
Abstract
Abstract
Purpose:
Panax Notoginseng, a traditional Chinese medicine, is known as an anti-inflammatory herb. However, the molecular mechanism by which it controls helper T cell mediated immune responses is largely unknown.
Methods:
Naive CD4+ T cells isolated from healthy donors, patients with Behcet's disease, and C57BL/6 mice were polarized into Th1, Th17, and Treg cells. Proliferation and cytokine expression were measured in these cells with the presence or absence of Panax Notoginseng saponins (PNS). Genomewide expression profiles of Th1, Th17, and Treg cells were assessed using Affymetrix microarray analysis.
Results:
We found that PNS control the proliferation and differentiation of Th17 cells by globally downregulating the expression of inflammatory cytokines and cell cycle genes.
Conclusions:
These findings demonstrated that PNS function as an anti-inflammatory agent through directly targeting Th17 cell mediated immune response.
Introduction
P
More than 50 different Panax Notoginseng saponins (PNS), including Rb1, Rg1, and Rh1, have been identified as the major pharmacologically active ingredients among the ∼200 chemical compounds isolated from Panax Notoginseng.2,7 Previous studies have suggested an anti-inflammatory role of PNS in vivo and in vitro. 8 Rb1 suppresses LPS-induced production of TNF-α and IL-6 in cultured macrophages in a dose-dependent manner, while Rh1 potently inhibits histamine release from mast cells and the IgE-mediated passive cutaneous anaphylaxis reaction in vivo.9,10 However, the immunomodulatory effects of PNS have not been fully characterized.
Behcet's disease (BD), an autoinflammatory disease mediated by both Th17 and Th1 cells, is a chronic recurrent inflammatory disorder of unknown etiology involving multiple organs. 11 Systemic vasculitis, frequent oral and genital ulcers, uveitis, and skin lesions are the most common manifestations. 12 In addition, cardiac or neurological manifestations can be seen in advanced BD patients with a geographical preference. 13 Immunosuppressive therapies remain the current standard treatment; however, a significant amount of patients do not respond to current therapies, 14 warranting the development of novel methods for treating BD.
In this study, we investigate the molecular mechanism underlying the anti-inflammatory effects of PNS. Our data suggest that PNS suppress both proliferation and differentiation of Th17 cells in cultures of T cells from peripheral blood of healthy volunteers and BD patients. These findings support the potential benefits of PNS in novel therapies for BD.
Methods
Human subjects and animals
Healthy volunteers and BD patients were recruited at the clinics of Zhongshan Ophthalmic Center, Sun Yat-sen University. The diagnosis of BD was made according to the diagnostic criteria designed by the International Study Group for BD. 15 To exclude the confounding effects of different medicines used by BD patients, only patients with inactive disease and free of drug usage were included in current study. All protocols were approved by institutional review boards, and written informed consents were provided by all subjects before enrollment. The animal study was approved by Ethics Committee in Animal Experimentation of Zhongshan Ophthalmic Center, Sun Yat-sen University and was performed in accordance with the tenets of the Declaration of Helsinki.
Panax Notoginseng saponins
The PNS used in current study were an aqueous extract from the stem, leaf, and flower of Panax Notoginseng whole plants grown in the Wenshan region of Yunnan Province in China and purified by high-performance liquid chromatography. It was purchased from Yunnan Hongyan Bioengineering Technology Co., Ltd., China and resuspended into solutions at various concentrations using phosphate-buffered saline.
Isolation and culture of CD4+ T cells
Human peripheral blood mononuclear cells (PBMCs) were isolated from fresh heparinized blood by Ficoll-Hypaque density gradient centrifugation. CD4+ T cells were purified from PBMCs using autoMACS Pro and Naive CD4+ T cell Isolation Kits (Miltenyi) according to the manufacturer's instruction. The naive CD4+ T cells were stimulated with the plate-bound 3 μg/mL anti-CD3 (Dako) and the soluble 1 μg/mL anti-CD28 (Dako) antibodies, as well as other polarizing cytokines/antibodies for 72 h. For Th0 cell culture, no polarizing cytokines or antibodies were added in the culture. For Th1 cell differentiation, the cells were treated with 10 μg/mL α-IL-4 (Peprotech) and 10 ng/mL IL-12 (Peprotech). For Th17 cell differentiation, the cells were treated with 20 ng/mL IL-6, 5 ng/mL TGF-β1 (R&D Systems), 10 μg/mL α-IFN-γ (R&D Systems), and 5 μg/mL α-IL-4 (Peprotech). For Treg cell differentiation, the cells were treated with 10 ng/mL TGF-β1 (R&D Systems), 10 μg/mL α-IL-4 (Peprotech), 10 μg/mL α-IFN-γ (R&D Systems), and 30 U/mL IL-2 (R&D Systems). Various concentrations of PNS (0, 1, 10, and 100 μg/mL) were added to the culture at the beginning of 72-h cultures.
Naive murine CD4+ T cells were isolated from spleens and lymph nodes of C57BL/6 mice and further purified using naive CD4 Isolation Kit using autoMACS Pro (Miltenyi). After isolation, naive CD4+ T cells were differentiated and treated by PNS under Th conditions indicated as above for 72 h.
Intracellular cytokine staining
CD4+ T cells were incubated for 4 h with PMA (50 ng/mL; Sigma), ionomycin (1 μg/mL; Sigma), and GolgiPlug (BD Biosciences), followed by Fixation/Permeabilization and staining for 30 min at 4°C with the antibodies against IFN-γ, IL-17A, and FOXP3 (BD Biosciences), using an Intracellular Cytokine Staining Kit (BD Biosciences). Cells expressing IFN-γ, IL-17A, and FOXP3 were analyzed by flow cytometry.
RNA extraction and microarray analysis
Total cellular RNA was extracted from Th0, Th1, Th17, and Treg cells with TRIzol reagent (Invitrogen) in normal donors. Approximately, 1 μg of total RNA was labeled using MessageAmp™ II-Biotin Enhanced Kit (Ambion) and hybridized to GeneChip® Human Genome U133 Plus 2.0 Array (Affymetrix) in accordance with the manufacturer's protocols. Expression values were determined with GeneChip Operating Software (GCOS) v1.1.1 software. All data analyses were performed using Partek Genomics Suite 6.6. Expression values for each probe were normalized using the Robust Multiarray Average (RMA) method. The fold changes for each probe were calculated by pairwise comparisons between PNS treated and control samples.
Results
Previous reports examined the effects of PNS largely on innate immune cells such as macrophages. 8 Uveitis, a manifestation of BD, is considered to be a T cell–driven disorder based on studies of a murine model of uveitis (Experimental autoimmune uveitis). 15 We therefore investigated whether PNS acted on CD4+ T cells in vitro. Naive CD4+ T cells from 2 healthy donors (HDs) and 2 BD patients were cultured under polarization conditions of Th0, Th1, Th17, and Treg as indicated in the Methods section for 72 h with or without various concentrations of PNS treatment. As shown in Fig. 1, the presence of low dose PNS (1 μg/mL) slightly promoted the proliferation of Th1 cells in both HD and BD individuals (Fig. 1A, D), while treatment of high dose PNS (100 μg/mL) led to the inhibition of proliferation in all Th1, Th17, and Treg cells (Fig. 1A–F). These data suggest that high dose PNS suppress CD4+ T cell proliferation both in BD patients and in HD.
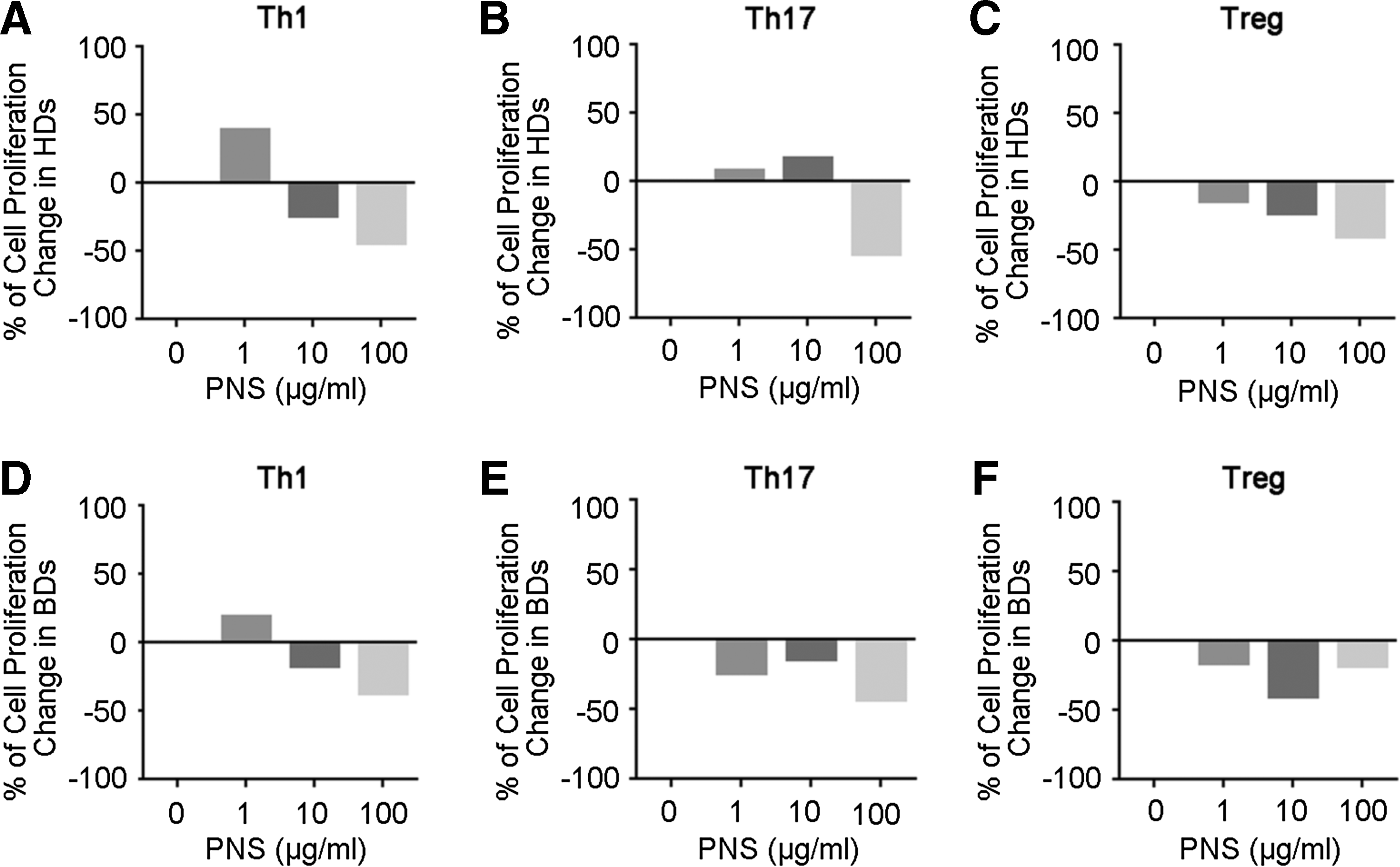
Effect of PNS on the proliferation of helper T cells for HDs and BD patients. Cells were cultured under indicated polarizing conditions and were treated without (0 μg/mL) or with 1, 10, or 100 μg/mL PNS for 72 h. The effects of different concentration of PNS on the proliferation of polarized Th1, Th17, and Treg cells from HDs are shown in
Cytokines produced by polarized CD4+ T cells are crucial for their inflammatory activity. 16 Therefore, we next examined whether PNS affected the expression of Th1/Th17 signature cytokines, IFN-γ and IL-17A, or the signature Treg transcription factor FOXP3 under culture conditions for Th1, Th17, and Treg, respectively. In the in vitro polarized CD4+ T cells from HDs, we found a relatively high expression of FOXP protein and low expression of IFN-γ, compared to its expression in BD patients, while we did not find the baseline difference in expression of IL-17 between HD and BD patients in response to Th17 polarizing stimuli (Fig. 2A). In both HD and BD individuals, we found that the high dose PNS treatment (100 μg/mL) was able to reduce the expression of inflammatory cytokines IFN-γ and IL-17A without significantly downregulating the expression of FOXP3 (Fig. 2A). Interestingly, a lower dose of PNS was required to suppress the expression of IFN-γ and IL-17A in murine Th1 and Th17 cells, respectively (Fig. 2B), suggesting that PNS might function as an anti-inflammatory agent in both human and mouse cells.
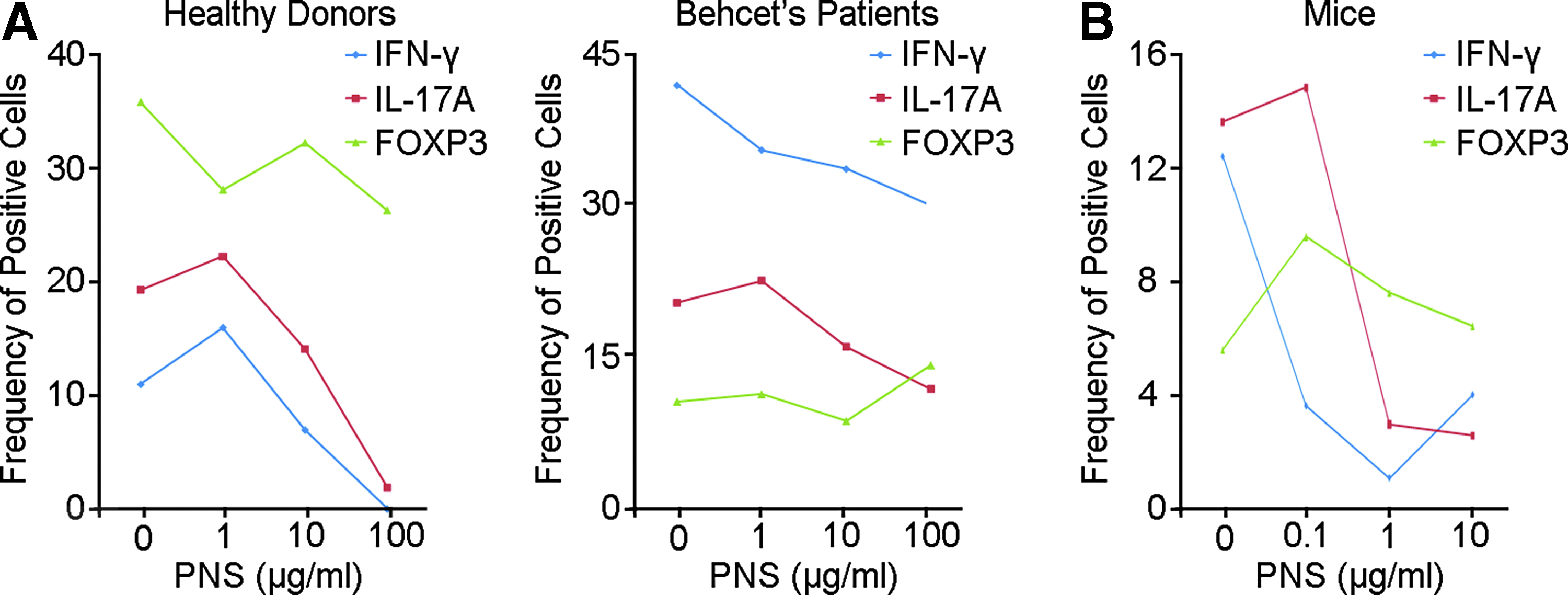
Cytokine expression of Th cells in healthy donors, Behcet's patients
In addition to signature cytokines, Th cells function through expressing many molecules as the effectors of either antipathogenic or inflammatory mediators. 17 We therefore next assessed the genomewide expression profiles of Th cells (polarized from a HD) in response to high dose (100 μg/mL) PNS treatment using Affymetrix microarray analysis. The principal component analysis (PCA) showed that among 4 Th cells, Th17 had the greatest overall gene expression alternation in response to PNS treatment (Fig. 3A). Interestingly, elevation by at least 2-fold in the expression of 12, 5, 99, and 90 genes was found in Th0, Th1, Th17, and Treg cultures in response to PNS treatment, respectively (Fig. 3B). However, the expression of 332 genes was downregulated by PNS (>2-fold decrease) in Th17 cells, while the numbers of genes with decreased expression by at least 2-fold were 22, 13, and 5, in Th0, Th1, and Treg cells, respectively (Fig. 3B). These data demonstrate that PNS have a preferential effect on Th17 cells.
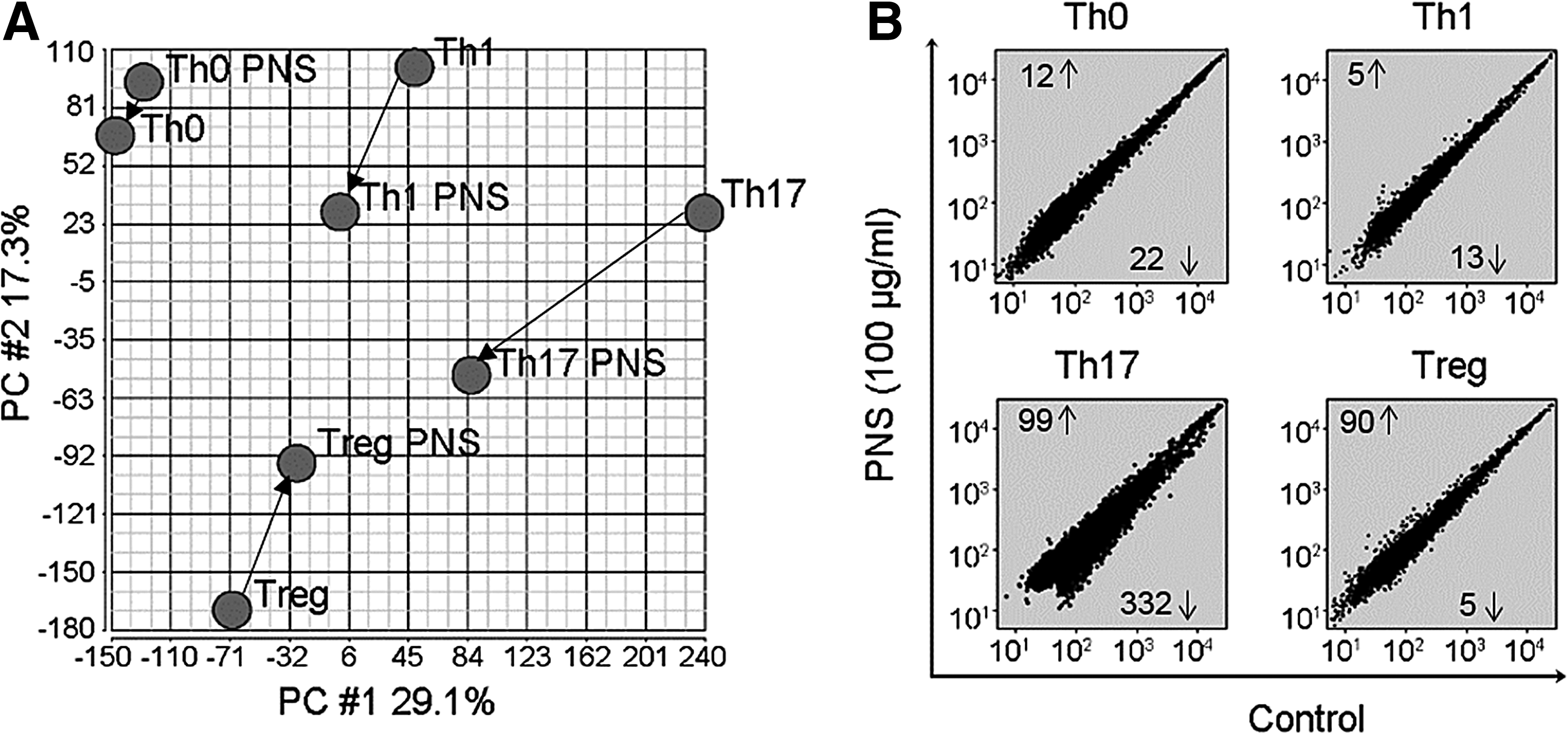
Microarray analysis of global gene expression in response to PNS treatment.
Notably, PNS suppressed the expression of many cytokine genes beyond IL17A. As indicated in Table 1, treatment of PNS reduced the expression of key cytokines and receptors in Th17 cells, including IL26, IL12RB2, IL2RA, IL1R2, IL23R, and IL21R. These data suggest that PNS specifically inhibit the inflammatory function of Th17 cells by controlling the expression of many key cytokines and cytokine receptors. In addition to these cytokines and receptors, we performed Gene Ontology analysis on the 431 genes with at least 2-fold difference in their expression between PNS treated and untreated Th17 cells, using DAVID (6.7) (https://david.ncifcrf.gov/). 18 Our results suggested that PNS significantly modified the genes regulating Cell Cycle and Cell Proliferation in Th17 cells (Table 2), confirming its antiproliferative effects shown in Fig. 1.
Fold change in PNS treated cells compared to untreated cells.
PNS, Panax Notoginseng saponins.
Discussion
Panax Notoginseng and PNS have been used for treating trauma and inflammatory diseases for hundreds of years. Recent experiments are revealing the molecular mechanisms by which PNS control inflammation. Numerous studies have demonstrated that PNS function by directly modulating macrophages through PKC-NFκB pathways.19,20 Our current study indicates that PNS also directly control the proliferation and differentiation of Th17 cells by globally downregulating the expression of inflammatory cytokines and cell cycle genes. However, current study only included 2 inactive BD patients and 2 HDs. Therefore, a comprehensive study with large sample size and active patients is warranted to define PNS's effect on BD pathogenesis.
BD is a chronic disorder characterized by Th17-driven inflammatory responses. Conventional immunosuppressive therapies such as glucocorticoids and CsA, as well as recently developed biologics, can effectively control the clinical symptoms of BD patients; however, many patients develop drug resistance or intolerance21,22 to these treatments highlighting the need for new remedies. As PNS preferentially targets Th17 cells in controlling the immune responses, it may serve as a new therapy for autoinflammatory diseases such as BD.
Footnotes
Acknowledgments
This work was supported by the National Basic Research Program of China (2015CB964601 and 2013CB967001) and the Li Foundation Heritage Prize to L.W.
Authors' Contribution
L.W. designed the study; J.W., X.W., and Z.L. performed experiments; P.W.B. and R.B.N. contributed to data analysis; J.W., X.W., and L.W. wrote the article.
Author Disclosure Statement
No competing financial interests exist.