
Abstract
Abstract
Purines, when present in the extracellular space, can mediate fast neurotransmission in the retina and central nervous system. Over the last decade there has been emerging evidence for the expression of P2X and P2Y receptors in a range of retinal neuronal subtypes. These results have highlighted important roles for purines in modulating specific retinal circuits, including the rod pathway and amacrine cell circuits. Traditionally, synaptic release of adenosine triphosphate (ATP) involves the novel anion vesicular nucleotide transporter, VNUT, which has recently been identified in a single wide-field amacrine cell population. In addition, nontraditional, conductive mechanisms of release have also been described in the retina. In the synapse, the enzymes involved in rapid degradation of purines are present in both plexiform layers of the retina. A role for P2X receptors in retinal diseases has also emerged recently. High concentrations of ATP lead to photoreceptor loss, through mechanisms involving P2X7 receptors. In addition, activation of P2X7 receptors is associated with activation of the inflammasome, a protein complex important for the release of proinflammatory cytokines. P2X receptors, especially P2X7, are emerging as targets to combat retinal disease.
Introduction
N
In addition to a role in normal signaling within the retina, there is growing evidence that anomalies in purinergic signaling contribute to retinal disease.12,13 Genetic association studies have implicated purinergic receptors in the development of age-related macular degeneration (AMD). 14 Abnormal signaling through P2X7 receptors has been implicated in cell death in the central nervous system.15,16 ATP is found in high concentration within cells and following cell death can be released into the extracellular milieu. 15 Further cell death can follow, possibly through pathological overactivation of the receptors for ATP. Indeed, extracellular ATP has been shown to cause both photoreceptor death17–19 as well as neurotoxicity within the ganglion cell layer in the retina. 20
The aim of this review is to outline the role that purines, especially ATP, have in modulating specific circuits within the mammalian retina and to detail how anomalies in purinergic signaling contribute to retinal diseases.
The Role of Purines in Normal Retinal Processing
Localization of P2X receptors in the mammalian retina
The actions of ATP and its analogs are mediated by 2 broad families of purinergic receptors. P2X receptors are ligand-gated ion channels that are directly gated by ATP and its analogs. P2X receptors are trimers formed from individual subunits encoded by 7 distinct genes called P2X1–7. 21 These 7 P2X isoforms have a high degree of sequence homology, 22 but differing sensitivities to ATP and desensitization properties. By contrast, P2Y receptors belong to the large family of 7 transmembrane spanning G-protein -coupled receptors. To date, 8 P2Y isoforms have been identified in mammalian tissues (P2Y1,2,4,6,11,12,13,14). 23
Most purinergic receptor subtypes have been identified in the mammalian retina. Using RT-PCR and in situ hybridization, mRNA expression of P2X2-P2X5 and P2X7 receptors in the rat neural retina and retinal pigment epithelium (RPE) has been demonstrated.24–28 The gene expression of P2Y receptor subtype genes has also been identified in the neural retina and RPE of the rat, including P2Y1, P2Y2, P2Y4, and P2Y6.29,30
Elucidation of the retinal circuits that are modulated by purines has been achieved by evaluation of high-resolution immunocytochemical studies by double labeling P2X and P2Y receptors with known markers of retinal neurons. Immunocytochemistry has revealed labeling of processes within the outer plexiform layer for P2X3, P2X4, and P2X7 (Fig. 1; examples of P2X receptor immunolocalization for (A-B) P2X2, (C-D) P2X4, and (E-F) P2X7).).4,6,31 In particular, P2X7 labeling was localized presynaptically to rod and cone pedicles in rodent and primate retina.4,7 In addition, application of the P2X7 agonist, 3′-O-(4-benzoyl)benzoyl adenosine 5′-triphosphate (Bz-ATP), induced an increase in the amplitude of the a-wave of the electroretinogram implying possible depolarization of the photoreceptor membrane potential by this receptor. 7 These results suggest that P2X7 receptors have a role in regulating photoreceptor function, perhaps potentiating the release of glutamate, or alternatively modulating a range of calcium-dependent functions of photoreceptors.

Localization of P2X receptors and VNUT in the mouse retina. Vertical sections of mouse retina labeled for
In addition to photoreceptor terminals, P2X4 and P2X7 receptors were also localized to horizontal cell processes. Recent reports have suggested that close physical and functional interactions may occur between P2X4 and P2X7. 32 In addition, P2X4 and P2X7 receptors show distinct responses to ATP, desensitizing at different rates. Although it is not known whether P2X4 and P2X7 receptors are localized to the same synaptic sites on horizontal cell processes, it is possible that ATP can modulate horizontal cell function in distinct ways depending on the P2X receptor subtype activated. Overall, these results highlight the complex role that purines have in modulating signaling in the outer retina.
The amino acids GABA and/or glycine are known to mediate neurotransmission between amacrine cells and their postsynaptic partners (amacrine, bipolar, or ganglion cells). 33 A number of studies have shown colabeling of neural processes with P2X2, P2X3, or P2X7 receptors and GABA, suggesting that P2X receptors modulate signaling by GABAergic amacrine cells.4–6 In addition, specific P2X receptors are localized to the rod or cone pathways. For example, P2X2 receptors are localized to amacrine cells that are postsynaptic to cone bipolar, but not rod bipolar cells. By contrast, P2X3, P2X4, and P2X7 are localized to neurons postsynaptic to both cone and rod bipolar cells.4,6,7,31,34
Identification of the specific amacrine cell types that express P2X receptors reveals the complexity of circuits that purines may modulate. The neural circuit integral to encoding scotopic vision is well characterized and consists of rod photoreceptors synapsing with rod bipolar cells, which in turn synapse with 2 amacrine cell types, AII and A17 amacrine cells. 35 The GABAergic A17 amacrine cell receives input from rod bipolar cells and provides GABAergic inhibition through reciprocal synapses that feed back onto rod bipolar cell axon terminals. Double labeling and ultrastructural analysis has shown P2X3 and P2X7 associated with the A17 amacrine cell at both conventional and reciprocal synapses [Fig. 1B, F, respectively; showing these receptors colocalize with conventional synapses labeled with bassoon in the IPL (red) and are closely associated and postsynaptic to bipolar cells labeled with VGLUT (blue)]. This suggests that GABA inhibition of the rod bipolar cell function may be regulated by purines acting on P2X3 and/or P2X7 receptors. In particular, activation of P2X3 and/or P2X7 receptors potentiates the release of GABA from A17 amacrine cells. In line with this finding, administration of the P2X7 agonist, Bz-ATP, was associated with a reduction in the amplitude of the scotopic b-wave of the ERG, 7 whereas genetic depletion of P2X7 receptors was associated with an increase in scotopic b-wave amplitude. 34 These results imply that ATP-induced GABA release plays a role in the regulation of rod bipolar cells.
Purinergic receptors may have a role in modulating a major neural circuit within the inner retina that has been implicated in motion detection. Starburst amacrine cells are well described neurons that coexpress acetylcholine and GABA. P2X2 receptor expression has been demonstrated in subsets of amacrine cells in the rodent retina 5 (Fig. 1A), including OFF-type cholinergic amacrine cells in the mouse, indicating a possible role for the P2X2 receptor in modulating the function of this circuit. 36 Consistent with this finding, patch-clamp recordings of mouse OFF-cholinergic amacrine cells in response to exogenous ATP showed a slow inward current and an increase in the frequency of inhibitory postsynaptic current 37 that was blocked by the P2 antagonist, pyridoxal-phosphate-6-azophenyl-2′,4′-disulfonic acid (PPADS) as well as the GABAA receptor antagonist, SR95531. In contrast, ON-cholinergic amacrine cells showed little response to ATP or application of P2X antagonists. 37 These results suggest that purines regulate the ON and OFF starburst amacrine cells in different ways; GABAergic inhibition of OFF cholinergic starburst amacrine cells is mediated by P2X receptors. The functional role for this cell-specific modulation by purines may be important for adjusting the threshold of OFF response to a mean light sensitivity. 37
In addition to being expressed by neurons, several P2X receptors, including P2X4 and P2X7, are expressed by microglia, the resident immune cells of the central nervous system (CNS). 14 Under normal circumstances, microglia perform an important surveillance role by dynamically contacting synapses. ATP is thought to be an important signaling molecule regulating this dynamic activity. 38 In addition, ATP is known to be a potent signaling molecule for activation and chemotaxis of microglia to the CNS and plays an important role in the neuroinflammatory response associated with injury and disease. 39
Localization of P2Y receptors in the retina
P2Y receptors elicit neural responses through activation of G protein-mediated signaling. The cellular localization of the metabotropic P2Y1, 2, 4, and 6 receptor subtypes have been identified in the human and rodent retina.8,9,29,40,41. The P2Y1 receptor has been localized to cholinergic amacrine cells and ganglion cells in the rat retina.8,42 Similarly, P2Y2 and P2Y6 receptor expression are restricted to ganglion cells and cells of the inner nuclear layer of the rat retina. 41 P2Y4 receptor expression has been demonstrated in rod bipolar cell dendrites and axon terminals, and amacrine cell processes postsynaptic to cone bipolar cells, 9 suggesting a role in the modulation of outer and inner retinal processing. The association of the P2Y4 receptor with inner retinal function has been further demonstrated by a reduction in amplitude of both the scotopic b-wave following application of uridine triphosphate (UTP), a nonspecific P2Y receptor agonist in the rat retina. 9
Various P2Y receptor subtypes are expressed by Müller cells. The P2Y1 receptor is the principal purinergic receptor expressed in Müller cells,9,41 and is involved in the glutamatergic-purinergic signaling cascade that enables Müller cells to maintain their cell volume. 43 P2Y4 receptor immunoreactivity has been localized predominantly to the Müller cell endfeet in the rat retina, 41 which suggests a role in homeostatic support of neuronal and vascular activities, particularly in the inner retina. 44 Moreover, P2Y4 receptors have been shown to regulate osmotic volume in murine retinal slices when activated by exogenous uracil nucleotides (UTP and UDP). 45 Immunolabeling analyses of P2Y2 and P2Y6 receptors demonstrated expression in Müller cell processes in the human retina 40 and a possible role in regulating osmotic volume. 45
Storage and release mechanisms of ATP
In order for ATP to affect neural function, a mechanism for ATP release at synaptic sites is necessary. ATP release mechanisms can be broadly classified into 2 types: conductive mechanisms and exocytotic mechanisms. Conductive mechanisms involve passive, osmotic diffusion into the extracellular space across volume-regulated ion channels, pore-forming connexins, pannexins, or P2X7 receptors. 46 Exocytotic mechanisms involve ATP packaging within synaptic vesicles and Ca2+-regulated exocytotic release of purines at synaptic sites. 11
Connexin and pannexin hemichannels have been implicated in the release of ATP through conductive mechanisms. These channels are widely expressed in the CNS. Connexin hemichannels can open in response to a reduction in extracellular Ca2+ concentration and membrane voltage, allowing ATP efflux from the cytoplasm into the extracellular space through these large diameter pores.47,48 In the retina, astrocytes and Müller cells are coupled extensively through connexin gap junctions, 49 and ATP release from murine Müller cells has been suggested to occur through connexin-43 hemichannels. 50 The role of pannexins in ATP release in the retina is less well undertstood. Pannexins are expressed widely across the retina in amacrine and horizontal cells 51 and analysis of ATP release in the retina in response to mechanical stimulation has implicated this channel type. 52
Conductive mechanisms of ATP release may also be integral to P2X7 receptor function. Following extended exposure to ATP, P2X7 receptors undergo a conformational change leading to the formation of a large pore. These large conductance pores allow the influx of molecules as large as 900 kDa and in turn can also facilitate the release of ATP.53,54 This mechanism of ATP release that may be particularly important in microglial signaling in the CNS activation of P2X7 receptors in mouse cortical microglia has been shown to release ATP.55,56 Conversely, the P2X7 receptor has been functionally associated with pannexin-1 channels in isolated human macrophages 57 and purified rat retinal ganglion cells, 52 whereby pannexin-1 may have been the actual conduit for the release of ATP at a sufficiently high concentration to activate the P2X7 receptor. Although P2X7 receptor pore formation has been demonstrated on retinal microglial cells and capillaries,58,59 it remains unknown whether ATP-mediated P2X7 receptor activation-dependent ATP release contributes to retinal function.
Exocytotic release of neurotransmitters is integral to synaptic release of a range of neurotransmitters in the CNS and there are a number of neurotransmitters that utilize vesicular transporters that belong to the large SLC17 family of anion transporters (SLC17). Recently, a novel member of the SLC17 family of anion transporters, vesicular nucleotide transporter (VNUT), encoded by the solute carrier 17, member 9 (SLC17A9) gene, has been identified implying that a similar vesicular packaging mechanism exists for purines. 11 VNUT expression has been identified in a range of tissues, including adrenal gland, 11 stomach, liver, 60 spleen, blood, 61 immune T-cells, 62 hippocampal neurons, 63 and also in the retina. 34 Moreover, depending on the antibody dilution used, it is possible to detect either punctate or cellular labeling for VNUT. Indeed, punctate labeling was observed in sublamina 1 of the IPL when a very low antibody dilution is used, implying labeling of synaptic vesicles. Double labeling immunocytochemistry revealed that VNUT colocalizes extensively with P2X7 receptors in the processes of dopaminergic neurons of the retina and substantia nigra 10 (Fig. 1G, H). Of the 4 predicted isoforms of VNUT, 2 are expressed in the retina and brain. 10 In view of the labeling of only one cell type in the retina, it is likely that ATP release in the retina involves multiple mechanisms, or there are as yet unidentified isoforms of vesicular ATP transporters that would account for ATP release from the remaining retinal neurons.
Degradation mechanisms of ATP
An effective extracellular degradation mechanism for purines is a crucial component of purinergic signaling within the nervous system. For ATP and related analogs, this is achieved by the activity of a family of ectonucleotidases that catalyze the sequential breakdown of ATP to ADP and then the breakdown of ADP and AMP and adenosine. There are at least 4 major groups of ectoenyzmes involved in this catalytic process, ectonucleoside triphosphate diphosphohydrolase (E-NTPDase), ectonucleotide pryophosphatase (E-NPP), alkaline phosphatase, and the ecto-5′-nucleotidase family. NTPDases are thought to be the most abundant of all ectonucleotidases in the CNS. Ecto-ATPases have been identified in both the outer and inner plexiform layers using enzyme histochemistry.6,7
In summary, over the last decade, there has been a dramatic increase in our understanding of the role that purines play in modulating neural processing within the retina. A range of retinal cell types are now known to express P2X and P2Y receptors including macro and microglia (Fig. 2A), as well as distinct neuronal circuits (Fig. 2B–D). In addition, a specific amacrine cell type, the dopaminergic amacrine cell, expresses VNUT, the principal vesicular transporter important for packaging purines into synaptic vesicles. Mechanisms for degradation of ATP are localized to membranes within both the inner and outer plexiform layers. More work is necessary, however, to more clearly define the functional role of purines in retinal processing.
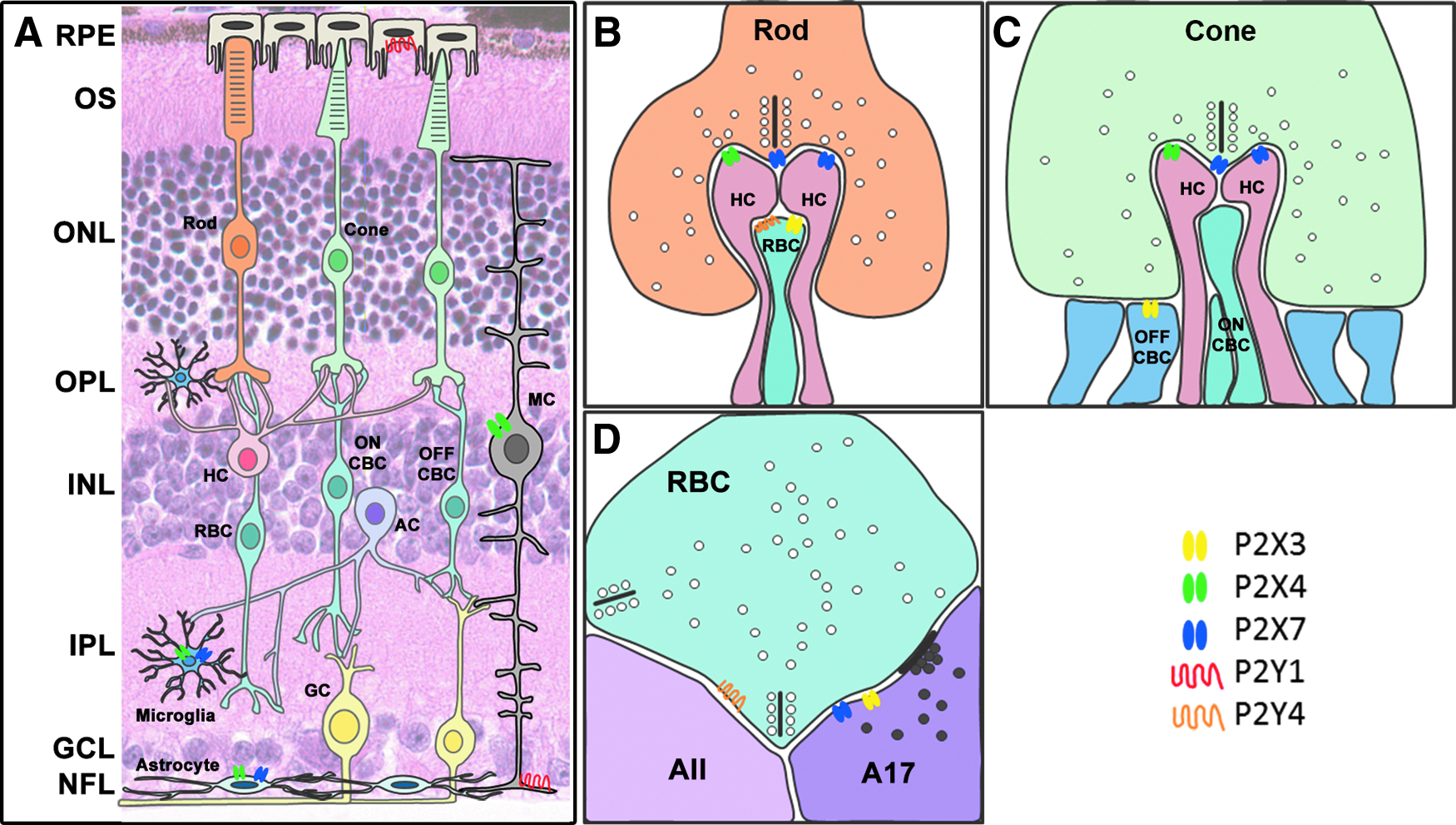
Summary of P2 receptors in the retina. Schematic diagrams showing neuronal and glial cells in the retina
Role of Purines in Retinal Disease
Abnormal purinergic signaling has been implicated in degenerative conditions of the retina, including AMD. AMD is the leading cause of irreversible blindness in people over 50 years of age in the western world. 64 There are 2 types of advanced AMD referred to as dry or wet AMD, depending on the presence of chronic atrophic changes (dry AMD) or pathological growth of blood vessels into the retina (wet AMD). The hallmark signs of early AMD include: (1) thickening of Bruch's membrane, a basement membrane located beneath the retina, between the RPE and underlying vasculature called the choroid, and (2) the presence of deposits, called drusen that develop between the RPE and choroid. 65 Photoreceptor death and loss of vision is thought to occur in dry AMD as a consequence of a lack of nutrient transfer from the vasculature to the RPE/photoreceptors. In contrast, vision loss in wet AMD occurs from growth of new blood vessels from the choroid into the retina. Thus, factors that lead to RPE and photoreceptor dysfunction and death as well as the factors that lead to pathological growth of choroidal blood vessels are crucial for enhancing our understanding of the disease and its progression. The role of P2X receptors in the development of AMD can be considered in 2 distinct ways. First, aberrant activation of P2X receptors, especially P2X7 receptors, may contribute to neuronal death through calcium-dependent mechanisms. Alternatively, P2X receptors expressed by retinal immune cells may contribute to the abnormal activation of the inflammatory cells. Below, we consider the role of P2X7 receptors in direct death of neurons and as a bystander of P2X7 receptor activation on immune cells.
The role of ATP in mediating neuronal death
Neuronal death has been attributed to direct activation of P2X receptors, notably P2X7 receptors, by extracellular ATP. P2X7 receptors are highly permeable to calcium ions, and excessive activation can lead to a substantial increase in intracellular Ca2+ concentration. Additionally, stimulation of P2X7 receptors on neurons in vitro has been shown to induce sustained Ca2+-store depletion, cellular stress, and subsequent cell death. 66 Evidence for a role of purines in neuronal death comes from studies examining spinal cord injury and cerebral ischemia. Indeed, spinal cord injury is associated with an increase in extracellular ATP and subsequent death of neurons that can be abrogated by treatment with P2X7 receptor antagonists.15,16 Similarly, cerebral ischemia is associated with neural death that can be reduced by P2X7 blockade. 67
Photoreceptor death is a key cause of vision loss in both AMD and inherited retinal degeneration. In addition, patients with subretinal hemorrhage, a common event in advanced wet AMD, have a particularly poor visual prognosis. A number of studies have highlighted the potential role that ATP plays in death of photoreceptors. The concentration of ATP in the vitreous is known to be increased in patients with a combination of AMD and vitreous hemorrhage. 68 Subretinal hemorrhage is associated with significant photoreceptor death, a finding that has been attributed to the high concentration of ATP in whole blood. 68 Intravitreal and subretinal injection of ATP induces rapid and selective photoreceptor loss in rodent or feline retinae (Fig. 3).17–19,68 Moreover, cotreatment with the nonspecific P2X antagonist, PPADS, or the P2X7 antagonist, Brilliant Blue G, reduced photoreceptor loss.17,68 In contrast, subretinal or intravitreal injection of ATP or the P2X agonist, bzATP in P2X7 null mice, had minimal effect on photoreceptor integrity.68,69 Finally, treatment with a P2X antagonist reduces photoreceptor death associated with subretinal hemorrhage, 68 or inherited retinal degeneration. 17 These results all highlight the contribution that ATP has on modulating photoreceptor integrity.

ATP induces photoreceptor loss in the feline eye. Vertical sections of a normal feline retina
Abnormal signaling by purines has also been implicated in death of retinal ganglion cells both in vitro and in vivo. Notably, application of the P2X agonist, BzATP to the retina, leads to death of ganglion cells in human organotypic retinal cultures as well as in vivo in the rat retina, and can be blocked by application of the P2X7 receptor antagonists, Brilliant Blue G and MRS-2540.20,70 In addition, administration of Brilliant Blue G reduces ganglion cell death in rats following experimentally induced elevation in intraocular pressure. 71
In view of the broad expression of P2X receptors across many neuronal subtypes in the retina, it is perhaps surprising that ATP selectively ablates photoreceptors and P2X agonists kill ganglion cells. It is not clear why ATP selectively ablates photoreceptors. One possibility is that death of photoreceptors occurs as a secondary event to ATP-induced functional changes in support cells such as RPE and/or glial cells. Indeed, ATP is known to modulate glial cell regulation of fluid balance within the retina 72 and RPE cells are known to be regulated by P2X7 receptors.73,74 However, it should be noted that a previous study examining ATP-induced photoreceptor death showed at the ultrastructural level that RPE cells were intact, and contained outer segment debris, suggesting that not only do RPE cells remain following ATP-induced death, but they also show evidence of function. 17 Alternatively, there may be differences in the expression of adenosine receptors across the retina, including in photoreceptors. Adenosine, a breakdown product of ATP, is known to be neuroprotective. Indeed, application of P2X agonists in combination with an adenosine receptor antagonist leads to ganglion cell death. 75 Adenosine receptors are known to be expressed across the retina; A3 receptors in ganglion cells, whereas other adenosine subtypes are expressed in other retinal neurons. More work is needed to determine whether selective expression of specific adenosine receptor subtypes, such as the A3 receptors on ganglion cells, explains the selective neuroprotection observed in ganglion cells. 76
The role of P2X receptors in inflammation
Immune cells, including microglia and macrophages, are known to express P2X and P2Y receptors and can respond in a variety of different ways to ATP.14,39 P2X7 receptors, in particular, play a powerful role in activating the innate immune system. On activation of P2X7 receptors by extracellular ATP leads to opening of a cation channel, which gradually dilates to form a large pore. This is associated with a large efflux of K+ membrane, membrane blebbing, release of cytokines, and cell death. The large efflux of K+ that occurs following formation of a P2X7 pore acts as a stimulus for the formation of the inflammasome, a protein complex consisting of caspase 1, apoptosis-associated speck-like protein (ASC) and nod-like receptor protein (NRLP1 or NRLP3) that is important for catalyzing the maturation of IL1-β. With respect to AMD, activation of the inflammasome, particularly NLRP3, has implicated in regulating RPE cell loss in a model of AluRNA-induced RPE degeneration. 77
P2X7 receptors as scavenger receptors
An intriguing less well-characterized function of P2X7 receptors is its role as a scavenger receptor. 78 Phagocytosis of cellular debris by macrophages involves recognition by scavenger receptors of moieties exposed on the surface of dead or dying cells. Moieties on the surface of extracellular debris or dying cells are recognized by 6 cysteine residues in the extracellular domain of P2X7 receptors, allowing binding and engulfment. 79 Moreover, in macrophages, P2X7 receptors are tightly associated with nonmuscle myosin H chain, the ATPase that provides the energy for cytoskeletal rearrangement needed for engulfment.79–81 These studies suggest that activation of P2X7 receptors by ATP inhibits the innate scavenger activity and phagocytosis function of P2X7 receptors. 78 Thus, under physiological conditions, when extracellular ATP levels are low, P2X7 receptors could play a scavenger role. However, in a situation where extracellular ATP increases, such as might occur following neural death or injury, the phagocytic role of P2X7 receptors is inhibited, and the inflammatory/neurotoxic role potentiated. 82 With respect to retinal disease, a recent association study showed that inheritance of a combination of loss of function single nucleotide polymorphisms in P2X7 and P2X4 increased the risk of developing advanced AMD 4-fold. In addition, in a subset of patients heterozygous for both the P2X4 and PX7 single nucleotide polymorphism, phagocytic function of peripheral blood monocytes was reduced. 14 More work is needed to determine the precise role that each of the 2 roles of P2X7 has in contributing to AMD.
Conclusions
Over the last decade our knowledge of the role that purines play in regulating retinal function in the normal and diseased retina has increased dramatically. The receptors for purines, including P2X and P2Y receptors, are now known to be expressed by virtually every cell type in the retina. P2X7 receptors have an intriguing role in potentiating photoreceptor function and more recently have been implicated in their death. By contrast, a number of P2X receptors, including P2X3 and P2X7, have been localized to amacrine cells, which are postsynaptic to rod bipolar cells, implicating purines in modulation of the scotopic rod pathway, perhaps by regulating A17 amacrine cells.
With respect to retinal disease, there is emerging evidence that abnormal activation of P2X receptors, leads to loss of photoreceptors and ganglion cells. These studies highlight that P2X receptors could be a novel target for neuroprotection. In addition to its role as a ligand-gated ion channel, the P2X7 receptor has been implicated in the phagocytosis of debris. More work is needed to fully understand the role of this receptor as a scavenger receptor.
Footnotes
Acknowledgments
This work was supported by the National Health and Medical Research Council of Australia (#APP1027624, #APP1061419) and the Macular Disease Foundation of Australia.
Author Disclosure Statement
No competing financial interests exist.