
Abstract
Abstract
Purpose:
Repeated subconjunctival injections with 5-fluorouracil (5-FU) after trabeculectomy are used in glaucoma patients for the inhibition of overproliferation in wound site. Thus, a certain amount of the drug may penetrate into epithelial layer, where it causes toxicity to corneal epithelial cells. The aim of this study was to evaluate the toxic effects of 5-FU and mechanisms of drug-induced apoptosis in cultured corneal epithelial cells.
Methods:
Cellular damage and the caspase pathway were estimated with a 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) assay. The apoptotic characteristics were detected by flow cytometry, a TUNEL test, and western blotting in cultured corneal epithelial cells.
Results:
The results indicated that 5-FU was toxic to corneal epithelial cells in a time- and dose-dependent manner. Pretreatment with a general caspase inhibitor (Z-VAD-FMK), a caspase-8 inhibitor (Z-IETD-FMK), and a caspase-9 inhibitor (Z-LEHD-FMK) reversed 5-FU-induced cellular damage. Following exposure to 5-FU, a flow cytometric assay with MitoLight dye demonstrated the significant loss of mitochondrial membrane potential. A positive TUNEL test revealed that cellular DNA apoptosis occurred following exposure to 0.5, 1, and 5 mg/mL 5-FU for 15 h. Positive annexin V–FITC and negative propidium iodide (PI) staining indicated that the cell membrane exhibited apoptosis upon exposure to 1 and 5 mg/mL 5-FU for 15 h. The western blot assay demonstrated upregulation of the p21 protein but downregulation of the Bcl-2 proteins induced by 5-FU.
Conclusion:
These data reveal that 5-FU-induced cellular apoptosis in corneal epithelial cells may be mediated through caspase-8, caspase-9, and mitochondria-regulated pathways, as well as by upregulation of p21 and downregulation of Bcl-2-dependent signal transduction pathways.
Introduction
5-
5-FU is widely used as a chemotherapeutic agent because of its capacity to induce apoptosis in malignant cells. 11 Subconjunctival 5-FU injections administered after glaucoma surgery led to apoptotic cell death in the conjunctival epithelium. 12 Thus, the exposure of corneal epithelial cells to specific dosage of 5-FU may induce cellular apoptosis. To date, a limited number of studies have addressed the apoptotic effect of 5-FU on corneal epithelial cells. Apoptosis is a process of natural cell death, which is characterized by extreme heterogeneity of signal transduction pathways. It leads to DNA degradation, cell membrane damage, mitochondrial dysfunction, and several degenerative pathways.13,14 To evaluate the potential chronic toxicity of 5-FU, cultured corneal epithelial cells were used in the present study to investigate the apoptotic mechanism involved in corneal damage induced by 5-FU.
Methods
Materials
MTT (3-[4,5-dimethylthiazol-2-yl]-2,5-diphenyltetrazolium bromide), Hoechst 33342, and 5-FU were purchased from Sigma-Aldrich Chemical (St. Louis, MO). The general caspase inhibitor (Z-VAD-FMK), caspase-8 inhibitor (Z-IETD-FMK), caspase-9 inhibitor (Z-LEHD-FMK), and a TUNEL Apoptosis Detection Kit were purchased from Calbiochem (Bad Soden, Germany). A MitoLight Mitochondrial Apoptosis Detection Kit was purchased from Chemicon International, Inc. (Temecula, CA). Annexin V–FITC plus propidium iodide (PI) double staining kit was purchased from Invitrogen (Carlsbad, CA). The Bcl-2 mouse antibodies and p21 rabbit antibody were purchased from Millipore (Billerica, MA). Western Blot Chemiluminescence Reagent Plus was purchased from New England Nuclear (Du Pont, Boston, MA). Protein assay dye and agents used for electrophoresis were purchased from Bio-Rad (Richmond, CA). Horseradish peroxidase-conjugated sheep anti-mouse IgG and donkey anti-rabbit IgG were obtained from Amersham Pharmacia (Buckinghamshire, England). All other chemicals were obtained from Merck (Darmstadt, Germany).
Culture of Staten's Seruminstitut rabbit corneal epithelial cell
Staten's Seruminstitut rabbit corneal epithelial cell was cultured as published previously. 15 Cells were obtained from the ATCC (American Type Culture Collection, CCL-60) and cultured in Dulbecco's modified Eagle's minimal essential medium (DMEM) supplemented with 10% fetal calf serum, 3.8 mM L-glutamine, and 50 μg/mL gentamicin (GIBCO, Grand Island, NY) at 37°C and 5% CO2 atmosphere.
Assay of cell viability with MTT
The MTT assay was used as a marker of cell viability. Cell viability was measured with MTT dye following previously published procedures. 16 The MTT assay is based on the production of purple formazan from a methyl tetrazolium salt by the mitochondrial enzymes of viable cells. Cultured cells at a concentration of 4,000 cells/well were seeded on 96-well culture plates and allowed to form a monolayer for 24 h. The cells were then exposed to 150 μL serum-free MEM containing various concentrations of 5-FU for diverse periods. Control cells were treated with phosphate-buffered saline (PBS). For analysis of apoptotic caspase pathways, cells were preincubated with different caspase inhibitors, including a general caspase inhibitor (Z-VAD-FMK), a caspase-8 inhibitor (Z-IETD-FMK), or a caspase-9 inhibitor (Z-LEHD-FMK) for 1 h before addition of 1 mg/mL 5-FU for 15 h. After exposure to the drug for 15 h, cells were washed twice with PBS and incubated with 150 μL MTT solution (0.833 mg/mL in PBS) for 4 h at 37°C. At the end of the incubation period, MTT solution was carefully aspirated, taking care not to disturb the crystal of purple formazan at the bottom of each well. The formazan reaction product was dissolved by the addition of 150 μL dimethyl sulfoxide (DMSO), and the optical density of the fluid in each well was read at 510 nm in a multiwell spectrophotometer (Titertek Multiscan, Flow Lab, Scotland, UK). Cytotoxicity was calculated based on a significant difference in the optical density between the 5-FU-treated and negative control groups.
Flow cytometry assay of apoptotic changes in the cell membrane and mitochondrial membrane potential
Apoptosis in the cell membrane was identified by flow cytometry with annexin V–FITC/PI double staining. Mitochondrial membrane potential changes were assayed with MitoLight dye. In the absence or presence of various concentrations of 5-FU, cells were incubated with 1 μg/mL annexin V–FITC and PI for 10 min or 50 μL prediluted MitoLight solution (900 μL water, 1 μL MitoLight dye, and 100 μL 10X incubation buffer) for 15 min, following the manufacturer's instructions. Cells were then analyzed by flow cytometry (Becton, Dickinson and Company) for a cell count of 10,000.
DNA staining with Hoechst 33342
To identify the apoptotic cells, DNA was viewed by staining with Hoechst 33342, a DNA-specific dye, at a final concentration of 1 mg/mL at 37°C for 1 h. Nuclear morphology was assessed by fluorescence microscopy with filters for blue fluorescence.
Apoptotic TUNEL staining on DNA
Immunohistochemical evidence for DNA strand breaks was obtained using the terminal deoxyribonucleotidyl transferase (TdT)-mediated deoxyuridine-5′-triphosphate-digoxigenin (dUTP) nick-end labeling (TUNEL) assay. The cells were cultured on coverslips for at least 24 h for attachment and then incubated with various concentrations of 5-FU at 37°C. After exposure to 5-FU, the cells were washed twice with TBS (20 mM Tris–HCl and 140 mM NaCl, pH 7.6). According to the manufacturer's instruction, the monolayer of cells was fixed for 20 min in 4% formaldehyde at room temperature and then washed thrice in TBS. Cell membrane permeability was increased by treating the samples with 20 μg/mL proteinase K solution at room temperature for 5 min. Endogenous peroxidase activity was quenched by immersing the specimens in 2% H2O2 at room temperature for 5 min. The samples were rinsed by replacing the hydrogen peroxide solution with labeling buffer and reaction mixture solution. The specimens were then placed in a humidified incubator for 1 h at 37°C. The labeling reaction was stopped by immersion in stop buffer at room temperature for 5 min. The specimens were subsequently washed in TBS. Then, 100 μL streptavidin–horseradish peroxidase was applied to the cells for 20 min at room temperature. After washing with TBS, the specimens were immersed in diaminobenzidine (DAB) solution at room temperature for 5–10 min until a satisfactory color reaction was achieved. This assay was validated with the control slides, which had been ascertained to contain apoptotic (positive control) and nonapoptotic (negative control) cells.
Western blot assay of proteins involved in apoptosis
The Bcl-2 and p21 proteins were detected by sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE) following previously published procedures. 17 Briefly, cells were treated with 5-FU and washed with 10 mL of buffer A (20 mM N-2-hydroxyethyl-1-piperazine-N'-2-ethanesulfonic acid [HEPES], 1 mM ethylene diamine tetraacetic acid [EDTA], 2 μg/mL aprotinin, 2 μg/mL leupeptin, and 1 μg/mL pepstatin A, pH 7.4). Cells were then scraped into ice-cold buffer A and immediately homogenized. Protein concentrations were determined with Bio-Rad protein assay dye. Ten micrograms of protein from each sample was added to SDS-PAGE sample buffer and heated in a boiling water bath for 5 min. An aliquot was then subjected to 10% SDS-PAGE. The proteins separated by SDS-PAGE were transferred in a Bio-Rad Trans-Blot Cell onto nitrocellulose membranes. Transfer was performed at 100 V for 2 h in a buffer containing 25 mM Tris–HCl, 190 mM glycine, 0.01% SDS, and 20% methanol. The blots were blocked at room temperature for 1 h with buffer solution (20 mM Tris–HCl and 137 mM NaCl, pH 7.6) containing 5% nonfat milk and 0.1% Tween 20. Incubation was then performed for 1 h at 37°C with monoclonal anti-Bcl-2 (1:50) or rabbit anti-p21 (1:250) antibodies. The nitrocellulose membrane was washed thrice with the same buffer used during the blocking phase and incubated with horseradish peroxidase-conjugated sheep anti-mouse IgG (1:2,000) or donkey anti-rabbit (1:10,000) as secondary antibody at room temperature for 1 h. After washing, immunocomplexes were visualized by adding Western Blot Chemiluminescence Reagent Plus. The molecular size of the immunoreactive bands was determined in comparison with a set of molecular weight marker proteins (Bio-Rad). Relative band intensity was then analyzed using the LabWorks software 4.6 from UVP Bioimaging Systems (Upland, CA).
Statistical analysis
Data were analyzed by one-way ANOVA, followed by Dunnett's post-hoc analysis. The values were expressed as mean ± standard deviation (SD). All data were significantly different from the corresponding control or 5-FU-only-treated group at P < 0.05.
Results
Effects of 5-FU on cell viability and apoptotic caspase pathways
After incubation with 5-FU for 4, 15, and 24 h, the cell viability was significantly decreased in a time- and dose-dependent manner at concentrations ranging from 0.1, 0.5, and 1 mg/mL to 5 mg/mL (Fig. 1A). To investigate the role of caspases in 5-FU-induced apoptosis, the cells were pretreated with various caspase inhibitors for 1 h and then incubated with 1 mg/mL 5-FU for 15 h. After application of the caspase inhibitors, the cellular MTT values were significantly increased in comparison with the 5-FU-only group. Compared with the corresponding control group, the 5-FU-only cells had lower MTT values (as shown in Fig. 1B). The addition of caspase-8, general caspase, and caspase-9 inhibitors (Z-IETD-FMK, Z-VAD-FMK, and Z-LEHD-FMK, respectively) at 10−5 reversed the 5-FU-induced cellular damage.
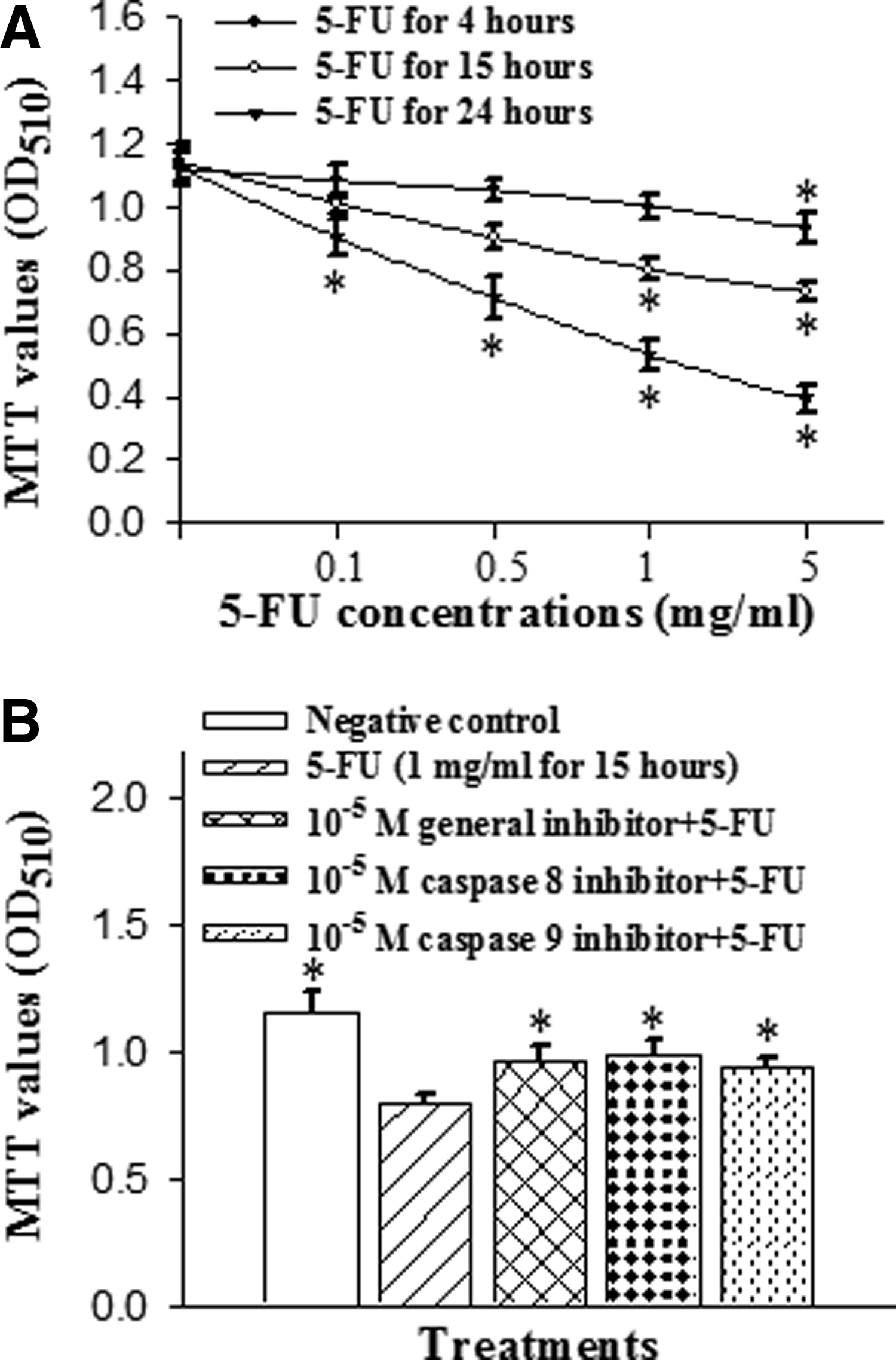
Effects of 5-fluorouracil (5-FU) on cell viability and apoptotic caspase pathways in corneal epithelial cells.
Assay of mitochondrial membrane potential change after 5-FU exposure
Disruption of the mitochondrial transmembrane potential is one of the earliest intracellular changes induced by apoptosis. In healthy cells, MitoLight dye accumulates in the mitochondria and yields red fluorescence. In apoptotic cells, where the mitochondrial membrane potential has been depolarized, the dye aggregates in the cytoplasm and emits green fluorescence, allowing discrimination of apoptotic and nonapoptotic cells. The red color area of control cells indicates normal mitochondrial membrane potential. Following exposure to 0.5 mg/mL 5-FU for 15 h, indicated with the black color, as well as the green and blue color lines of the cells treated with 1 mg/mL and 5 mg/mL 5-FU, respectively (Fig. 2A), the fluorescence of the MitoLight dye in the cells showed an obvious change, with red fluorescence in the mitochondria and green in the cytoplasm, as detected by the shift in the relative fluorescence intensity in Fig. 2B. The percentage of MitoLight that accumulated in the mitochondria of the control (100.3% ± 0.2%) decreased to 80.3% ± 0.4% at 0.5 mg/mL, 68.3% ± 0.5% at 1 mg/mL, and 60.2% ± 0.4% at 5 mg/mL (Fig. 2B). The percentage of MitoLight increased in the cytoplasm from 0.5% ± 0.1% in the control to 20.4% ± 0.3% at 0.5 mg/mL, 32.2% ± 0.4% at 1 mg/mL, and 40.3% ± 0.5% at 5 mg/mL (Fig. 2B).

Flow cytometry assay of mitochondrial membrane potential changes with MitoLight dye in cultured corneal epithelial cells in the presence of 0.5, 1, and 5 mg/mL 5-FU for 15 h.
Apoptotic TUNEL staining and Hoechst 33342 staining
Apoptosis was characterized by TUNEL and Hoechst 33342 staining. Morphological evidence of apoptotic cells is characterized by chromatin condensation in the nucleus after treatment with 5-FU. No DNA fragmentation was detected in the PBS-treated negative control after 15 h (Figs. 3A and 4A). In comparison to the negative staining in the control (Figs. 3A and 4A), some of the cells exposed to 0.5 mg/mL 5-FU for 15 h exhibited sparsely cellular condensed chromatin, which was stained with brown color by TUNEL staining (Fig. 3B) but is not obviously observed by Hoechst 33342 staining (Fig. 4B). Figures 3C and 4C shows that the cells treated with 1 mg/mL 5-FU exhibit more apoptotic characteristics. The 5-FU induced apoptosis in a dose-dependent manner, and a decrease in viable cell number correlated with an increase in the percentage of apoptosis. Following exposure to 5 mg/mL for 15 h, almost all the cells showed typical apoptotic characteristics with more bright fluorescence in Hoechst 33342-stained cells (Fig. 4D) and more positive brown in TUNEL-stained cells (Fig. 3D).
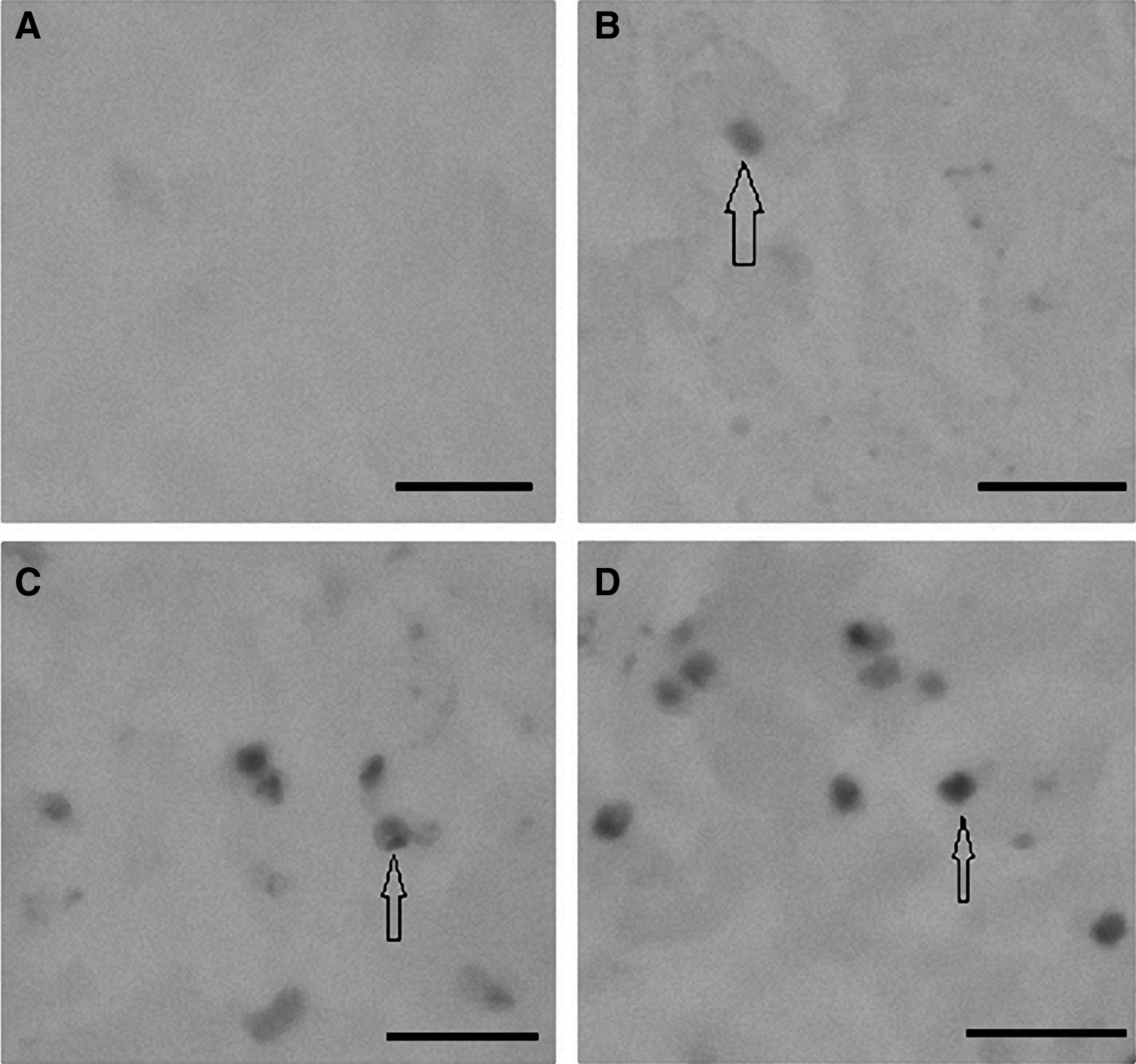
Apoptotic DNA characteristics of corneal epithelial cells visualized with TUNEL staining.
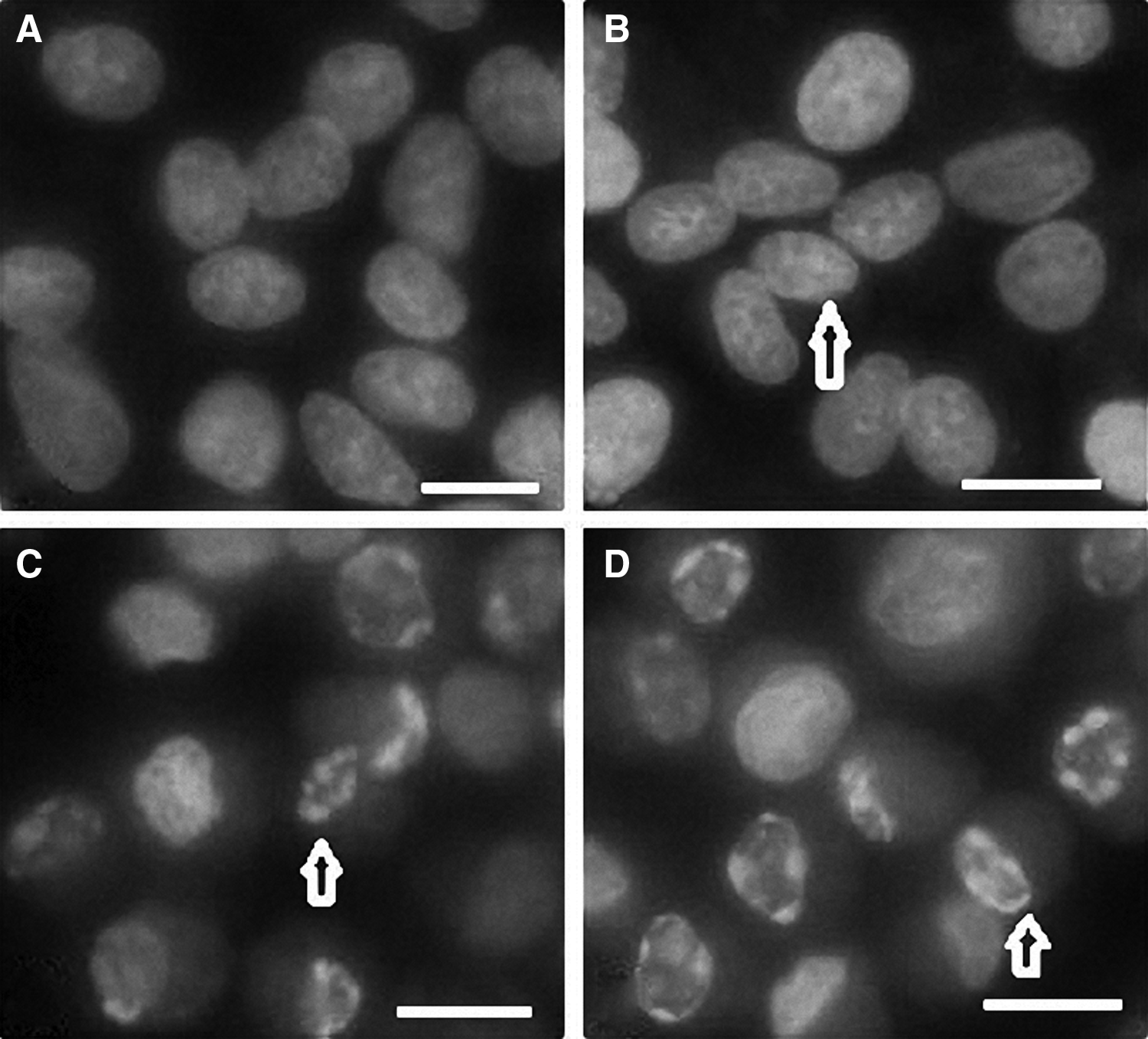
Apoptotic DNA characteristics of corneal epithelial cells visualized with Hoechst 33342 staining.
Annexin–PI staining of apoptosis in cell membrane with flow cytometry
To identify the apoptotic effect of 5-FU on cell membrane, annexin V–FITC and PI double staining of cells was performed, and the stained cells were examined with a flow cytometry. The results suggested that 5-FU appeared to have less of an apoptotic effect on the cell membrane than on cellular organelles, such as the mitochondria and DNA of nucleus, in corneal epithelial cells. In the nonapoptotic viable control cells, the annexin V–FITC and PI-negative stained cells are located in the bottom left quadrant of the dots (Fig. 5A). After exposure of cells to 0.5, 1, and 5 mg/mL 5-FU for 15 h, a significant proportion of cells (0.37% ± 0.04%, 2.25 ± 0.03%, and 2.21% ± 0.04%, respectively, compared to the control of 0.17% ± 0.03%) showed positive annexin V–FITC and negative PI staining, depicted by the significant increase in the dot numbers in the bottom right quadrant (Fig. 5B–D). The cells at this stage of apoptosis were still viable. Following exposure to 1 and 5 mg/mL 5-FU for 15 h, only the PI-positive stained cells (upper right quadrant) increased significantly to 17.5% ± 0.04% and 24.09% ± 0.05% compared to the control (5.62% ± 0.03%), indicating that the cells were no more viable (Fig. 5C, D).
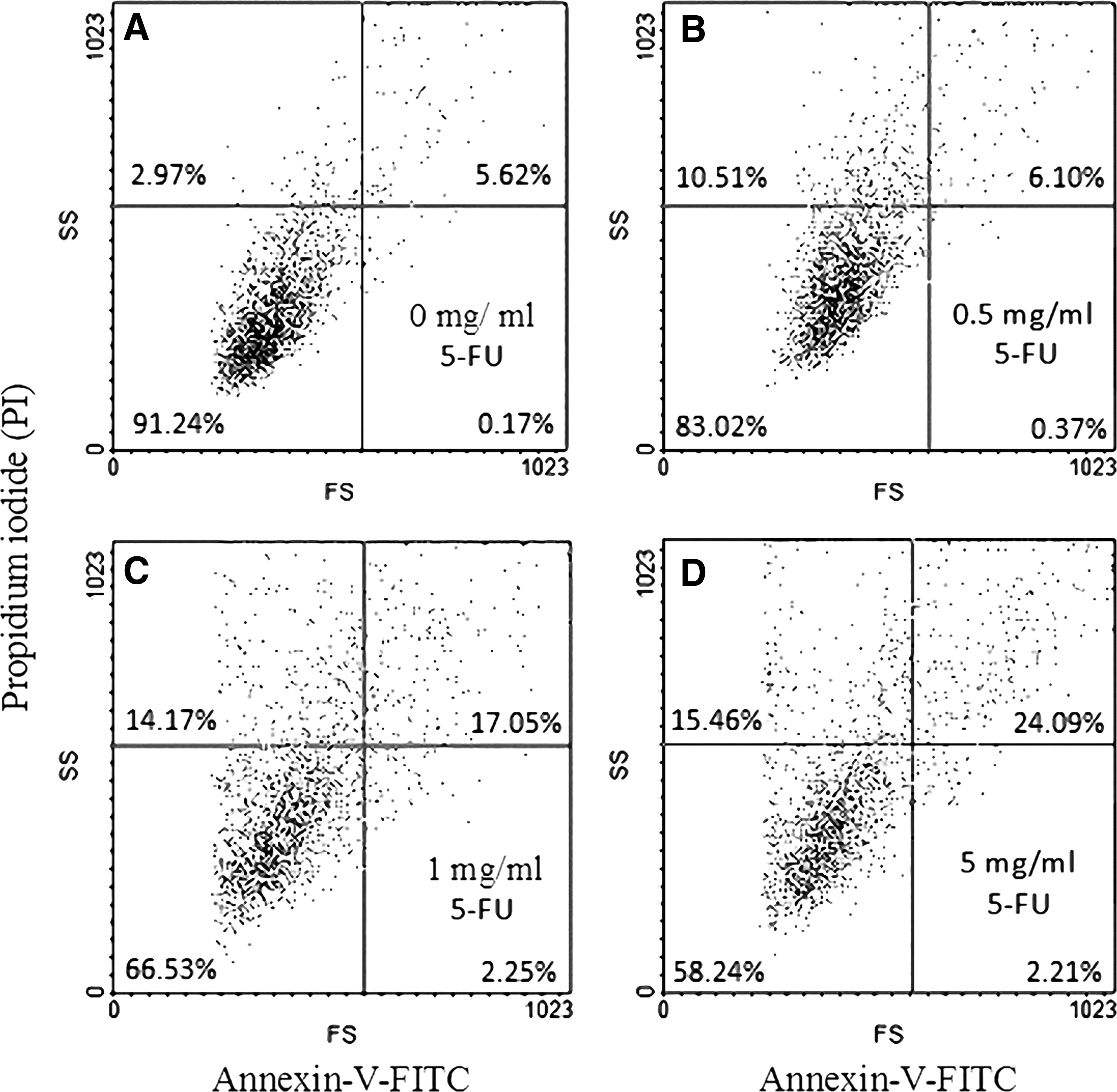
Flow cytometry analysis of cell membranes with annexin V–FITC and PI double staining.
Immunoblot analysis of proteins involved in apoptosis
To investigate the effect of 5-FU on proteins involved in apoptosis in the cultured corneal epithelial cells, we examined the antiapoptotic protein (Bcl-2) and apoptotic protein (p21) by western blotting. The results of 3 independent experiments demonstrated that 5-FU significantly increased the amount of p21 proteins but decreased the Bcl-2 protein in a dose-dependent manner (Fig. 6). Densitometric analysis of Bcl-2 protein bands showed that the optical density of the proteins in control was 60 ± 2, and the optical densities in the 5-FU-treated cells were 55 ± 3 (0.5 mg/mL 5-FU) and 43 ± 2 (5 mg/mL 5-FU). In comparison with the control cells (3% ± 1%), the optical density for p21 in the 5-FU-treated cells increased to 24% ± 3% with 0.5 mg/mL exposure and 56% ± 4% with 5 mg/mL exposure. The data are representative of the average from 3 independent experiments.
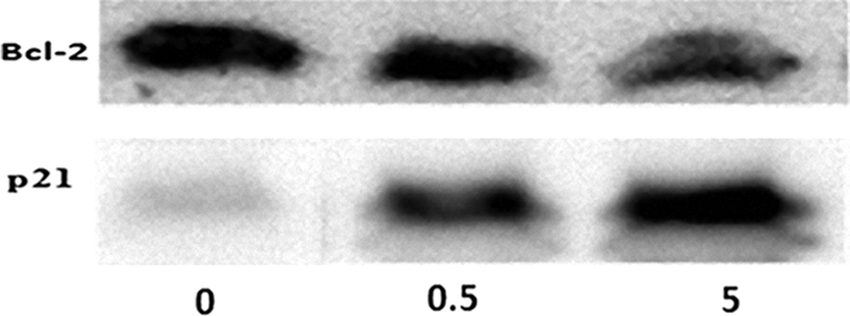
Western blot assay of proteins involved in apoptosis in corneal epithelial cells. Following incubation of the cells in the absence of 5-FU (labeled as 0 in the image) or in the presence of 0.5 mg/mL (labeled as 0.5 in the image) and 5 mg/mL (labeled as 5 in the image) 5-FU for 15 h, the cells were subjected to sodium dodecyl sulfate–polyacrylamide gel electrophoresis and immunoblotting using antisera against Bcl-2 and p21. Two other independent experiments produced similar results.
Discussion
The present study shows that the 5-FU treatment not only decreased cell viability but also induced apoptosis. In the corneal epithelial cells, the apoptotic characteristics of 5-FU were found with small partial turnover of cell membrane phosphatidylserine, significant DNA degradation, destruction of the mitochondrial membrane potential, and upregulation of p21 protein and caspase-8 and caspase-9, and downregulation of Bcl-2 protein.
In cancer cells, 5-FU inhibited cell growth and induced apoptosis, which was accompanied by changes in the mitochondria and activity of caspase-9. 18 5-FU is used in glaucoma trabeculectomy surgery. Glaucoma patients may receive total doses of subconjunctival 5-FU injections between 2 and 65 mg for a period.6,19 A certain amount of 5-FU may penetrate into the ocular surface, where it causes apoptosis of corneal epithelial cells.
The apoptotic caspases induced by 5-FU are activated by 2 signal complexes: the extrinsic apoptosis pathway and the intrinsic apoptosis pathway. The extrinsic apoptosis pathway activates caspase-8, in response to the ligation of cell surface death receptors. The intrinsic apoptosis pathway activates caspase-3 and caspase-9, in response to signals originating from the mitochondria. 18 The intrinsic pathway relies on extrinsic signals to promote cell death in a cell- and tissue-dependent manner following DNA damage. 20 Evidence has shown that treatment with 5-FU increased the expression of Fas, which belongs to the death receptor pathway. 21 In the normal thymus, 5-FU-induced apoptosis is related to Fas, Bax, and caspase-3 coexpression. 22 Receptor binding results in activation of caspase-8, leading to activation of proapoptotic downstream molecules. Our data indicated that both caspase-8 and caspase-9 inhibitors reversed the 5-FU-induced damage, suggesting that 5-FU-induced apoptosis in corneal epithelial cells may occur through both intrinsic mitochondria-mediated pathway and extrinsic pathway by death receptor. However, the 5-FU-induced extrinsic apoptosis pathway, in particular in corneal epithelial cells, needs to be explored deeply.
In a previous study with human corneal epithelial cell, 1% 5-FU showed time-dependent cytotoxic changes. In our study, such amount as low as 0.5 mg/mL 5-FU caused obvious damage to the mitochondria in corneal epithelial cells. In clinical trabeculectomy, some patients received intraoperative subconjunctival local soaking with 5-FU (50 mg/mL for 4–5 min) during surgery23,24 or others received repeated subconjunctival injections of 5-FU, with an initial dose of 2–5 mg/mL once daily and up to a total dose of 65 mg.6,19
A recent study also reported that even 0.0013 and 0.00013 mg/mL 5-FU caused obvious damage to corneal endothelial cells, including actin disruption and changes in migrating cells, which they did not display elongated processes typical of control tissues, and cellular translocation into the injury zone is retarded. 25 Studies have revealed that a single 5-min application of 25 mg/mL 5-FU induced cellular apoptosis in Tenon's fibroblasts. 7 Apoptosis characteristics were also reported in conjunctival epithelium after a subconjunctival injection of 5-FU, including dying conjunctival epithelium cells with nuclear apoptotic cell death. 5 Thus, clinically high doses of repeated injections of 5-FU are highly likely to exert direct/indirect actions on corneal epithelial cells.
In hepatoma cells, 5-FU induced the upregulation of p21 and triggered the collapse of the mitochondrial membrane potential. 26 5-FU also induced expression of the proapoptotic proteins (caspase-8 and caspase-9) in HeLa cervical carcinoma. 27 The antiapoptotic protein Bcl-2 may play an important role in the regulation of corneal epithelial renewal. 28 However, the mechanism of apoptosis induction by 5-FU in ocular tissue is poorly understood at present. Our data illustrate that 5-FU induced cellular DNA fragmentation and triggered apoptosis-related proteins, for example, antiapoptotic Bcl-2 and apoptotic protein p21 in corneal epithelial cells. In response to DNA-damaging agents, the p21 protein is an important downstream effector in the growth arrest pathway. 29 Although our previous data found that 5-FU induced cellular apoptosis in corneal endothelial cells that are also mediated through caspase-8, caspase-9, and mitochondria-regulated pathways, 5-FU to corneal endothelial cells caused only upregulation of p53- and p21-dependent signal transduction pathways but no affect on Bcl-2 protein. 30
Based on the dose response of 5-FU in corneal epithelial cells, we found that disruption of the mitochondrial transmembrane potential was the first apoptotic step detected. The mitochondria then released the proapoptotic factors, such as cytochrome c, from the inner mitochondrial membrane into the cytosol, thereby activating the caspase-9 cascade. Thus, mitochondrial depolarization is an early event marker of more severe apoptotic events in the corneal epithelial cells.
The use of annexin V–FITC/PI to identify the turnover of cell membrane phosphatidylserine in the presence of 5-FU indicated that only a small number of early apoptotic cells were clearly stained by annexin V–FITC after 15 h of incubation with 1 and 5 mg/mL 5-FU. They were stained positive for annexin V–FITC and negative for PI. Comparing the apoptotic dose response for the mitochondria and DNA with that of the cell membrane suggests that the cellular membrane appears to be the most resistant to 5-FU-induced damage.
It is not easy to detect drug-induced damage of corneal epithelial cells unless the side effects of 5-FU are severe enough to interfere with normal cellular physiological function and wound healing. Many side effects of 5-FU may occur after a particular period of clinical use. The delayed healing of corneal epithelial defects and limbal stem cell deficiency have been reported with 5-FU treatment after glaucoma surgeries.8,9 Our data showed that the mitochondria of corneal epithelial cells are damaged in the presence of 0.5 mg/mL 5-FU for 15 h. The efficiency of ocular wound healing may be affected after a period of 5-FU use, particularly in cases of accidental access of 5-FU to ocular surface especially to corneal epithelial cells.
Conclusion
In conclusion, 5-FU induced significantly apoptosis in corneal epithelial cells that occurs through activation of intrinsic mitochondrial and extrinsic caspase-8 apoptotic pathways, as well as by upregulation of p21 and downregulation of Bcl-2-dependent signal transduction pathways. Although evidence suggests that 5-FU is less toxic than mitomycin C to Tenon's fibroblasts, high-dose 5-FU may result in the apoptosis of ocular surface cells. 7 The appearance of apoptotic characteristics in corneal epithelial cells may extend the chronic toxicity of 5-FU to ocular surface cells. Thus, the use of 5-FU may also need to be carefully monitored for adverse changes in corneal epithelial cell.
Footnotes
Acknowledgment
This project was supported by the research grant from the Kaohsiung Medical University Hospital (KMUH102-2 M33).
Author Disclosure Statement
The authors have no proprietary or commercial interest in any materials discussed in this article.