
Abstract
Abstract
Purposes:
To establish the in silico ocular pharmacokinetic modeling for eye drops, and to simulate the dose regimen for FK962 in human choroid/retinal diseases.
Methods:
Pharmacokinetics for FK962 in vivo was performed by a single instillation of drops containing 0.1% 14C-FK962 in rabbit eyes. Permeation of FK962 across the cornea, sclera, and choroid/retina was measured in vitro. Neurite elongation by FK962 was measured in cultured rat retinal ganglion cells. Parameters from the experimental data were used in an improved in silico model of ocular pharmacokinetics of FK962 in man.
Results:
The mean concentration of FK962 in ocular tissues predicted by in silico modeling was consistent with in vivo results, validating the in silico model. FK962 rapidly penetrated into the anterior and posterior segments of the eye and then diffused into the vitreous body. The in silico pharmacokinetic modeling also predicted that a dose regimen of 0.0054% FK962 twice per day would produce biologically effective concentrations of FK962 in the choroid/retina, where FK962 facilitates rat neurite elongation.
Conclusions:
Our in silico model for ocular pharmacokinetics is useful (1) for predicting drug concentrations in specific ocular tissues after topical instillation, and (2) for suggesting the optimal dose regimens for eye drops. The pharmacodynamics for FK962 produced by this model may be useful for clinical trials against retinal neuropathy.
Introduction
D
Retinal ganglion cells (RGCs) with their axons bundled as the optic nerve pass visual stimuli from the photoreceptor nerves to the brain. 9 Degeneration of the optic nerve leads to visual field loss and blindness seen in glaucoma, axial optic neuritis, Leber's hereditary optic neuropathy, and anterior ischemic optic neuropathy. 10 Some neurotrophic factors may be protective against retinal neuropathy, 11 but topical application may not be acceptable.
FK962 (N-[1-acetylpiperidin-4-yl]-4-fluorobenzamide) may induce glial cell line-derived neurotropic factor (GDNF) in rat cultured trigeminal ganglion and/or glial cells.12,13 In vitro, FK962 caused neurite elongation in trigeminal ganglion cells, possibly through GDNF induction. We previously showed that in silico pharmacokinetic modeling was useful for predicting optimal dosing regimens for FK962 in vivo. 13 However, this model only considered anterior segments and did not predict FK962 concentrations in the posterior segments.
A cylindrical eye model used previously has provided useful data as to how topical instillation of other drugs penetrates and accumulates in specific regions of the eye. Prediction of pharmacokinetics in the choroid/retina was insufficient.14–16 Therefore, the purpose of present study was to establish an in silico ocular pharmacokinetic model for analyzing topical drug distribution in both the posterior and anterior segments of the rabbit eye. FK962 was used to test the model, with the goal of predicting a dosing regimen causing neurite elongation in human RGCs.
Methods
Animals
Male Japanese white rabbits weighing 2.0–2.5 kg (KITAYAMA LABES, Nagano, Japan) provided the cornea, sclera, and sclera/choroid/retina for in vitro permeation studies and were also used in in vivo ocular pharmacokinetic studies. Dutch male rabbits weighing 1.8–2.3 kg (KITAYAMA LABES) were used for an in vivo ocular pharmacokinetic study. Retinas from 7-day-old Sprague-Dawley rats (Japan SLC, Shizuoka, Japan) were used as a source of RGC for culture. All experimental animals were handled in accordance with the Declaration of Helsinki and the Guiding Principles in the Care and Use of Animals (DHEW Publication, NIH 80-23) and in accordance with the tenets of the ARVO Statement for the Use of Animals in Ophthalmic and Vision Research.
Drugs
FK962 was kindly provided by Astellas Pharma, Inc. (Tokyo, Japan). 14C-FK962 was synthesized by GE Healthcare Life Sciences (Tokyo, Japan). The specific activity was 7.59 MBq/mg, and the radiochemical purity was 99.6%.
In vitro permeation study
The in vitro permeation study was performed according to previous reports.17–19 Freshly excised eyes from Japanese white rabbits were used. The eyes were opened circumferentially ∼3 mm behind the limbus, and the corneas were carefully excised. Then, the vitreous humor was separated from retina/choroid/sclera tissues, and retina/choroid/sclera tissues were circularly dissected. In the case of scleral tissue penetrations, retina and choroid were carefully peeled from the sclera.
Cornea, sclera, and sclera/choroid/retina samples were mounted side-by-side in Ussing diffusion chambers (Vidrex, Fukuoka, Japan). Test solution (3.6 mL) was added to the donor chamber, while receptor buffer solution (3.6 mL) was present in the receptor chamber. The test solution was 0.1% FK962 in isotonic phosphate buffer (pH 7.0), and the receptor buffer solution was composed of 0.013% calcium chloride, 0.04% potassium chloride, 0.02% magnesium sulfate, 0.019% sodium dihydrogen phosphate dehydrate, 0.787% sodium chloride, and 0.1% glucose (pH 7.2). 18 The 2 chambers were separated by the cornea, sclera, or sclera/choroid/retina. In vitro permeation studies were started within 30 min after eye excision. At predetermined time intervals, 200 μL samples were collected from the receptor chamber and replaced with fresh buffer. At the termination of permeation studies, the tissues appeared in good condition. FK962 in the receptor solution was measured according to a previously reported method 13 at 225 nm using high-performance liquid chromatography (HPLC, LC-10AD; Shimadzu, Kyoto, Japan) equipped with reverse-phase Inertsil ODS-3 column (inner diameter, 4.6 mm; length, 250 mm; pore size, 5 μm; GL Sciences, Tokyo, Japan). The mobile phase consisted of 35/65 (v/v) mixture of methanol/5 mM disodium hydrogen phosphate in 12-water (pH 6.0). The lower quantitative limit for FK962 allowed measurement of samples from the sclera/choroid/retina only at 60 min after permeation. FK962 was eluted at 13.2 ± 0.4 min after sample injection. The linearity of the regression line using standard solutions was confirmed in the concentration range of 0.05–20 μg/mL (r2 > 0.999).
Permeation profiles across the cornea, sclera, and sclera/choroid/retina were constructed by plotting the accumulation of permeated FK962 versus time. The permeation rate (dQ/dt) and time lag (td) were calculated from the slope and x-intercept of the linear regression line, respectively. Permeability (P), diffusion (D), and partition (K) coefficients in the cornea and sclera were calculated based on Fick's second law of diffusion:
where C is the drug concentration in the donor solution, and L is the thickness of the cornea or the sclera. Diffusion and partition coefficients in the choroid/retina were determined from the permeation rate and the time lag based on a bilayer membrane model,19–21
although there is some variation in the time lag.
where the subscripts, S, SCR, and CR, refer to the sclera, sclera/choroid/retina, and choroid/retina.
In silico ocular pharmacokinetic model
The new models were modifications of the cylindrical eye model based on Fick's second law of diffusion,14–16 considering the anatomical geometry of rabbit eye (Fig. 1A) and human eye (Fig. 1B).22–30 In these models, the drug was assumed to diffuse by 2 pathways across the corneal and the scleral surfaces. The drug concentrations in the ocular tissues were computed by solving the equations below with previously reported parameters.14–16
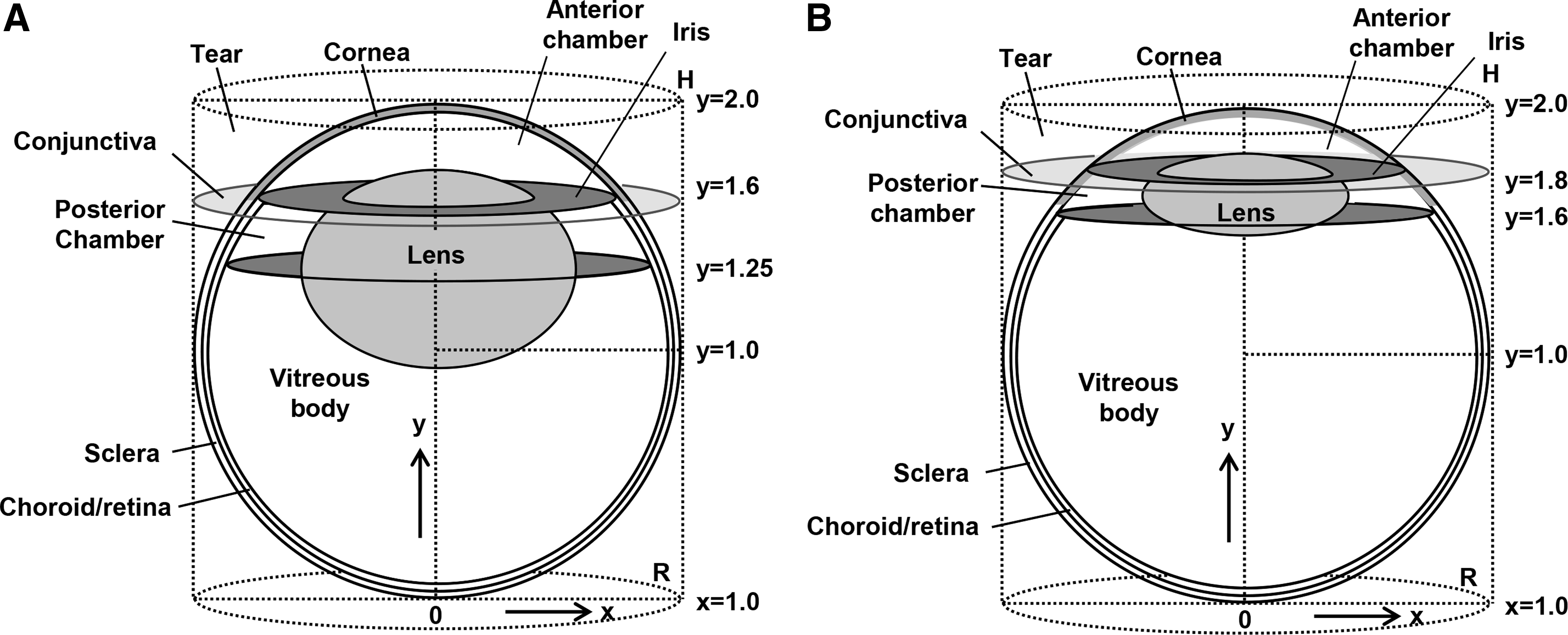
Current cylindrical model for ocular pharmacokinetic studies in rabbit
The tear concentrations were shown for Equation (6):
where C0 is the initial concentration of drug in the tear fluid, and k1 and k2 are the elimination rate constant and the absorption rate constant of the drug in the tear fluid (s−1). The initial concentration of drug in tear fluid was instantaneously diluted to 45% by tear fluid after instillation.15,16 The elimination and the absorption rate constants for FK962 in tear fluid were determined from concentration versus time profiles of FK962 from the present in vivo study using linear regression analysis by the least squares method; k1 = 2.3 × 10−3, k2 = 1.6 × 10−5, and the constant B = 0.001. The same elimination rate constant in tear fluid was used for human and rabbit because the instilled solution mostly drains from the nasolacrimal duct, 31 and consequently, the tear turnover has only a minor role in the removal of instilled solution from the ocular surface.
The drug concentration in each eye tissue is described in Equation (7), which was solved by the method of line.
21
D is the diffusion coefficient in the eye, R (x, y, t) is the metabolism and elimination rate. We did not consider binding or metabolism of FK962 in the eyes, because only minor influences of these factors were confirmed in the present in vitro and in vivo studies.
The initial and boundary conditions are described in Equations (8)–(12).
R and H are the effective radius and height of the eye globe. D is the diffusion coefficient in the eye tissues (cm2/s), and D0 is the diffusion coefficient in the space between the outside cylinder and the inscribed sphere. K is the membrane partition coefficient, L is the membrane thickness, and the subscripts, c and s, refer to the corneal membrane, the scleral membrane, respectively. The diffusion coefficients of the cornea, sclera, and choroid/retina were determined from the in vitro permeation experiments (Table 1). Other diffusion coefficients in ocular tissues and the cylindrical eye model were determined from reference values (Table 2).14–17 The effective diffusion coefficient in the vitreous body and the lens was previously determined from in vitro permeation experiments,14–17 and the diffusion coefficient in the vitreous body was the same order of magnitude as in water. 17 The diffusion coefficient in the space between the circumscribed cylinder and an inscribed sphere (Fig. 1) was assumed to be 1,000 times greater than that in the vitreous body (Table 2). This is because the diffusion resistance in the space between the circumscribed cylinder and an inscribed sphere was necessarily set as negligible in our cylindrical model.14–16 The effective diffusion coefficient in the tear fluid was assumed to be 5 times greater than that in the space between the circumscribed cylinder and an inscribed sphere, since the topically instilled drug was instantaneously spread over the corneal surface and was perfectly mixed by blinking and tear flow. The effective diffusion coefficient in the conjunctiva was assumed to be 2 times greater than that in sclera since conjunctival permeability was about 2 times higher than scleral permeability. 32 The effective diffusion coefficient in the anterior and posterior chamber was assumed to be 10 times greater than that in the vitreous body since the aqueous humor has convection flow, which enhances overall mixing of drug molecules.14–16 We assumed that the effective diffusion coefficients in the anterior and posterior chamber of rabbit and human are similar because the aqueous turnover rate is 1%–2%/min in both species. 33 The elimination rate of FK962 from aqueous humor was calculated to be 9.6 × 10−4 s−1 based on a previous report in rabbit.34–36 In contrast to aqueous humor, turnover of vitreous body was not taken into account. This was done because the vitreous body is a static unstirred fluid and so that transport of small drug or solute in the vitreous humor occurs primarily via diffusion. 37 Choroidal blood flow and conjunctival blood and lymphatic flows were also not incorporated in our cylindrical model.
Melanin affinity of FK962 in vitro
In vitro melanin binding study was performed according to a modified previous report. 38 A suspension of 2 mg/mL natural melanin from Sepia officinalis (Sigma-Aldrich, St. Louis, MO) was prepared in 20 mM phosphate buffer (pH 7.4). FK962 ranging from 2.5 to 500 μM in 20 mM phosphate buffer (0.5 mL) was mixed with melanin suspension (0.5 mL) and incubated for 24 h in an incubator at 37°C with shaking at 200 rpm. After incubation and to separate the melanin granules, the samples were centrifuged for 30 min at 12,000 rpm, and the supernatant was collected (free FK962). Then, 1 mL of fresh 20 mM phosphate buffer (pH 7.4) was added to the pellet containing the melanin-FK962 complex and incubated for 24 h in an incubator at 37°C with shaking at 200 rpm. The samples were centrifuged for 30 min at 12,000 rpm, and the supernatant was collected (dissociated FK962). The supernatants were filtered by 0.22 μm pore membrane filter, and FK962 was measured by HPLC as described above. The experiments at each FK962 concentration were performed in triplicate. As a control, the drug alone was incubated in the buffer (total FK962). Melanin-bound FK962 (%) and FK962 dissociated after binding (%) were calculated as [(total FK962 − free FK962)/total FK962] × 100 and [dissociated FK962/(total FK962 − free FK962)] × 100, respectively.
In vivo pharmacokinetic study
Pigmented rabbits received 0.1% 14C-FK962 ophthalmic solution (30 μL/eye) onto the upper bulbar conjunctiva in both eyes. Preliminary experiments after unilateral instillation observed lower concentrations of FK962 in the choroid/retina of the contralateral eye. This protocol also reduced animal numbers in consideration of experimental animal welfare. The rabbits were then anesthetized with 5% pentobarbital administered into the auricular vein at various times. To estimate pharmacokinetic differences between pigmented and nonpigmented eyes, albino rabbits were also treated and anesthetized 0.5 and 2 h after 14C-FK962 instillation. After tears were collected by inserting filter paper strips into the conjunctival sac, the rabbits were euthanized by exsanguination from the abdominal aorta. The inside of the conjunctival sac was washed with 10 mL of physiological saline, and the eyes were excised. Corneal epithelia were collected with a surgical knife and aqueous humors were obtained with a syringe and needle. After sampling the conjunctivas, eye globes were immediately frozen in dry-ice/acetone bath. The eye globes were then separated at the equatorial region into anterior and posterior poles. The corneal stroma/endothelium, lens, iris/ciliary body, and anterior choroid/retina and sclera were collected from anterior poles. The vitreous body and posterior choroid/retina and sclera were obtained from posterior poles. After drying the samples in combustion vials, radioactivity in each vial was measured for 5 min with a liquid scintillation counter (TRI-CARB 2500TR or 2700 TR; PerkinElmer, Kanagawa, Japan) equipped with a quenching correction system using tSIE (transformed Spectral Index of External standard). After subtracting background, the radioactivity in each sample was expressed as net counts. Metabolism of 14C-FK962 in eye was not determined. Instead, metabolic stability of FK962 was tested by incubation for 24 h of the chopped cornea, conjunctiva, iris/ciliary body, and choroid/retina. We found that more than 98% of FK962 was detected in the unchanged form. Therefore, measured radioactivity was considered intact 14C-FK962 and not metabolites or breakdown products.
Rat RGC culture and measurement of neurite length
RGCs from rats were cultured with a modified method previously reported.
39
Retinas were dissected from rat eyes and rinsed in cold Hanks' balanced salt solution (HBSS; Thermo Fisher Scientific). Retinal cells were dissociated with the neural tissue dissociation kit for postnatal neurons (Miltenyi Biotec, Bergisch Gladbach, Germany), and RGCs were isolated according to the manufacturer's protocol in the Retinal Ganglion Cell Isolation Kits (Miltenyi Biotec). To minimize variability, isolated RGCs, and other retinal cells excluding RGC/microglia/endothelial cells, were seeded together at 1 × 104 cells and 5 × 105 cells/well, respectively, on μ-Plate 96 well (ibid, Martinsried, Germany) coated with poly-
Cells were fixed for 20 min in 2% formaldehyde. To identify RGCs and their neurites, fixed cells were double labeled by incubating with primary antibodies against 200 kDa neurofilament (NF200; Abcam, Cambridge, UK) and brain-specific homeobox/POU domain protein 3A (BRN3A; Merck, Darmstadt, Germany). Astrocyte and Müller cells were identified with the anti-glial fibrillary acidic protein (GFAP) antibody (Merck) and anti-vimentin antibodies (Cell Signaling), respectively. After washing with phosphate-buffered saline (PBS) and to visualize nuclei, cells were incubated with secondary antibodies conjugated to Alexa Fluor-488 and Alexa Fluor-568 (1:1,000; Thermo Fisher Scientific), together with 4′,6-diamidino-2-phenylindole dihydrochloride (DAPI; Thermo Fisher Scientific). Images were observed with a fluorescence microscope (IX71; Olympus, Tokyo, Japan) and digitalized with a charge-coupled device (CCD) camera on a personal computer.
Measurement of neurite length was performed with a modification of the method reported by Lingor et al. 40 The length of neurites extending from NF200 and BRN3A double-positive RGC bodies was measured in 4 areas/well with Image-Pro Premier (Nippon Roper, Tokyo, Japan). Total length of neurite/RGC was calculated in each well and was averaged for 6 wells. Cultures from different rats were performed 3 times, and data are expressed as mean ± standard deviation (SD).
Statistical differences (P < 0.05) were determined by Dunnett's multiple comparison test with randomized blocks (JMP statistical software; SAS Institute).
Results
In vitro permeation of FK962 and calculated parameters for the in silico model
FK962 permeated from the donor chamber, across the cornea, sclera, and sclera/choroid/retina, and accumulated in the receptor chamber with increasing incubation times (Fig. 2). The concentration of FK962 in the donor chamber was virtually constant (data not shown), because of a larger amount of FK962 in the donor chamber compared to that permeated into the receptor chamber. All permeation profiles were linear (Fig. 2) because drug permeation is concentration dependent. Sclera allowed more accumulated FK962 compared to sclera/choroid/retina, indicating that choroid/retina was a permeability barrier to FK962. Cornea was also a moderate barrier to FK962 permeation. These plots were used to determine lag times, permeation rates, and permeability for FK962. The calculated permeation rate, lag time, and permeability for the cornea were 41.4 ± 9.4 μg/cm2/h, 0.24 ± 0.03 h, and 1.2 × 10−5 ± 2.7 × 10−6 cm/s, respectively. In sclera, the permeation rate, lag time, and permeability were 90.8 ± 9.9 μg/cm2/h, 0.12 ± 0.07 h, and 2.5 × 10−5 ± 2.8 × 10−6 cm/s, respectively. In the sclera/choroid/retina, the permeation rate and permeability decreased to 22.8 ± 3.5 μg/cm2/h and 6.4 × 10−6 ± 1.0 × 10−6 cm/s, respectively, and the lag time increased to 1.10 ± 0.29 h. These calculated values allowed determination of the diffusion and partition coefficients necessary for in silico pharmacokinetic modeling (Table 1). These parameters and those previously reported (Tables 2 and 3) were used in ocular pharmacokinetic modeling presented below.
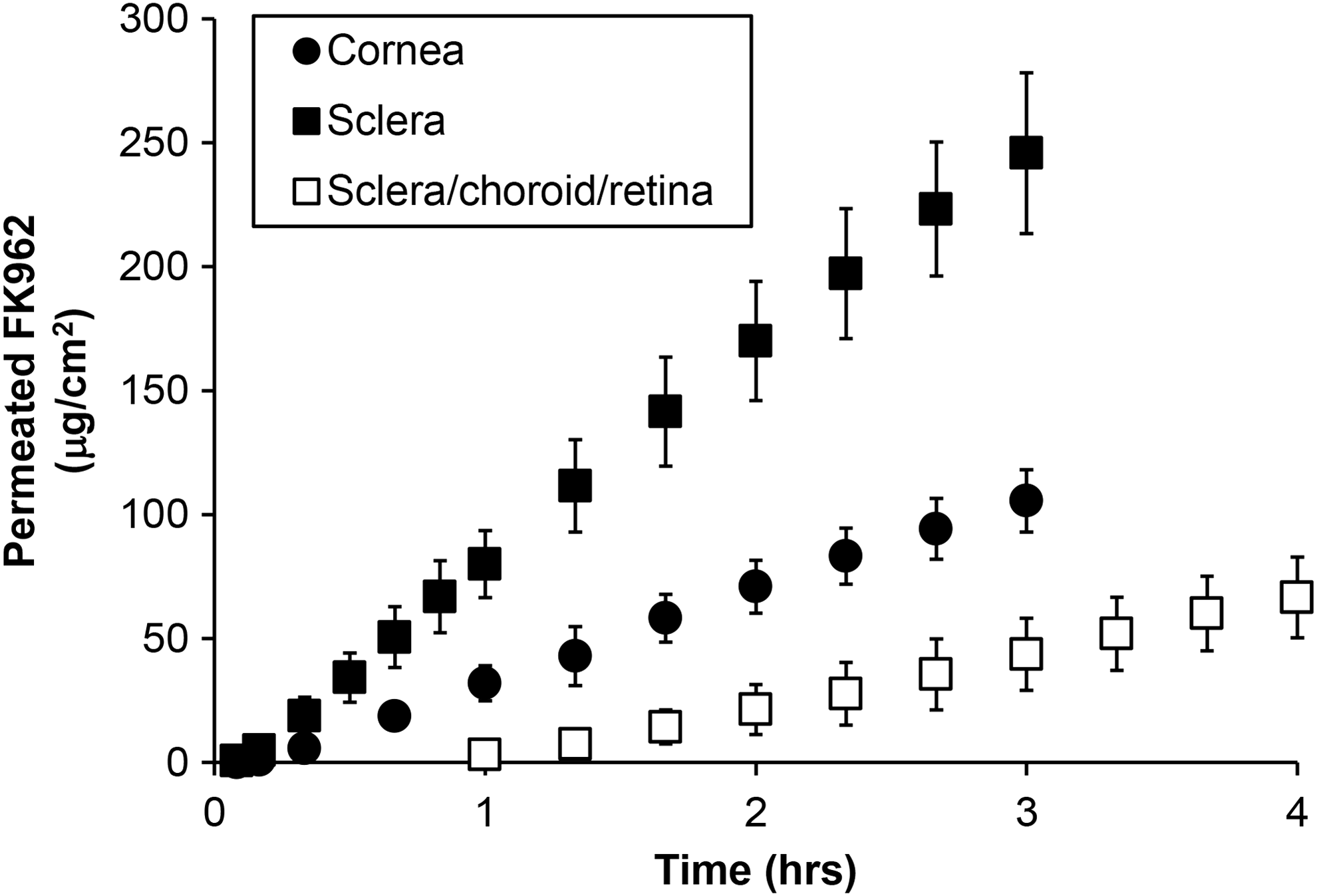
In vitro FK962 permeation across cornea (closed circles), sclera (closed squares), and sclera/choroid/retina (open squares). Each point is the mean ± SD (n = 3 or 4). SD, standard deviation.
Simulated ocular uptake after instillation of FK962
The distribution of FK962 in the eye was visualized after a single, virtual instillation of 0.1% FK962 (Fig. 3). Five minutes after instillation, high concentrations of FK962 were observed in the tear fluid. By 0.5 h, FK962 had penetrated into the anterior and posterior segments (Fig. 3; 5 min, 0.5 h). With increased time, FK962 further gradually penetrated into the posterior and anterior segments, even though the FK962 level in the vitreous body was very low (Fig. 3; 0.5 and 1 h). This suggested some penetration of FK962 into the choroid/retina via the conjunctival/scleral pathway. After 2 h, the level of FK962 gradually decreased in most tissues, but more FK962 had diffused into the vitreous body (Fig. 3; 2 and 4 h).
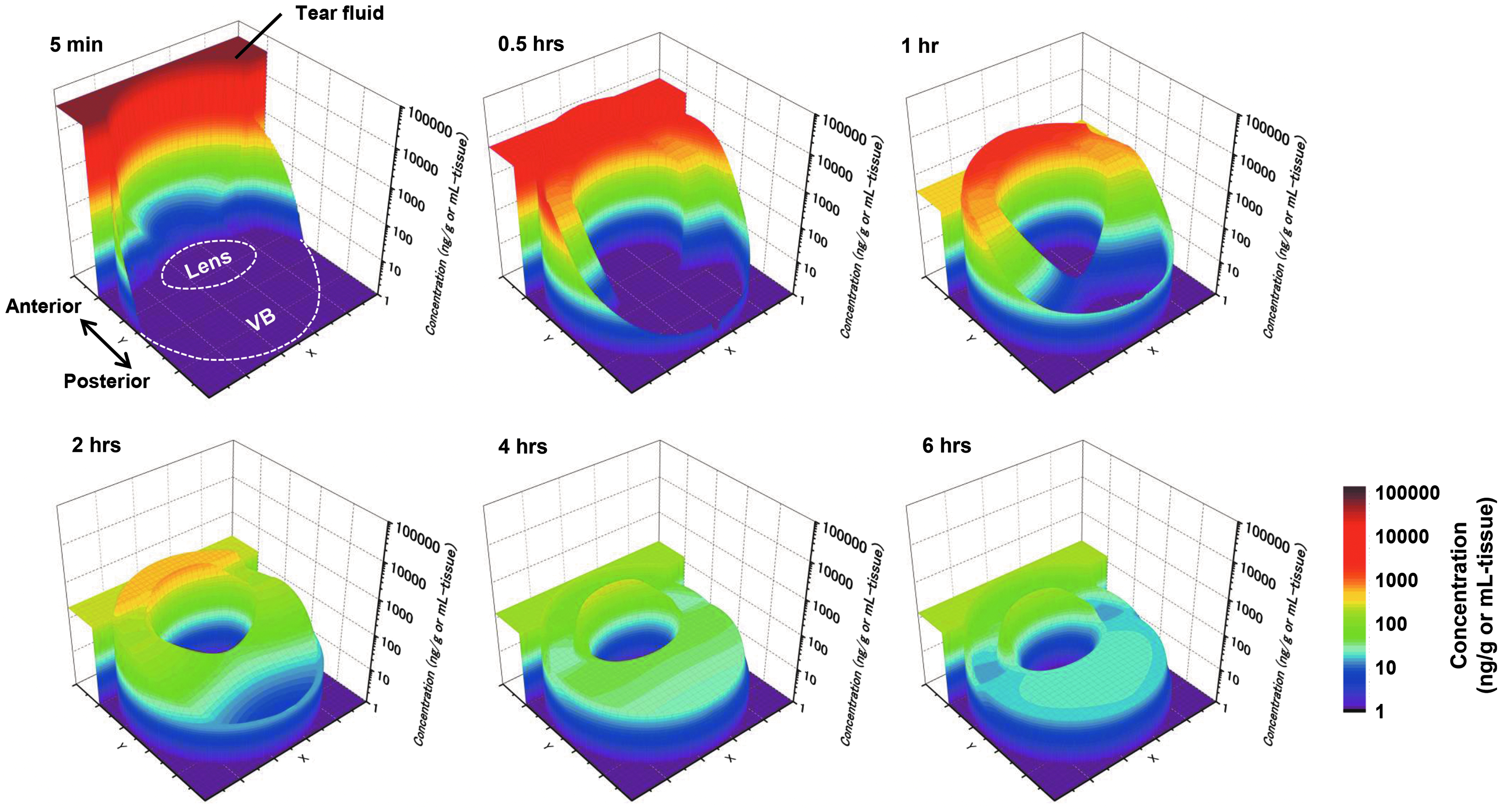
FK962 concentrations after 1 virtual instillation of FK962 into rabbit eyes, showing permeation of FK962 from the periphery to the center of the eye. Pseudocolor (highest to lowest concentration): red to blue. VB, vitreous body.
Melanin affinity of FK962 in vitro
Amounts of FK962 bound to melanin were 3.9 to 9.9% at the concentrations tested, and over 85% of the FK962 dissociated after binding to melanin (Table 4). This suggested that binding capacity and intensity of FK962 to melanin were low and weak.
Data are mean ± SD (n = 3).
SD, standard deviation.
Verification of simulated results by in vivo pharmacokinetics
After a single topical instillation of 14C-FK962 into pigmented rabbit eyes, FK962 was rapidly distributed throughout ocular tissues with a time of maximum concentration (Tmax) of 0.25–0.5 h (Fig. 4, closed squares). The concentration was highest in tear fluid, followed in descending order by the corneal epithelium, iris/ciliary body, corneal stroma/endothelium, aqueous humor, anterior sclera, anterior choroid/retina, posterior sclera, vitreous body, posterior choroid/retina, and lens. Although only 2 time points (near Tmax at 0.5 h and declining at 2 h) were measured in albino rabbits, similar concentrations were observed as in pigmented rabbits (Fig. 4, open circles vs. closed squares). Also, the elimination profiles from melanin-containing tissues were similar to those from nonpigmented tissues in pigmented rabbits. These data suggested low melanin binding by FK962 and agreed with the in vitro results above. Next, concentrations and kinetics for FK962 in ocular tissues from the in silico experiments (Fig. 4, solid lines) were compared to in vivo studies (closed squares). These pharmacokinetic profiles were very similar to the profiles from the in silico experiments when the data were expressed as mean concentrations in each tissue (Fig. 4). Thus, the in vivo experiments verified our in silico pharmacokinetic model.
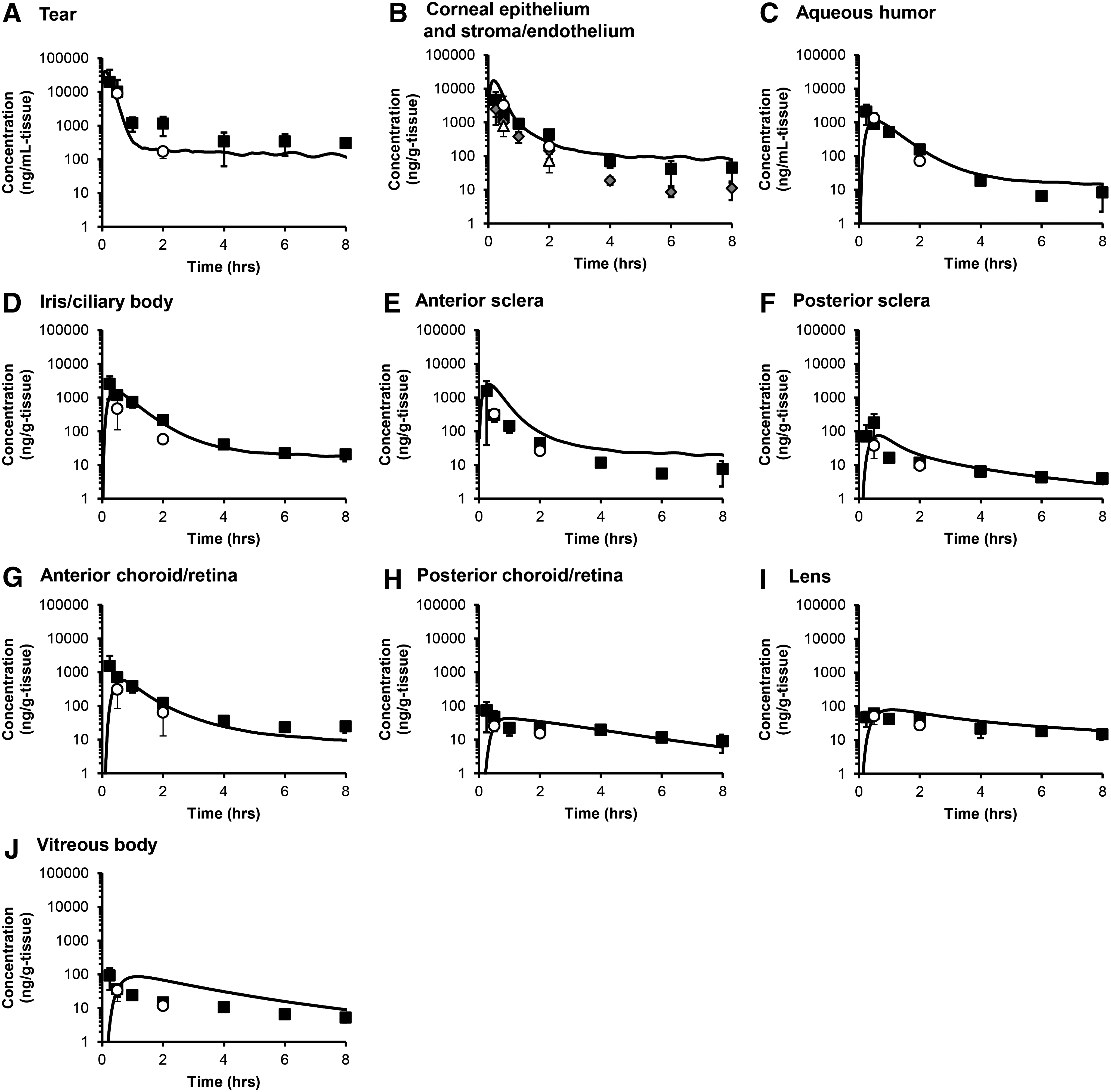
Pharmacokinetics of FK962 in silico (solid line) compared to in vivo instillation of 0.1% 14C-FK962 in albino (open circles) and pigmented rabbits (closed squares).
Effect of FK962 on neurite elongation in rat RGCs
In primary cultures of rat RGCs, we observed NF200/BRN3A double-positive RGCs (Fig. 5A, left), GFAP-positive astrocytes (center), and vimentin-positive Müller cells (right). Smaller numbers of amacrine, photoreceptor, horizontal, and bipolar cells were also present in these cultures (data not shown). After 24 h of culture, elongated neurites projected from the RGCs in control cultures (Fig. 5B, left), but more extended neurites were observed in the FK962-treated group (Fig. 5B, right). Treatment with various levels of FK962 produced a bell-shaped dose–response curve, with a significant, maximally extended neurite length at 10−9 M (Fig. 5C). The depressing effect of FK962 at higher concentrations was not due to toxicity, because cell numbers were the same at all levels of FK962 (data not shown).

Prediction of the optimum human dosing regimen from in silico modeling
FK962 is a candidate drug against retinal neuropathy. Therefore, we used our verified in silico pharmacokinetic model to determine the optimal dose for topical instillation of FK962 in human eyes. For effective stimulation of neurite elongation in the human eye, we assumed that the final concentration of FK962 in human choroid/retina needed to be in the range of 5 × 10−10 to 5 × 10−9 M (shaded bar in Fig. 6A, B). This was based on the data showing that 10−9 M FK962 in cultured rat RGCs caused significant neurite elongation (Fig. 5C).
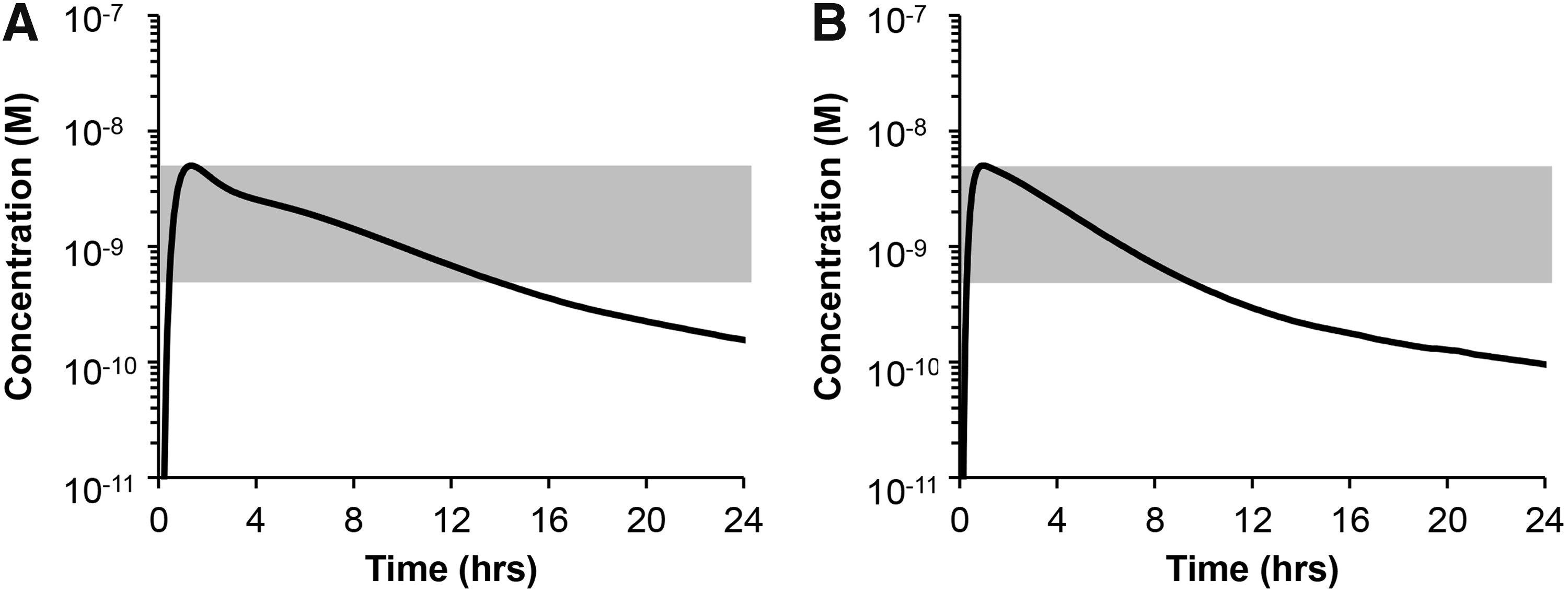
Simulated FK962 concentrations in the posterior choroid/retina after a single instillation of 0.0054% FK962 in human
Pharmacokinetic modeling simulating application of 0.0054% FK962 instilled into human eye showed that FK962 immediately increased in the posterior choroid/retina, peaked 5 × 10−9 M at 1.3 h, and then gradually decreased (Fig. 6A). Between 0.4 and 13 h after instillation, FK962 in the posterior choroid/retina remained in the effective range of 5 × 10−10 to 5 × 10−9 M (Fig. 6A, shaded area). The pharmacokinetic profile in rabbit (Fig. 6B) was similar in human, except that the optimum effective dose for rabbit was 0.0031%, less than that for human.
Discussion
In the present study, we established an in silico ocular pharmacokinetic model for instillation of drugs in rabbit and human eyes. The rabbit model was verified by its similarity to an in vivo model using instilled 14C-FK962. The in silico model predicted that a single topical instillation of 0.0054% FK962 in human eyes would elevate FK962 levels in choroid/retina high enough to promote neurite elongation for ∼12 h.
The present in silico ocular pharmacokinetic model has been significantly improved by several methods. For example, a previous cylindrical eye model for ocular pharmacokinetics had been used to predict drug distribution in ocular tissues after virtual instillation.14–16 Although ocular tissues showed a variety of diffusion and partition coefficients, this previous model did not determine the parameters for sclera and choroid/retina, and the concentrations in these tissues were not predicted. To improve the model, we first measured drug permeation in vitro across the cornea, sclera, and sclera/choroid/retina and calculated diffusion and partition constants used in our in silico modeling. To allow for species-specific predictions, we further added values for tissue geometry that were specific for rabbit (Fig. 1A) and human eyes (Fig. 1B). Accurate modeling also requires drug-specific permeation parameters that are influenced by molecular weight, molecular radius, and lipophilicity.38,41 Our in silico pharmacokinetic profile used the calculated in vitro permeation parameters specific for FK962 that were verified with in vivo data in rabbits. However, the simulated concentration in the vitreous body and the anterior sclera deviated slightly from the in vivo results. A reason for this deviation is probably due to the use of reference values for diffusion coefficients in the vitreous body, anterior and posterior chambers, and conjunctiva; and for elimination rates in the anterior and posterior chambers. Calculation and use of our diffusion coefficients and elimination rates for FK962 may improve fitting in the vitreous body and anterior sclera.
Binding of drugs to melanin is known to affect drug efficacies, and melanin is presented in the iris, ciliary body, sclera, choroid, and retina. The present study in vitro showed minor binding activity of FK962, and most of FK962 dissociated even FK962 bound to melanin. Also, similar pharmacokinetic profiles were observed in albino and pigmented rabbits, suggesting that affinity of FK962 to melanin was low. Therefore, binding of FK962 to melanin was not considered in our cylindrical eye model. Protein binding may also affect drug distribution in ocular tissues. This factor in the tear and aqueous humor is not expected to contribute significantly in normal eyes because the protein concentrations in the tear and aqueous humor are negligible compared to that in plasma. 31 Therefore, protein binding in the tear and aqueous humor is not thought to be a major factor in pharmacokinetics in normal eyes. 31 Also, in vivo ocular pharmacokinetic studies of FK962 showed that elimination profiles in ocular tissues were almost same as that in tear and aqueous humor. Consequently, we considered that FK962 bound to proteins at low level, and protein binding was not incorporated in in silico analysis of FK962. When simulating pharmacokinetic of other drugs that are indeed bound to proteins, the model would then need to account for protein binding affects.
Our cylindrical model accounted for drug elimination from ocular tissues [Eq. (7)], but not from the blood flow from choroid and conjunctiva or the lymphatic flow. When our in silico modeling incorporated elimination from conjunctival blood flow based on a previous report, 42 the pharmacokinetic profiles (not shown) were similar to the present modeling data. Regarding the choroidal blood flow, contradictions exist. Some studies reported important flow for drug elimination from the choroid, 43 which was not observed in other reports.44,45 A detailed understanding of the influence of these flows on the clearance of topical drugs is still unclear. However, the pharmacokinetic profiles in silico were very similar to the profiles from the in vivo experiments (Fig. 4), suggesting that these flows may only minimally contribute to FK962 elimination. When compounds known to be eliminated significantly by these flows are tested, incorporation of their elimination rates would help refine the model.
Our model also has some limitations. For example, the choroid and retina were treated as a single tissue because of the difficulty in separating and isolating intact choroid and retina. When experimental techniques and computational capability improve, concentration of drugs will need to be predicted separately for the choroid and retina.
FK962 permeated the sclera>cornea>sclera/choroid/retina in our in vitro study (Fig. 2). The diffusion coefficient of FK962 was approximately 3–5 times higher in the sclera than in the cornea and choroid/retina. These data suggested that both the cornea and choroid/retina were permeability barriers to FK962. The transcorneal route has been traditionally recognized as a pathway for ophthalmic drugs to gain access to the intraocular tissues. A route crossing the conjunctiva and sclera may also significantly contribute to drug penetration.4,6,46 Our in silico modeling showed wide distribution and sequential changes in drug concentrations (Fig. 3). FK962 reached the anterior segment after only short time postvirtual instillation (Fig. 3; 5 min and 0.5 h), followed by penetration into the posterior segment (Fig. 3; 0.5 and 1 h). FK962 then diffused into the vitreous body (Fig. 3; 2 and 4 h). These data suggested that FK962 initially penetrated into the choroid/retina mainly by the transscleral pathway and only secondarily by a minor transcorneal pathway. A histochemical study with radiolabeled FK962 in vivo is expected to produce more specific details.
FK962 enhanced neurite elongation of RGCs significantly only at 10−9 M (Fig. 5). The bell-shaped dose–response curve of FK962 was observed in neurite elongation of trigeminal ganglion cells in rat, rabbit, and monkey.11,12,47 The mechanism of neurite elongation is suggested to occur via induction of GDNF in rat and monkey.12,47 FK962 also stimulates somatostatin release from rat hippocampal slices and increases somatostatin-induced neurite elongation in rat cerebellar granule cells. 48 Many neurotrophic factors and peptides causing axonal elongation show bell-shaped dose responses. 49 Although the precise mechanism for the negative influence of the higher doses is not established yet, the higher doses are thought to cause negative feedback. 50 Since the effective doses of FK962 were narrow, a prediction of optimum concentration is useful for in vivo animal experiment and human clinical research.
Our in silico pharmacokinetic modeling was also useful for predicting dosage and administration in human eyes. Tissue concentration of FK962 producing neurite elongation in vitro was retained for ∼12 h in the choroid/retina after a single virtual instillation of topical 0.0054% FK962 (Fig. 6A). Thus, topical instillation of 0.0054% FK962 twice a day was suggested as a dosing regimen for humans. This prediction was based on parameters calculated from rabbit in vitro permeation experiments. No significant difference was reported in drug permeation in ocular tissues between rabbit and human, 24 suggesting that our in silico prediction was acceptable. A limitation of the prediction was that biologically effective tissue concentrations of FK962 were based on rat RGC neurite elongation studies. Human RGCs were not currently available in our laboratory. When induced pluripotent stem cell-derived human RGCs become available, data from such cells will replace the rat data.
Footnotes
Acknowledgments
The authors thank Dr. Kakuji Tojo (Kyushu Institute of Technology, Fukuoka, Japan) for helpful comments for computer simulation, and Mr. Kenichiro Hayashi (formerly Mitsubishi Chemical Medience Corporation, Tokyo, Japan) and coworkers for the in vivo pharmacokinetic study.
Author Disclosure Statement
Dr. Shearer is a paid consultant for Senju Pharmaceutical Co., Ltd., a company that may have a commercial interest in the results of this research and technology. Dr. Azuma is an employee of Senju Pharmaceutical Co., Ltd. This potential conflict of interest was reviewed, and a management plan approved by the OHSU Conflict of Interest in Research Committee was implemented. All other authors have no competing financial interests.