
Abstract
Abstract
Purpose:
Chronic dryness of the ocular surface evokes sensitization of corneal cold-sensitive neurons through an increase of sodium currents and a decrease of potassium currents, leading to the unpleasant dryness and pain sensations typical of dry eye disease. Here, we explored the effects of amitriptyline, a voltage-gated Na+ channel blocker used for the treatment of depression and chronic pain, on nerve terminal impulse (NTI) activity of cold-sensitive nerve terminals recorded in intact and tear-deficient guinea pig corneas.
Methods:
Main lachrymal gland was surgically removed in anesthetized guinea pigs to induce chronic tear deficiency. Four to 6 weeks afterward, animals were sacrificed and both corneas placed in a perfusion chamber superfused at 34°C. Thermal stimuli were induced by changing the solution temperature from 34°C to 20°C (cooling ramp) and from 34°C to 50°C (heating ramp). Spontaneous and stimulus-evoked NTIs of cold-sensitive nerve terminals were recorded before, during, and after perfusion with solutions containing amitriptyline at different concentrations (3–30 μM).
Results:
Perfusion with amitriptyline inhibited irreversibly and in a concentration-dependent manner the spontaneous NTI activity of cold thermoreceptors of intact corneas. This effect was less evident in tear-deficient corneas. In addition, amitriptyline (10 μM) attenuated the maximal response to cooling ramps without changing cold threshold in intact but not in tear-deficient corneas. Only cold thermoreceptors with low cooling threshold values were sensitive to amitriptyline.
Conclusion:
Amitriptyline effectively reduces the activity of cold thermoreceptors, although its efficacy is different in intact and tear-deficient corneas, which might be due to the changes induced by ocular dryness in the expression of the various voltage-gated Na+ channels responsible of the action potential generation and propagation.
Introduction
D
Peripheral axons of the trigeminal neurons innervating the ocular surface in mice have been classified into 3 functional types: mechanoreceptor (∼15%), polymodal nociceptor (∼70%), and cold thermoreceptor fibers (∼10%–15%). 5 The latter are subdivided into high background activity, low threshold (HB-LT) cold thermoreceptors, and low background activity, high threshold (LB-HT) cold thermoreceptors. 6 HB-LT thermoreceptors display a continuous ongoing discharge at the normal temperature of the cornea (∼35°C), increasing their nerve impulse firing in response to small temperature reductions as those occurring during normal tear evaporation. 7 It has been postulated that the background activity of the HB-LT cold thermoreceptors contributes to maintain the tonic basal tearing and blinking,8–10 whereas the normally silent and less sensitive LB-HT cold thermoreceptors are recruited only by intense cooling produced during pronounced drying and contribute to the discomfort sensations and irritative tearing accompanying DED.10–12
Amitriptyline is a highly lipophilic tricyclic antidepressant drug additionally used to treat chronic and neuropathic pain. 13 This analgesic effect seems to be independent of its antidepressant action. 14 The precise mechanism for pain relief of amitriptyline is not fully understood. However, this drug inhibits various voltage-gated Na+ channels, thus reducing the generation and conduction of action potentials in primary sensory neurons 15 and thereby, the hyperexcitability and aberrant firing often observed in peripheral nociceptor fibers of inflamed/injured tissues mediating chronic pain.
An abnormally augmented activity of corneal cold-sensitive nerve terminals and in a lesser degree of polymodal nociceptor terminals has been reported in guinea pigs, wherein basal tearing was markedly decreased by removal of the exorbital lachrymal gland, thus reproducing experimentally the DED conditions. 12 Such disturbed corneal nerve activity in tear-deficient animals was due to an increase of Na+ currents and a reduction of K+ currents in trigeminal corneal neurons, particularly prominent in cold-sensitive neurons, suggesting that such altered currents are responsible for the abnormal cold thermoreceptor activity underlying sustained discomfort observed in chronic DED patients.12,16,17 In this study, we analyzed whether the Na+ channel-blocker drug amitriptyline affects the impulse activity of corneal cold-sensitive nerve terminals of intact and tear-deficient guinea pigs.
Methods
Dry eye model
Guinea pigs of both sexes (250–350 g, n = 11) were anesthetized with intraperitoneal (i.p.) injection of xylazine hydrochloride (10 mg/kg) and ketamine hydrochloride (90 mg/kg) and the unilateral lachrymal gland was surgically removed as previously described. 12 Guinea pigs were bred for 4–6 weeks postsurgery, receiving water and food ad libitum before corneal nerve activity was recorded. A group of nonoperated age-matched guinea pigs (n = 20) was housed in the same conditions and used as control (intact corneas). The study was carried out in accordance with the Association for Research in Vision and Ophthalmology statement for the Use of Animals in Ophthalmic and Vision Research, and the European Union Directive and the Spanish Regulations on the protection of Animals Used for Research, following a protocol approved and supervised by the Ethics Committee of the University Miguel Hernández.
Electrical recording of nerve terminals in isolated corneas
Recordings were conducted as described previously.18–20 Guinea pigs were killed with an overdose of sodium pentobarbitone (100 mg/kg, i.p.). Both corneas were then isolated by a circular cut around the limbus and incubated for 1 h in saline (138.6 mM NaCl, 3.35 mM KCl, 21 mM NaHCO3, 9.9 mM glucose, 0.6 mM NaH2PO4, 2.5 mM CaCl2, and 1 mM MgCl2) gassed with a mixture of 95% O2 and 5% CO2. Afterward, the excised cornea was pinned to the bottom of a silicone-coated chamber filled with saline and maintained at 34°C (basal temperature) with a feedback-controlled Peltier device. Nerve terminal impulse (NTI) activity was recorded using a glass micropipette applied to the surface of the cornea with a micromanipulator and fixed in place by slight suction. Pipettes had a tip diameter of around 50 μm and were filled with the perfusion physiological saline. Electrical signals were recorded with respect to an Ag/AgCl pellet in the bath. Electrical activity was amplified (AC preamplifier NL104; Digitimer, Welwyn, UK), filtered (low frequency cutoff 150 Hz, high frequency cutoff 5 kHz; bandpass filter module NL125; Digitimer), and transferred to a PC at 20 kHz with a CED micro1401 acquisition system (Cambridge Electronic Design Limited, Cambridge, UK), for off-line analysis (Spike2 also from Cambridge Electronic Design). The corneal surface was explored using the tip of the micropipette until a site in which spontaneous activity of a single nerve terminal was detected. NTIs originating from a single cold-sensitive nerve ending were identified usually by their similar amplitude and their rhythmic, regular ongoing discharge that increased upon cooling and decreased upon warming. For thermal stimulation, the temperature of the physiological solution bathing the cornea was changed using the Peltier cell from the basal temperature of 34°C down to 20°C (at approximately −0.5°C.s−1; cooling ramp) or up to 50°C (at approximately +0.4°C.s−1; heating ramp). Spontaneous NTI activity at basal temperature was recorded for at least 1 min before application of a cooling ramp to 20°C, followed by rewarming to the basal temperature. After recovering a stable baseline activity, the solution bathing the cornea was heated to about 50°C and immediately cooled again to basal temperature. The protocol of thermal stimulation was applied 10 min before and 2 min after starting drug perfusion.
Different parameters were calculated to quantify the response to thermal stimuli (Fig. 1): (1) ongoing activity at basal temperature of 34°C (expressed as the mean frequency, in impulses per second, at the basal temperature); (2) cooling threshold during the cooling ramp from 34°C to 20°C (expressed as the decrement in temperature required to increase by 25% the frequency of discharge at basal temperature); (3) peak frequency during the cooling ramp from 34°C to 20°C (maximal firing frequency in Hz reached during the cooling ramp); (4) temperature change to reach the peak frequency during the cooling ramp from 34°C to 20°C (expressed as the temperature change in °C required to reach maximal firing frequency per second during the cooling ramp); (5) warming threshold when rewarming from 20°C to 34°C (expressed as the increment in temperature at which the terminal started firing again); (6) cooling threshold when returning from 50°C to basal temperature of 34°C (expressed as the temperature decrement required to recover NTI firing after being silenced by heating); (7) peak frequency when cooling from 50°C to 34°C (maximal firing frequency in Hz, reached during cooling from 50°C to 34°C); (8) temperature change required to reach peak frequency (expressed as the temperature change to reach the peak frequency when cooling from 50°C to 34°C); (9) silencing temperature during the heating ramp from 34°C to 50°C (temperature value at which the terminal stopped firing during the heating ramp); (10) heating response threshold for the paradoxical response to heat (when present) (expressed as the value of temperature in °C required to evoke firing during the heating ramp); and (11) paradoxical response to heat (expressed as the total number of impulses during the heating ramp).

Example of NTI activity of corneal cold thermoreceptors in response to thermal stimulation. Upper panel: temperature changes. Middle panel: impulse firing direct record. Lower panel: frequency of NTI activity. The onset and termination of experimental maneuvers are indicated by vertical dotted lines. See Methods section for explanation of the different parameters measured. NTI, nerve terminal impulse.
Solutions
Amitriptyline (Sigma-Aldrich, St. Louis, MO) was prepared as a 100 mM stock solution in water and diluted to the desired final concentration (3, 10, or 30 μM) with the perfusion saline.
Experimental protocol
The presence of an active cold nerve terminal was ascertained by the appearance of spontaneous NTI activity after application of the micropipette to the corneal surface. After a recording period of at least 2 min at basal temperature, thermal stimuli were applied. First, cold stimulation was performed by decreasing the bath temperature from 34°C down to 20°C (cooling ramp), followed by rewarming to return to the basal temperature. After a time period of 120–150 s at 34°C, a heating ramp from 34°C to 50°C was performed, returning to 34°C afterward.
After a resting period of 15 min at 34°C, perfusion with control saline was switched to saline containing amitriptyline at one of the tested concentrations (3, 10, or 30 μM). Two minutes after the onset of the perfusion with amitriptyline, a cooling ramp down to 20°C and a heating ramp to 50°C were applied, as previously described. Ten minutes after starting perfusion with amitriptyline, the cornea was perfused again with the control saline for a period of at least 45 min.
Statistics
Data were collected and processed for statistical analysis using SigmaStat software (Systat Software, Inc., San Jose, CA). Data are expressed as mean ± SEM. The differences in each NTI activity parameter between intact and tear-deficient corneas were compared with independent Student's t-test or Mann–Whitney U-test, as needed. The differences in NTI activity parameters measured before and after amitriptyline perfusion were compared with paired t-test or Wilcoxon signed rank test, as needed. P < 0.05 was considered statistically significant.
Results
Differences in NTI firing between corneal cold-sensitive units of intact and tear-deficient eyes
As shown in Table 1, mean background NTI activity at 34°C and peak firing frequency values reached during a cooling ramp performed at different initial temperatures (from 34°C to 20°C and from 50°C to 34°C) were slightly increased in tear-deficient corneas when compared with those in intact corneas, although differences did not reach the significance level. However, chronic tear deficiency caused a significant reduction of the cooling threshold and also of the temperature fall required to reach the peak frequency value during a cooling ramp from 50°C to 34°C, whereas these parameters measured during a cooling ramp from 34°C to 20°C were less affected. Corneal dryness did not modify significantly the parameters defining the response to heat of cold nerve terminals (Table 1), although this negative result should be taken cautiously due to the low number of recorded terminals.
Data represented as mean ± SEM.
P < 0.05, **P < 0.01, significant difference between intact and tear-deficient corneas.
In Fig. 2, we represent the individual cold nerve terminals of intact and tear-deficient corneas according to their NTI firing characteristics. The shaded dark blue area shown in Fig. 2A segregates a subpopulation of terminals exhibiting low background activity and a weaker peak response to the cooling ramp and thus considered LB-HT cold thermoreceptor terminals, whereas those included within the light blue area were considered HB-LT cold thermoreceptor terminals. 6 As shown in Fig. 2B, the group of LB-HT cold thermoreceptor terminals was absent in tear-deficient corneas.
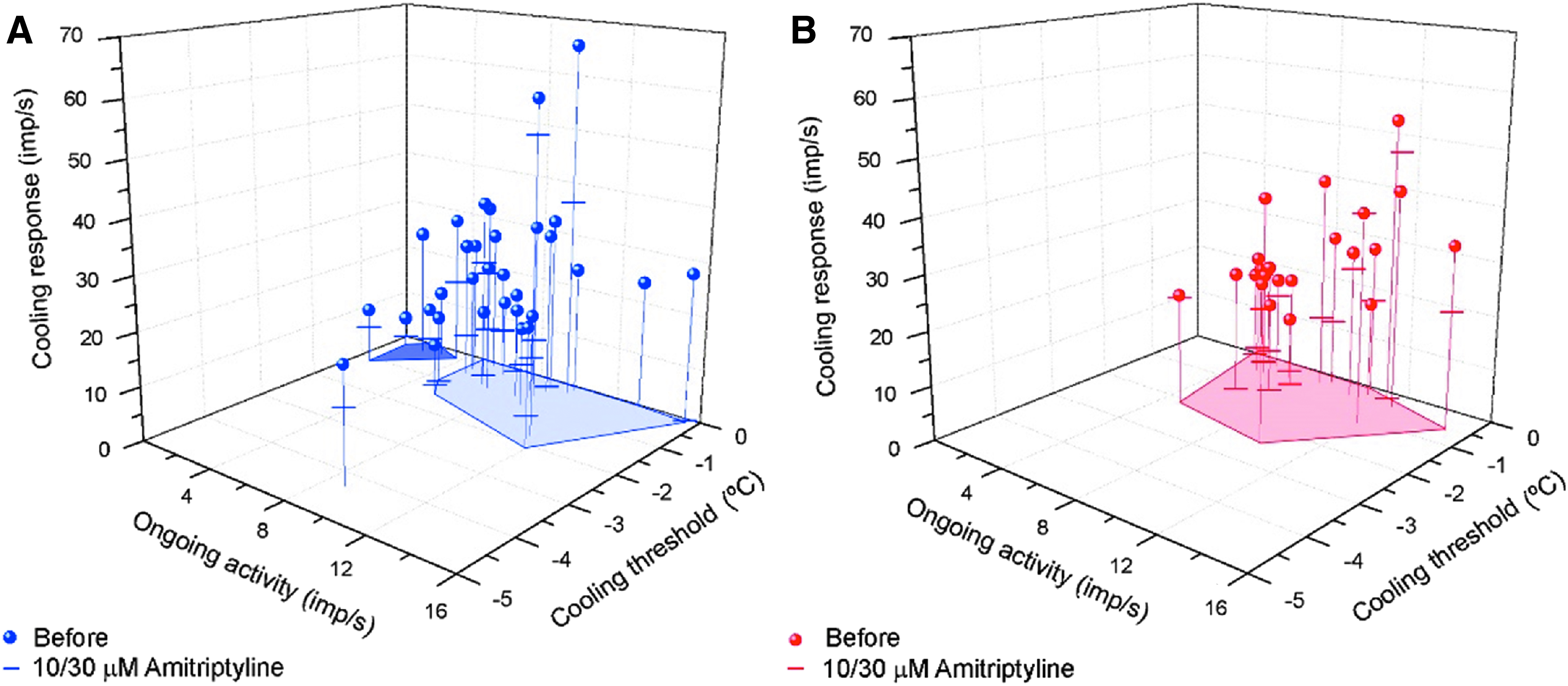
Effects of amitriptyline on NTI activity of corneal cold-sensitive terminals from intact
Effect of amitriptyline on the spontaneous and stimulus-evoked NTI activity of cold-sensitive units from intact and tear-deficient corneas
NTI activity was measured before and during perfusion of the recording chamber with solution containing amitriptyline for 10 min at 34°C.
Ongoing NTI activity at 34°C was gradually and irreversibly decreased during exposure to amitriptyline in a dose-dependent manner both in intact (reductions of 34%, 56%, and 68% of the pretreatment value for 3, 10, and 30 μM amitriptyline, respectively. P < 0.001, ANOVA on ranks) and in tear-deficient corneas (reductions of 34% and 85% of pretreatment values for 10 and 30 μM amitriptyline, respectively. P < 0.001, ANOVA on ranks) (Fig. 3A, B). As shown in Fig. 3C, the mean inhibitory effect of 10 μM amitriptyline on the ongoing activity was weaker in tear-deficient corneas, so that differences in ongoing activity between intact and tear-deficient corneas were significant for the values measured 2 and 10 min after the onset of the perfusion with the drug (Fig. 3C).

Effects of amitriptyline on ongoing NTI activity in corneal cold-sensitive terminals.
Amitriptyline also decreased significantly the NTI activity peak evoked by a cooling ramp in cold thermoreceptors of intact but not of tear-deficient corneas, as shown in Table 2. In Fig. 2A and B, the value of the peak response to cooling ramps after exposure to amitriptyline in each one of the individual terminals is indicated by a horizontal bar, evidencing that the effect of amitriptyline varied widely among individual cold-sensitive terminals and was, altogether, more pronounced in the terminals of intact corneas. Likewise, amitriptyline at 10 μM significantly decreased the peak frequency induced by cooling from 34°C to 20°C and from 50°C to 34°C in intact corneas, whereas in tear-deficient corneas the change in these parameters was small and did not reach significance levels (Table 2). Moreover, mean values of cooling threshold and of the temperature change required to reach peak frequency were affected by amitriptyline in the terminals of the intact but not of the tear-deficient corneas. Nonetheless, in both groups, amitriptyline decreased significantly the temperature change required to silence background NTI activity by application of a heating ramp from 34°C to 50°C (Table 2). These differences appear to be due, at least, in part, to absence of sensitivity to amitriptyline in a larger number cold thermoreceptor terminals. To test this possibility, we segregated cold thermoreceptor terminals into a group of amitriptyline-sensitive units, wherein the drug caused a reduction of mean background activity of >15% of the control value, and another of amitriptyline-insensitive units, where the inhibitory effect of the drug was <15%. Three out of 9 units in the intact corneas and 5 out of 12 units in tear-deficient corneas were considered, according to this criterion, insensitive to amitriptyline (Fig. 2A, B). Notably, amitriptyline-insensitive terminals of both intact and tear-deficient corneas exhibited comparatively higher cooling thresholds and required a larger temperature decrease to reach peak frequency during a cooling ramp (Tables 3 and 4). When we compared the effects of amitriptyline on peak firing frequency evoked by cold in amitriptyline-sensitive units of intact and tear-deficient corneas, we noticed in both cases a similar and significant reduction in peak frequency values in comparison with pretreatment values (Fig. 2A, B; Tables 3 and 4).
Data represented as mean ± SEM.
P < 0.05, **P < 0.01, significant difference between intact and tear-deficient corneas.
Data represented as mean ± SEM.
P < 0.05, **P < 0.01, significant difference between amitriptyline-sensitive and amitriptyline-insensitive units.
NR, no response.
Data represented as mean ± SEM.
P < 0.05, **P < 0.01, significant difference between amitriptyline-sensitive and amitriptyline-insensitive units.
Discussion
This study reveals that amitriptyline inhibits irreversibly and in a concentration-dependent mode, the spontaneous activity at 34°C of cold thermosensitive nerve terminals of the cornea, and decreases their peak frequency response to cooling ramps but not their cooling threshold. These effects were more pronounced in terminals showing a high sensitivity to temperature reductions (HB-LT cold thermoreceptors). In tear-deficient corneas, where sensitivity to cold of thermoreceptor terminals is altered, we observed a generally weaker inhibition by amitriptyline of the spontaneous and stimulus-evoked activity, due to an insensitivity to the drug of LB-HT cold thermoreceptor terminals.
Amitriptyline acts as a potent use-dependent blocker of voltage-gated sodium channels.21–24 Nav channels are critical to establish the resting potential of neurons and to setting the threshold for the generation of action potentials. 25 Most of the sodium channel isoforms are sensitive to tetrodotoxin (TTXs), whereas Nav1.8 and Nav1.9, which are expressed almost exclusively by peripheral sensory neurons,26,27 are resistant to TTX up to millimolar concentrations. The importance of TTXr NaV channels for the generation of propagated nerve impulse activity at peripheral sensory terminals of unmyelinated afferent nerve fibers could be proved for the first time in the cornea.20,28–30 Recent work 31 on skin sensory nerves has additionally demonstrated that Nav1.8 is the most prominent Nav channel isoform sustaining nerve impulse conduction at the terminal portion of unmyelinated sensory axons.
Amitriptyline is known to block voltage-gated Na+ channels by binding to specific amino acid residues in the α-subunit S6 segments, which are located within the ion-conducting pore of the channel.15,32,33 Amitriptyline causes an use-dependent blockade of both TTXs and TTXr Nav1.8 currents that is state dependent, with a preference for open and inactivated states rather than for resting channel states. 33 Expression and normal function of both families of Nav channels are essential for the generation of impulse activity in cold thermoreceptor corneal nerve terminals.20,28–30 Sodium channel inhibition by amitriptyline23,24 explains the attenuating effects of the drug on the basal ongoing discharge and nerve impulse firing in response to cooling seen in corneal cold thermoreceptors. As could be expected, other voltage-gated sodium channel blockers such as lidocaine, lacosamide, or hainantoxin-IV produce similar firing frequency reductions in corneal cold thermoreceptors. 12 Amitriptyline seems to also block Kv channels, 34 a mechanism that may additionally contribute to the impulse firing changes evoked by the drug in cold terminals. Dynamic responses of corneal cold thermoreceptors to heating seem to depend on potassium channels. 35 Thus an effect of amitriptyline on these channels could also be partly responsible for the silencing by heating ramp of the ongoing activity of corneal cold thermoreceptors at 34°C, which occurs at lower temperatures in the presence of amitriptyline.
Collectively, our observations confirm that chronic corneal desiccation resulting from surgical removal of the main lacrimal gland evokes an enhanced excitability of cold thermoreceptors,12,36,37 a change attributable to damage of corneal sensory nerve terminals. The spontaneous, aberrant impulse firing developed by injured sensory neurons is caused by alterations in the expression of various Na+ channel isoforms, including TTXs Nav1.3 in the soma and injury-evoked redistribution of Nav1.8 and Nav1.9 between cell body and peripheral terminals.38–40 The characteristics and extent of these disturbances are variable, depending on the nature, location, and extension of the injury and on the molecular and functional signature of the affected neurons. In intact corneas, the expected differences in the expression of voltage-dependent sodium channels between HB-LT and LB-HT cold thermoreceptors can explain their variable sensitivity to amitriptyline, generally higher in HB-LT cold thermoreceptors. Under sustained corneal dryness, nerve terminal injury additionally stimulates upregulation and downregulation and impaired distribution of Na+ channel isoforms. Such changes further alter the excitability of cold thermoreceptor terminals, also modifying their sensitivity to amitriptyline according to the type of cold thermoreceptor terminal affected by injury. We cannot exclude the possibility that other mechanisms such as changes in activity/expression of K+ channels activated by voltage and cyclic nucleotide (i.e., HCN) contribute to the altered excitability of cold thermoreceptors in chronic dryness and the differential effect of amitriptyline in naive and tear-deficient eyes.
In the present experiments, no low background cold thermoreceptor terminals were recorded in tear-deficient corneas, although cooling threshold was not significantly modified by dryness. This may be due to changes in the expression and/or activity of ion channels produced by nerve injury consecutive to chronic dryness or developed with aging, which result in a higher probability of recording HB-LT nerve endings and also in the appearance of nerve endings exhibiting a mixture of the electrophysiological properties of HB-LT and LB-HT cold nerve terminals, that is, nerve terminals with high background activity and higher threshold for activation during cooling (HB-HT), and nerve terminals with low background activity and weak response to cooling ramps (LB-LT cold terminals).12,41
Amitriptyline is a drug originally used to treat depression. 42 Depression has been often associated with dry eye syndrome and this pathology is frequently found in patients taking antidepressants and medication for other psychiatric conditions for long periods of time,43,44 although no causal link has been established. The marked inhibitory effect of amitriptyline on corneal cold thermoreceptor terminal background activity, whose role in maintaining basal tear secretion is important, has to be taken into consideration when prescribing this drug to depressive patients suffering from DED. Nonetheless, the possibility of reducing the abnormal activity in dryness-injured corneal nerve terminals underlying unpleasant dry eye sensations with topical application of amitriptyline possibly deserves further scrutiny.
Footnotes
Acknowledgments
This work was supported by grants SAF2014-54518-C3-1-R and SAF2014-54518-C3-2-R (from Ministerio de Economía y Competitividad, Spain, and ERDF, European Union), and 25870852, 26460348, 15K08250, and 16K19023 (from JSPS KAKENHI, Japan). The Instituto de Neurociencias de Alicante is a Severo Ochoa Excellence Center. Authors thank technical assistance of Carolina Luna.
Author Disclosure Statement
No competing financial interests exist.