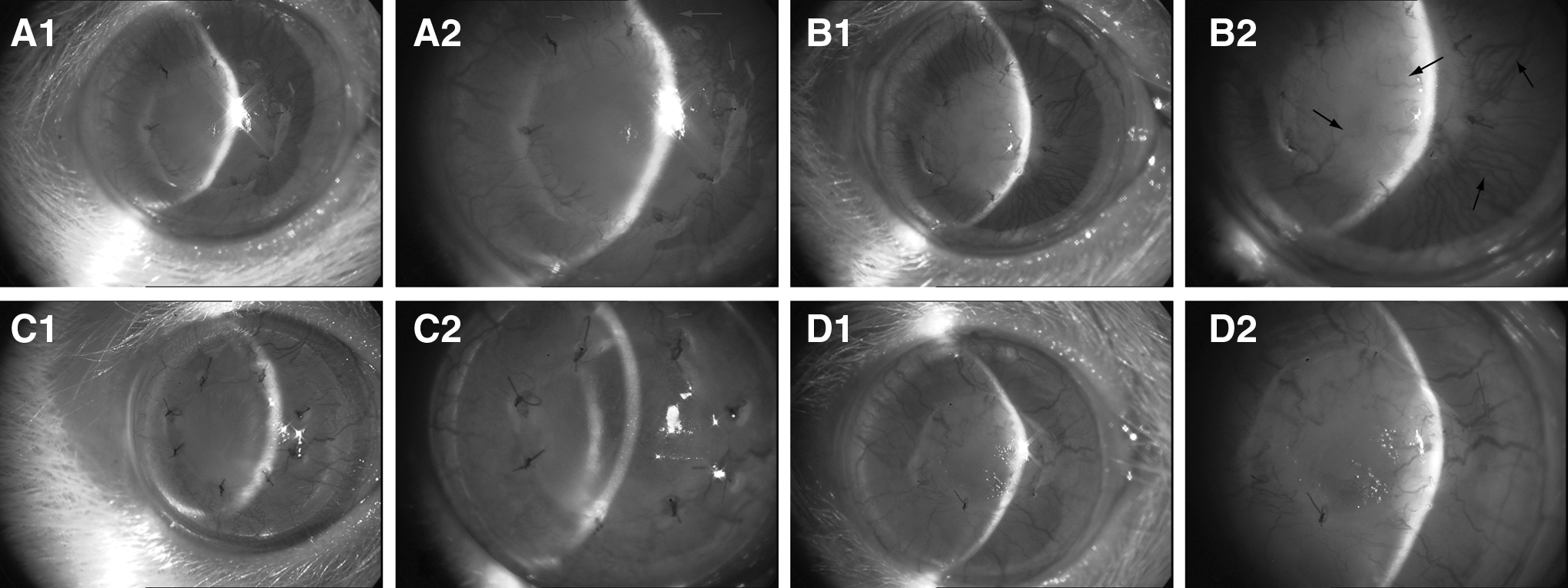
Slit-lamp microscopy after corneal transplantation. Corneal neovascularization (hemangiogenesis and lymphangiogenesis), appearing as a hairbrush from the limbus to the graft, developed at day 7
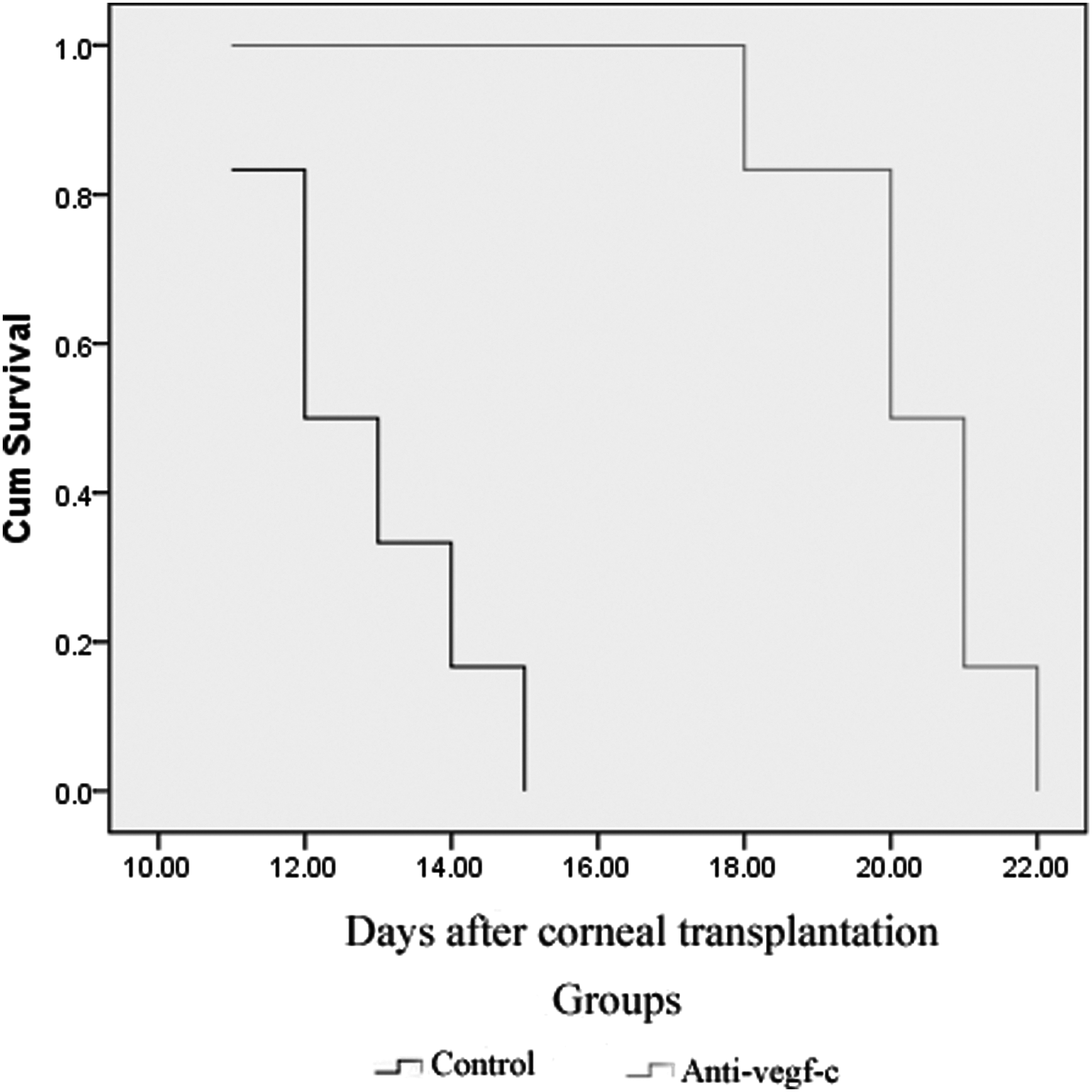
Survival curve of the grafts for the 2 groups. The recipients in the control group showed accelerated rejection. The median survival was significantly different between the 2 groups (P<0.05).
Data are expressed as the mean±standard deviation.
P>0.05, compared with group B
P<0.05, compared with group B.
RI, rejection index.
Discussion
We previously examined the development of corneal lymphangiogenesis, hemangiogenesis, and inflammation after alkaline burns and then performed keratoplasty in alkali-burned corneal beds, according to the state of corneal hemangiogenesis, lymphangiogenesis, and inflammation, to compare their respective roles in allograft rejection. 6 The results showed that corneal allograft immunity decreased with the disappearance of corneal lymphatic vessels, regardless of whether corneal hemangiogenesis and inflammation were present, suggesting that corneal lymphangiogenesis played a more important role than the other 2 factors in allograft rejection after alkali burns. A similar study performed by Dietrich et al. 5 also showed that corneal lymphangiogenesis played a critical role in allograft failure. Due to the important role of VEGF-C in corneal lymphangiogenesis, we hypothesized that blocking VEGF-C might be useful in promoting corneal allograft survival by selectively inhibiting the outgrowth of corneal lymphatic vessels after keratoplasty. However, until recently, there have been few reports of anti-VEGF-C therapy during corneal lymphangiogenesis or corneal transplantation.
To determine the effects of anti-VEGF-C therapy on corneal lymphangiogenesis after keratoplasty, we first examined corneal lymphangiogenesis, hemangiogenesis, and VEGF-C mRNA levels at different time points in groups A and B. We then examined the relationship between the decrease in LVA and BVA and the reduction in VEGF-C levels. We characterized the development of corneal lymphangiogenesis, hemangiogenesis, and VEGF-C and analyzed the inhibitory role of VEGF-C blockade in corneal lymphangiogenesis and hemangiogenesis. In a recent study similar to ours, Emami-Naeini et al. 19 also examined corneal lymphangiogenesis in transplanted corneas after VEGF-C blockade using soluble VEGFR3 and found that VEGF-C blockade had a role in inhibiting corneal lymphatic vessels and promoting allograft survival. However, this study only examined corneal lymphatic and blood vessels on day 14 after treatment. In our study, we examined corneal lymphangiogenesis and hemangiogenesis at 3, 7, 10, and 14 days after corneal transplantation to better characterize the changes to and development of both blood and lymphatic vessels. Moreover, by comparing VEGF-C mRNA levels at different time points, we were able to determine whether anti-VEGF-C therapy inhibited VEGF-C expression and efficiently decreased corneal lymphangiogenesis or hemangiogenesis in transplanted corneas.
We also examined the effect of VEGF-C blockade on allograft rejection. Chen et al. showed that blockade of VEGFR3, one of the most important receptors for VEGF-C, significantly suppressed corneal APCs trafficking to draining lymph nodes, the induction of delayed-type hypersensitivity, and the rejection of corneal transplants. 20 Recently, both Emami-Naeini et al. and Hajrasouliha et al. reported that targeting VEGF-C inhibited the trafficking and maturation of APCs and improved allograft survival.19,21 In the present study, we focused on the antilymphangiogenic role of VEGF-C blockade and characterized the effect of this role on allograft survival. Based on our previous finding that most recipient rats in similar conditions began to show allograft rejection on day 14, we examined changes in the RI, lymphangiogenesis, and hemangiogenesis within 14 days after transplantation. 22 We found that although there was a significant difference in RI between groups A and B, the clarity and edema scores between the 2 groups were not significantly different. However, NV (blood and lymphatic vessels) scores in group A were dramatically lower at 7, 10, and 14 days after transplantation, which might be responsible for the significant difference in RI between the 2 groups. Taken together with our previous finding of a parallel relationship between RI and lymphangiogenesis from 7 to 14 days after corneal transplantation, 23 we concluded that corneal lymphatic vessels played an important role in allograft rejection and that the improved graft survival was closely associated with the reduction in corneal lymphangiogenesis after VEGF-C blockade.
Recently, Goyal et al. reported a significant reduction in lymphatic, but not blood vessel, ingrowth upon VEGF-C blockade in dry eye disease. 24 However, in the present study, we found that both corneal lymphatic and blood vessels were inhibited by VEGF-C blockade after keratoplasty. The results were not consistent with the results of Goyal et al. or with our hypothesis that anti-VEGF-C therapy would selectively inhibit corneal lymphangiogenesis. In addition to inducing lymphangiogenesis through the VEGFR3 pathway, VEGF-C has also been reported to act by binding VEGFR2.7,25 Whereas the 31-kDa form of VEGF-C only bound to VEGFR3 and was therefore exclusively lymphangiogenic, the 21-kDa form also induced hemangiogenesis by binding to VEGFR2.26, 27 Therefore, we also examined the expression of VEGFR3 and VEGFR2 in transplanted corneas by real-time PCR. Our data showed that the expression of both VEGFR3 and VEGFR2 decreased to some extent after anti-VEGF-C treatment. However, compared to the VEGFR2 transcript levels, the VEGFR3 levels were much lower. The experimental group treated with the anti-VEGF-C antibody showed a significant reduction in VEGFR3 compared with the control group, whereas VEGFR2 expression was only mildly reduced (except for day 3 after keratoplasty) in the anti-VEGF-C group. Together with our finding that group A rat corneas showed a dramatic reduction in LVA, but only a mild decrease in BVA, we hypothesize that the majority of VEGF-C interacted with VEGFR3 to induce lymphangiogenesis and that the VEGF-C blockade had a crucial role in inhibiting the outgrowth of lymphatic vessels, but not blood vessels, in transplanted corneas.
In our study, VEGF-C was blocked through the intraperitoneal injection of a neutralizing monoclonal anti-VEGF-C antibody, which was previously used in angiogenesis and transplant models.19,24 However, although anti-VEGF-C treatment can significantly decrease VEGF-C transcript levels, it also inhibits VEGF-D. VEGF-D is another important factor in the induction of corneal lymphangiogenesis. 24 Therefore, in addition to VEGF-C blockade, corneal lymphangiogenesis might also be reduced by VEGF-D downregulation during anti-VEGF-C antibody therapy. Furthermore, although corneal lymphangiogenesis and allograft failure were effectively inhibited by anti-VEGF-C antibody treatment, all grafts were rejected within 1 month, suggesting that the VEGF-C-lymphangiogenesis axis plays only a partial role in allograft rejection. Combined treatment with an immunosuppressant, for example, would be valuable in future studies.
In summary, our study characterized the development of corneal lymphangiogenesis and hemangiogenesis rejection scores in transplanted rat corneas after VEGF-C blockade and provided evidence that anti-VEGF-C treatment helped to inhibit corneal lymphangiogenesis and promote allograft survival. In future studies, these results may form the basis for novel treatments that target the lymphatic system after corneal transplantation.
Footnotes
Acknowledgments
This study was supported by the China Natural Science Foundation (81070711), the Guangdong Natural Science Foundation (S2013010016324), the Guangdong Provincial Science and Technology Projects (2013B060300003), and the Young Teachers Training Program of Sun Yat-Sen University (11ykpy42). The authors thank Dr. Chaoyang Li and Dr. Chuangchao Xu for their invaluable technical support.
Author Disclosure Statement
No competing financial interests exist.