
Abstract
Abstract
Purpose:
In the P23H rat model of retinitis pigmentosa, the dopamine D2 receptor antagonists sulpiride and eticlopride appear to improve visual responses of retinal ganglion cells (RGCs) by increasing light sensitivity of RGCs and transforming abnormal, long-latency ON-center RGCs into OFF-center cells. Antipsychotic drugs are believed to mediate their therapeutic benefits by blocking D2 receptors. This investigation was conducted to test whether haloperidol (a typical antipsychotic drug) and clozapine (an atypical antipsychotic drug) could similarly alter the light responses of RGCs in the P23H rat retina.
Methods:
Extracellular recordings were made from RGCs in isolated P23H rat retinas. Responses of RGCs to flashes of light were evaluated before and during bath application of a drug.
Results:
Both haloperidol and clozapine increased light sensitivity of RGCs on average by ∼0.3 log unit. For those ON-center RGCs that exhibit an abnormally long-latency response to the onset of a small spot of light, both haloperidol and clozapine brought out a short-latency OFF response and markedly reduced the long-latency ON response. The selective serotonin 5-HT2A antagonist MDL 100907 had similar effects on RGCs.
Conclusions:
The effects of haloperidol on light responses of RGCs can be explained by its D2 receptor antagonism. The effects of clozapine on light responses of RGCs on the other hand may largely be due to its 5-HT2A receptor antagonism. Overall, the results suggest that antipsychotic drugs may be useful in improving vision in patients with retinitis pigmentosa.
Introduction
R
One cell type that appears to be affected in degenerating retinas is the dopaminergic amacrine cell.4,5 Recently, I reported that the dopamine D2 receptor antagonists sulpiride and eticlopride appear to improve the retinal responses of retinal ganglion cells (RGCs) in the P23H rat model of retinitis pigmentosa. 6 Both sulpiride and eticlopride increase light sensitivity of RGCs and transform abnormal, long-latency ON-center RGCs into OFF-center cells. It is thought that these long-latency ON-center RGCs were once OFF-center cells early in the disease process. 6 Antipsychotic drugs, commonly prescribed to treat the symptoms of schizophrenia and related disorders, are believed to mediate their therapeutic effects by antagonizing the actions of dopamine on D2 receptors.7,8 Antipsychotic drugs have been divided in 2 major categories, typical and atypical agents. The significant difference lies in their different ability to produce debilitating extrapyramidal side effects. At clinically effective doses, atypical antipsychotic drugs are less liable to produce extrapyramidal side effects. Most atypical antipsychotic drugs have a relatively high affinity for serotonin 5-HT2 receptors compared to D2 receptors. 9 Representative drugs of typical and atypical antipsychotic drugs are haloperidol and clozapine, respectively. In this study, I examined the effects of these U.S. Food and Drug Administration (FDA)-approved antipsychotic drugs on the light responses of P23H rat RGCs.
Methods
Animals
P23H-line 1 homozygous rats of 22–35 weeks of age were used in the experiments. Breeding pairs of P23H-line 1 homozygous rats were provided by Dr. Matthew LaVail (University of California, San Francisco). The rats were housed in a room that was kept on a 12 h-light/12-h dark cycle using standard fluorescent lighting. The illumination at the level of the cages was 100–200 lux. All procedures in this study were carried out in accordance with the ARVO Statement of the Use of Animals in Ophthalmic and Vision Research, and the protocol was approved by the Institutional Animal Care and Use Committee of the VA Boston Healthcare System.
Tissue preparation
A rat was euthanized with sodium pentobarbital (150 mg/kg, i.p.) and the eyes removed and hemisected under normal room light. Following removal of the vitreous humour from each eye, one eye was transferred to a vessel containing the bicarbonate-buffered Ames medium (Sigma-Aldrich), which was continuously gassed at room temperature with 5% CO2/95% O2. The retina of the other eye was peeled from the retinal pigment epithelium/choroid in the Ames medium and trimmed into a square of ∼12 mm2. This piece of retina was then placed photoreceptor side down in a small-volume (0.1 mL) chamber and superfused at 1.5 mL/min with the bicarbonate-buffered Ames medium, equilibrated with 5% CO2/95% O2. An in-line heating device (Warner Instruments) was used to maintain recording temperature at 35–36°C. The retina of the other eyecup was used later in the afternoon.
Electrophysiological recordings
Action potentials (spikes) were recorded extracellularly from single RGCs with a glass-insulated platinum/tungsten microelectrode (0.6–1.0 MΩ impedance; Thomas Recording GmbH, Germany). Extracellular potentials from RGCs were amplified and bandpass filtered at 100–5,000 Hz by a differential amplifier (Xcell-3; FHC, Bowdoin, ME). Spikes from RGCs were converted to TTL pulses with a time–amplitude window discriminator (APM Neural Spike Discriminator; FHC). A laboratory data acquisition system (1401 Processor and Spike2 software; Cambridge Electronic Design Ltd., Cambridge, UK) was used to digitize both the TTL pulses and the raw spike train data.
Light stimulation
Light from a mercury arc lamp illuminated an aperture that was focused on the retina with a 4 × microscope objective. The image produced on the retina was either a 250-μm or 1.5-mm diameter spot, which was centered on the recorded RGC. In the light path was a 545 nm interference filter (bandwidth, 30 nm). The intensity of the unattenuated light stimulus on the retina, measured with a spectroradiometer (ILT900-R; International Light Technologies), was 8.5 × 1017 photons/cm2/s (∼2 × 106 lux). In this article, the light intensities are expressed in log units relative to the unattenuated light intensity. An electromechanical shutter (Uniblitz, Rochester, NY) was used to control the stimulus duration, which was set to 100 ms in constructing intensity–response curves. Light flashes were presented with interstimulus intervals of 3–15 s to avoid any adapting effect of the previous flash. All experiments were performed under dim light (10 lux).
Drugs
Haloperidol, clozapine, and MDL 100907, which were purchased from Tocris Bioscience (Minneapolis, MN), were dissolved in a small amount of dimethylsulfoxide (DMSO) and diluted in physiological saline (0.9% NaCl). The drug solutions were applied to the bathing solution using a syringe pump (Razel Scientific Instruments). The final concentration of DMSO at the retina was ∼0.02%. Drugs were bath applied for ∼10 min to ensure stable responses before effects were examined. Only one cell was studied in each retina to avoid possible residual drug effects.
Data analysis
Intensity–response curves were constructed by counting the number of spikes within a 100 ms window that encompassed the peak response and subtracting any spontaneous activity, measured between light stimuli. Responses were averaged from 5 stimulus presentations. Intensity–response curves were fitted with a sigmoidal dose–response (variable slope), using SigmaPlot 10.0 (SPSS, Chicago, IL). Three parameters were measured from the curve fits: maximum peak response, dynamic range, and light sensitivity. The maximum peak response was the result from the fit of data points. The dynamic range was defined as the range of light intensity that elicited responses between 10% and 90% of the maximum peak response. Drug-induced change in light sensitivity was determined by comparing the light intensity that evoked a half-maximum response before drug application with the light intensity that evoked the same peak response in the presence of the drug. Data in the text are expressed as the mean ± standard deviation. Statistical significance was evaluated using the two-tailed paired Student's t-test (SigmaStat 3.5 software), with P < 0.05 considered significant.
Results
Effects of haloperidol and clozapine on light sensitivity of RGCs
Both ON-center and OFF-center RGCs were sampled. Since the effects of haloperidol and clozapine on intensity–response curves appeared similar, data from both cell types were pooled in the overall analysis to improve statistical power.
Figure 1A shows the effects of haloperidol (0.5 μM) on the intensity–response function of a representative P23H rat RGC. The intensity of light that evoked a half-maximum response before application of haloperidol was 2.97 log units attenuation. With the application of haloperidol, the intensity of light that evoked the same response was 3.19 log units attenuation. By definition, haloperidol increased the light sensitivity of the cell by 0.22 log unit. Haloperidol reduced the maximum peak response of this cell from 191 to 180 spikes/s and increased the dynamic range (the light intensity span that elicits responses between 10% and 90% of the maximum peak response) from 0.79 log unit to 1.0 log unit. Figure 1B shows the effect of haloperidol (0.5 μM) on the light sensitivity of all RGCs (n = 6) examined in this study. All cells showed an increase in light sensitivity. The intensity of light that generated a half-maximum response before application of haloperidol was on average 3.18 ± 0.39 log units attenuation. In the presence of haloperidol, the same light response was obtained at a light intensity of 3.45 ± 0.41 log units attenuation (0.27 log unit lower intensity). The difference of the means was statistically significant (P < 0.001; paired t-test). No statistically significant effect was observed in either the maximum peak response (P = 0.257) or the dynamic range (P = 0.604) of the RGCs. Taken together, the data suggest that haloperidol simply shifts the intensity–response curve to the left.

Effect of haloperidol on light sensitivity of P23H rat RGCs.
Figure 2A shows the effects of clozapine (5 μM) on the intensity–response function of a representative P23H rat RGC. The intensity of light that evoked a half-maximum response before application of clozapine was 4.08 log units attenuation. With the application of clozapine, the intensity of light that evoked the same response was 4.48 log units attenuation. Clozapine had increased the light sensitivity of the cell by 0.40 log unit. Clozapine increased the maximum peak response from 141 to 162 spikes/s and the dynamic range from 0.70 log unit to 0.94 log unit. Figure 2B shows the effect clozapine (5 μM) on the light sensitivity of all RGCs (n = 7) examined in this study. All cells showed an increase in light sensitivity. The intensity of light that generated a half-maximum response before application of clozapine was on average 3.26 ± 0.59 log units attenuation. In the presence of clozapine, the same light response was obtained at a light intensity of 3.56 ± 0.55 log units attenuation (0.30 log unit lower intensity). The difference of the means was statistically significant (P = 0.005; paired t-test). No statistically significant effect was observed in either the maximum peak response (P = 0.333) or the dynamic range (P = 0.457) of the RGCs. Taken together, the data suggest that clozapine simply shifts the intensity–response curve to the left.
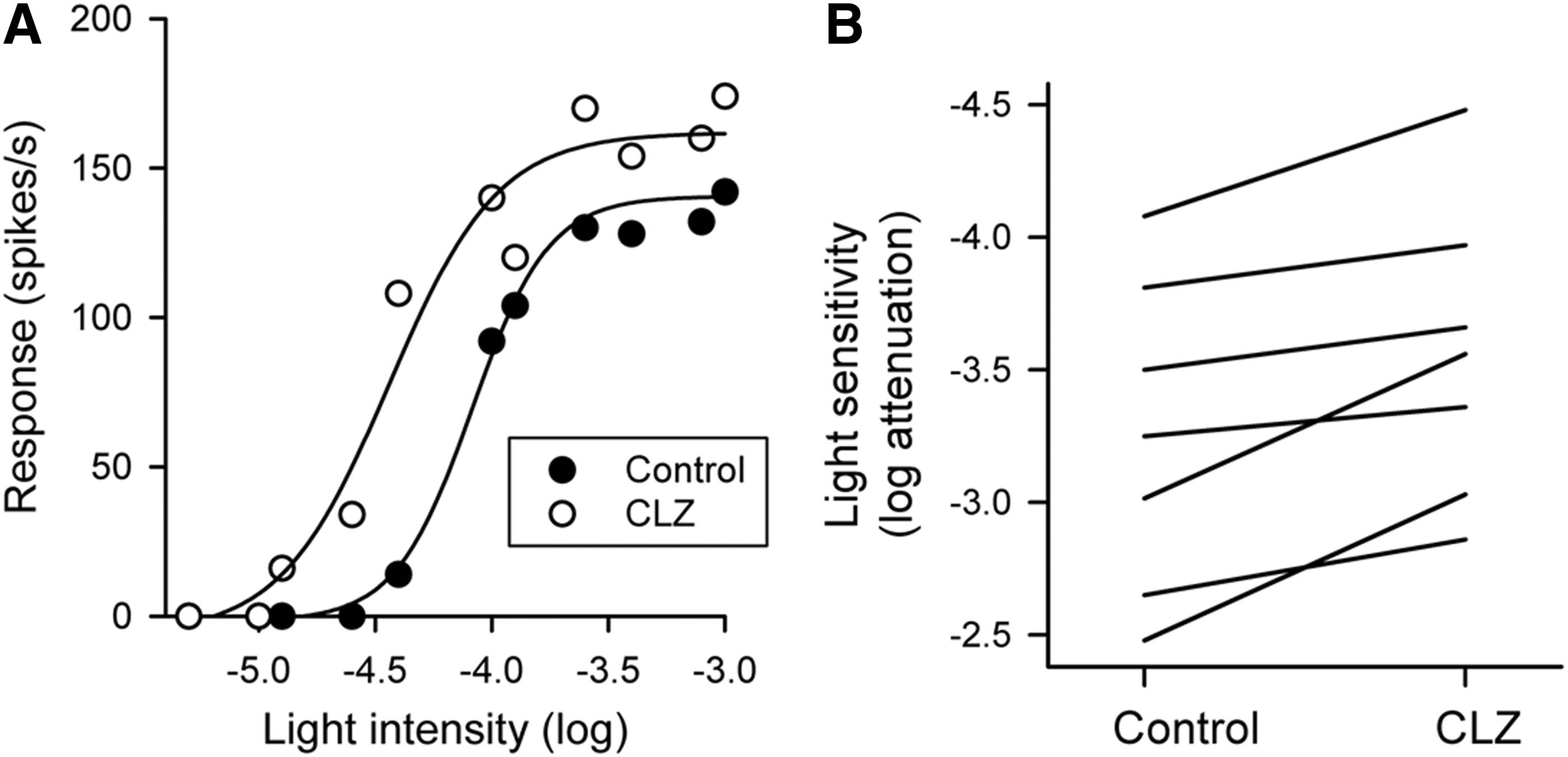
Effect of clozapine on light sensitivity of P23H rat RGCs.
Effects of haloperidol and clozapine on long-latency ON-center RGCs
Previously, I reported on a class of ON-center P23H rat RGCs that respond to light onset of a small spot of light centered over the receptive field with a long-latency ON response. 6 Both sulpiride and eticlopride markedly reduced the ON response and brought out a short-latency OFF response. 6 It is believed that these RGCs might have actually been OFF-center cells early in the disease process. As shown in Fig. 3A, and B haloperidol and clozapine markedly reduced the long-latency ON response of these RGCs to a small spot of light and brought out an OFF response.

Effects of haloperidol and clozapine on light responses of long-latency ON-center P23H rat RGCs. Displayed are spike raster plots and PSTHs of spike activity recorded from 2 RGCs to stimulation with a small (250-μm diameter) spot of light centered over the receptive field. Both haloperidol
Effects of a serotonin 5-HT2A antagonist on light responses of RGCs
Although clozapine can bind to dopamine D2 receptors, it can also bind to serotonin 5-HT2A receptors. In fact, the affinity of clozapine to 5-HT2A receptors is at least 8-fold greater.10,11 Clozapine, as well as other atypical antipsychotic drugs, is thought to mediate some of its therapeutic effects in patients with schizophrenia through 5-HT2A receptor antagonism. 11 I, therefore, examined the possible contribution of 5-HT2A receptors to the effects observed with clozapine using the selective 5-HT2A antagonist MDL 100907. 12
Figure 4A shows the effects of MDL 100907 (1 μM) on the intensity–response function of a representative P23H rat RGC. The intensity of light that evoked a half-maximum response before application of MDL 100907 was 3.35 log units attenuation. With the application of MDL 100907, the intensity of light that evoked the same response was 3.55 log units attenuation. MDL 100907 increased the light sensitivity of the cell by 0.20 log unit. MDL 100907 increased the maximum peak response from 139 to 157 spikes/s and the dynamic range from 0.54 log unit to 1.00 log unit. Figure 4B shows the effect of MDL 100907 (1 μM) on the light sensitivity of all RGCs (n = 8) examined in this study. All cells showed an increase in light sensitivity. The intensity of light that generated a half-maximum response before application of MDL 100907 was on average 2.77 ± 0.40 log units attenuation. In the presence of MDL 100907, the same light response was obtained at a light intensity of 3.03 ± 0.39 log units attenuation (0.26 log unit lower intensity). The difference of the means was statistically significant (P = 0.004; paired t-test). Although no statistically significant effect was observed in the maximum peak response (P = 0.359), there was a statistically significant effect on the dynamic range of the RGCs. On average, the dynamic range of the RGCs increased from 0.88 ± 0.44 log unit to 1.15 ± 0.44 log unit (P = 0.044).

Effects of MDL 100907 on light responses of P23H rat RGCs.
Figure 4C shows the effects of MDL 100907 on the light response of a long-latency ON-center RGC to a small spot of light flashed over center of the receptive field. MDL 100907 (1 μM) markedly reduced the long-latency ON response of this cell and brought out an OFF response. A similar finding was seen in 2 other long-latency ON-center RGCs.
Discussion
Overall, the effects of haloperidol and clozapine on light responses of P23H rat RGCs are comparable to those reported previously with the dopamine D2 receptor antagonists sulpiride and eticlopride. 6 D2 receptor antagonists were found to increase light sensitivity of P23H rat RGCs on average by 0.29 log unit. In this study, haloperidol and clozapine increase light sensitivity of P23H rat RGCs on average by 0.27 and 0.30 log unit, respectively. Another effect of sulpiride and eticlopride in P23H rat retinas is the transformation of abnormal, long-latency ON-center cells into OFF-center cells. These long-latency ON-center RGCs are thought to have been OFF-center cells early in the disease process. 6 I found that both haloperidol and clozapine also transform these long-latency ON-center cells into OFF-center cells. Thus, the effects of haloperidol and clozapine on the light responses of P23H rat RGCs would appear to be due to a blockade of D2 receptors.
While haloperidol is more potent at D2 receptors than 5-HT2A receptors, the converse is true for clozapine. The affinity of clozapine to 5-HT2A receptors is at least 8-fold greater.10,11 Interestingly, I found that the selective 5-HT2A receptor antagonist MDL 100907 increases light sensitivity of P23H rat RGCs and transforms the abnormal, long-latency ON-center cells into OFF-center cells. Although 5-HT2A receptors have been found in the retina,13,14 the functional role of these receptors is not as well understood as D2 receptors. Pharmacological studies in healthy rabbit retinas have shown that 5-HT2 receptor antagonists affect light responses of RGCs.15,16 A prominent effect is a reduction in the responses of RGCs to light onset. The effects of a selective 5-HT2A receptor antagonist on RGCs in healthy retinas have yet to be determined. In healthy Sprague-Dawley rat retinas, D2 receptor antagonists shift the intensity–response curves of RGCs to the right and not to the left as observed in P23H rat retinas. 6 Whether the effects of a selective 5-HT2A receptor antagonist, such as MDL 100907, on light sensitivity of RGCs in healthy retinas are similar to those of a D2 receptor antagonist will need to be determined. Interestingly, the 5-HT2A receptor antagonist sarpogrelate has recently been shown to protect the mouse retina from light-induced retinopathy. 17 This is an important finding since it suggests that blocking 5-HT2A receptors in retinitis pigmentosa retinas may have an additional neuroprotective effect.
In conclusion, the findings in this study suggest that antipsychotic drugs, such as haloperidol and clozapine, may be useful for symptomatic treatment of retinitis pigmentosa. One caveat is that the phenothiazine class of antipsychotic drugs (notably thioridazine) has been documented to cause pigmentary retinopathy with symptoms similar to retinitis pigmentosa 18 and thus should be avoided.
Footnotes
Author Disclosure Statement
No competing financial interests exist.