
Abstract
Abstract
Purpose:
To study the pharmacological profile of the serotonin (5-hydroxytryptamine [5-HT]) receptor subtype mediating contractions in bovine isolated ciliary muscles.
Methods:
Ciliary muscle strips were isolated from bovine eyeballs and mounted in organ baths containing aerated (95% O2, 5% CO2) Krebs buffer solution maintained at 37°C. Each muscle strip was attached at 1 end to a Grass Force-displacement Transducer connected to a Polyview Computer System for recording changes in isometric tension. After an equilibration period, ciliary muscle strips were exposed to selective agonists and antagonists of 5-HT receptors.
Results:
Both selective and nonselective agonists for 5-HT produced concentration-dependent contractions of isolated ciliary muscles with the following rank order of potency: BW723C86>α-methyl-5-HT>MK-212>>8-hydroxy-DPAT>quipazine>R-DOI>>5-HT>>tryptamine. The selective 5-HT2 receptor antagonists, M-100907 (5-HT2A), RS-127445 (5-HT2B), and RS-102221 (5-HT2C), produced noncompetitive inhibition of the contractile effects of selective agonists yielding antagonist potency (pKB) values of 251 ± 27.2 nM (n = 4), 52.5 ± 6.3 nM (n = 4), and 79.4 ± 9.5 nM (n = 4), respectively.
Conclusion:
On the basis of the profile of activity of selective agonists and antagonists, we conclude that the 5-HT2B and 5-HT2C receptor subtypes appear to be the predominant serotonin receptors that mediate the contractile action of this amine in bovine isolated ciliary muscles.
Introduction
S
Methods
Tissue preparation
Bovine eyeballs were obtained from a slaughterhouse in Dallas, Texas, and transported to the laboratory on ice within 24 h of enucleation. The methodology for dissection of the ciliary muscle was as described by Lograno and Romano, with slight modification. 9 Briefly, eyeballs were dissected, the vitreous humor and lens removed, and ciliary muscles separated from the scleral spur and choroid. Muscle strips were placed in aerated (95% O2, 5% CO2) Krebs buffer solution at 4°C until time of assay.
Measurement of isometric tension
Isolated bovine ciliary muscle strips (4–5 mm length) were mounted in 25 mL organ baths containing aerated Krebs solution (pH 7.4) at 37°C. The Krebs buffer solution had the following composition (mM): sodium chloride, 118; potassium chloride, 4.8; calcium chloride, 2.5; potassium phosphate monobasic, 1.2; sodium bicarbonate, 25; magnesium sulfate, 2.0; and dextrose, 10; with flurbiprofen (0.003 mM) added to inhibit the synthesis of prostaglandins through cyclooxygenase. Each muscle strip was attached to a Grass FT03 transducer under a resting tension of 300 mg using silk thread and allowed to equilibrate for 30 min. Changes in isometric tension were measured using Grass Polyview Computer Software. Tissues were initially challenged with carbachol (10 μM) to assess the functional state of the muscle strips. Tissues were then washed with physiological solution and the tension readjusted before addition of drugs for the actual study.
The contractile actions of selective 5-HT1 (8-OH-DPAT), 5-HT2 (R-DOI, BW732C86, α-Me-5HT, MK-212), and 5-HT3 (quipazine) agonists were observed under basal conditions of tone to determine relative potencies. Serotonin and tryptamine were used as standard nonselective agonists of 5-HT receptors. The bovine ciliary muscle strips were exposed to the selective 5-HT receptor antagonists for 45 min before reexamining the effects of the agonists.
Data analysis
The response of each tissue is expressed as percentage of the maximum response to each compound. Concentration-response curves were analyzed, and EC50 values (agonist concentration that produced 50% of the observed maximum response) were determined using GraphPad Prism 4.0 Software (San Diego, CA). Antagonist potencies were calculated as pKB values for noncompetitive antagonism. pKB values were calculated from the equation: pKB = log (dose ratio −1) − log (antagonist concentration). Values are expressed as arithmetic mean ± standard error of the mean. Significance of differences between control and test values was evaluated using one-way analysis of variance (GraphPad Prism software, San Diego, CA). P values <0.05 were accepted as statistically significant.
Chemicals
Standard buffers, common high-grade laboratory chemicals, and 5-HT agonists (BW732C86, α-Me-5HT, MK-212, R-DOI, 8-OH-DPAT, Quipazine) and antagonists (M-100907, RS-127445, and RS-102221) were obtained from Tocris Biochemicals (Minneapolis, MN), Sigma-Aldrich (St. Louis, MO), and Cayman Chemical (Ann Arbor, Michigan).
Results
Effects of 5-HT receptor agonists
All 5-HT receptor agonists examined caused concentration-dependent contractions of the isolated bovine ciliary muscles. Figure 1 illustrates the contractile concentration-response curves to BW723C86 and MK-212, selective agonists at 5-HT2B and 5-HT2C receptors, respectively. The 5-HT2-receptor-selective agonists that displayed the lowest EC50 values (0.5–4.1 nM) were BW723C86 (5-HT2B-selective), α-Me-5HT (5-HT2B/2C-selective), and MK-212 (5-HT2C-selective), while R-DOI (5-HT2A-selective) had one of the highest EC50 values (190 nM) (Table 1). The 5-HT1A receptor agonist, 8-hydroxy-DPAT, and 5-HT3 receptor agonist, quipazine, also elicited contractions of the bovine ciliary muscle yielding EC50 values of 130 and 170 nM, respectively (Table 1). Both 5-HT and tryptamine produced contractions of the ciliary muscle with a lower potency compared to agonists with selectivity for 5-HT1–3 receptors and their subtypes (Table 1). A comparison data from studies showing potencies of selective and nonselective serotonergic agonists in the rat isolated stomach fundus (contractile action), 23 human ciliary muscle (phosphoinositide turnover), 11 and human uterine cell line (phosphoinositide turnover) are also shown in Table 1. 24 The 5-HT2B antagonist RS-127445 displayed the lowest relative potency (52.5 nM) of serotonergic antagonists as shown in Table 2.
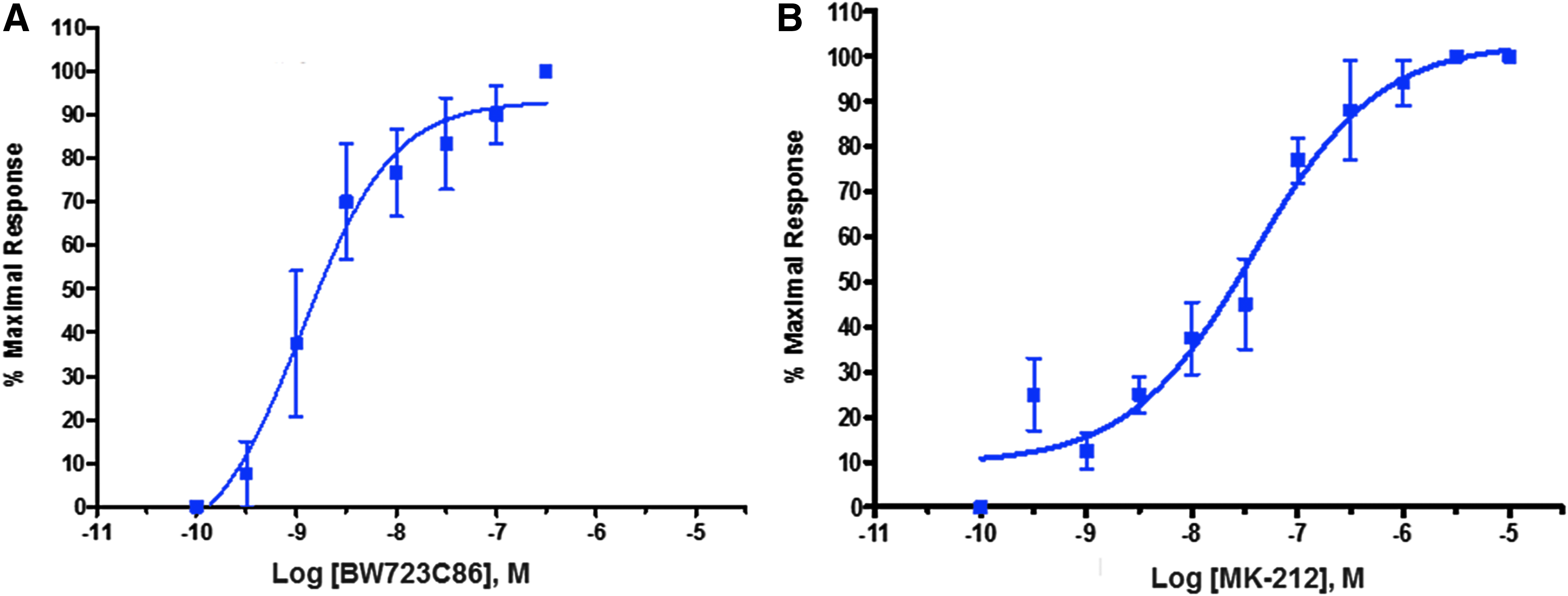
Contractile concentration-response curves to serotonergic agonists, BW723C86
Comparison of selective and nonselective serotonergic agonist potency data has been included for: human ciliary muscle PI turnover 11 ; rat stomach fundus contraction 23 ; and human uterine cell line PI turnover. 24 Data shown represent mean ± standard error of mean; the numbers of observation are shown in parenthesis.
PI, phosphoinositide.
Data shown represent mean ± standard error of mean; the numbers of observation are shown in parenthesis.
Effect of selective 5-HT2 receptor antagonists
The effects of different concentrations of antagonists selective for 5-HT2A (M-100907), 5-HT2B (RS-127445), and 5-HT2C (RS-102221) receptors (0.1–30 μM) were used to block concentration-response curves to selective agonists, R-DOI, BW7238C86, and MK-212, respectively. Figure 2 shows the antagonism of contractile response to BW723C86 by different concentrations of RS-127445. Analysis of the data using Schild Plots was not possible due to the noncompetitive nature of this antagonism. Consequently, dose ratios were calculated for each concentration of the antagonist, and pKB values were then estimated from the equation: pKB = log (dose ratio −1) − log (antagonist concentration). The calculated pKB value for RS-127445 antagonism of contractions elicited by BW723C86 was 52.5 ± 6.3 nM (n = 4). Both M-100907 and RS-102221 also produced noncompetitive antagonism of contractions elicited by R-DOI and MK-212 yielding pKB values of 251 ± 27.2 nM (n = 4) and 79.4 ± 9.5 nM (n = 4), respectively.

Antagonism of contractile concentration-response curves to BW723C86 by different concentrations of RS-127445 in isolated bovine ciliary muscle: control (blue line) and in the presence of RS-127445 [100 nM (red line); 300 nM (green line); 1 μM (pink line); and 30 μM (orange line)]. Values shown represent mean ± standard error of mean. Number of observations is 4–8.
Discussion
Evidence from the literature has shown that 5-HT acts as a signaling molecule with several important roles in physiological processes. 1 In the eye, 5-HT-containing nerves have been found in the anterior uveal and retinal tissues of several mammalian species.25–27 The presence of 5-HT in the aqueous humor has been reported in humans and several mammalian species.2,28–30 Furthermore, functional 5-HT2 receptors have been identified in ocular tissues critical to IOP regulation such as the ciliary muscle and trabecular meshwork.11,15 Cellular, molecular biological, and morphological techniques have been used to identify and characterize the pharmacology of 5-HT receptor subtypes in ocular tissues.5,11,12,31,32 In addition, the pharmacological actions of 5-HT agonists in ocular tissues of nonhuman primates suggest a potential therapeutic role for 5-HT in the eye,13–15,22 since administration of 5-HT2 receptor agonists has been shown to significantly reduce IOP. Although there is evidence for the presence of 5-HT2 receptors in the bovine ciliary muscle, 9 the subtype/s of 5-HT2 receptor that mediate contractile responses to serotonin are unknown. In the present study, we characterized the pharmacological profile of 5-HT2 receptors using selective agonists. All agonists examined produced concentration-related contractile responses of isolated bovine ciliary muscle with the following rank order of potency: BW723C86>α-methyl-5-HT>MK-212>>8-hydroxy-DPAT>quipazine>R-DOI>>5-HT>>tryptamine. The selective 5-HT2B and 5-HT2C agonists, BW723C86 (5-HT2B), α-Me-5HT (5-HT2B/2C), and MK-212 (5-HT2C), had highest potencies in inducing contractions in bovine ciliary muscle strips. These data support the view that stimulation of 5-HT2B and 5-HT2C receptors produced more significant contraction of bovine ciliary muscles from resting tone than did stimulation of other 5-HT receptor subtypes (5-HT1, 5-HT2A, and 5-HT3). In contrast, 5-HT and tryptamine had very low potencies, indicating their nonselective characteristics as 5-HT receptor agonists. Similarly, both BW723C86 and α-methyl-5-HT demonstrated relatively high potencies in eliciting contractions of the rat stomach fundus and stimulating phosphoinositide turnover in human uterine cell lines indicating that 5-HT2B receptors mediate the observed responses to these compounds.23,24 The high potency exhibited by MK-212 in the bovine ciliary muscle affirms the presence of 5-HT2C receptors in this tissue.
To further elucidate the pharmacological profile of the 5-HT2 receptor that mediates contraction of the isolated bovine ciliary muscle, we investigated the antagonistic action of selective 5-HT2A, 5-HT2B, and 5-HT2C receptor blockers on responses caused by their counterpart selective agonists. The antagonist/agonist pairs: M-100907/R-DOI; RS-127445/BW723C86; and RS-102221/MK-212 produced noncompetitive antagonism yielding pKB values that yielded the following rank order of antagonist potency: RS-127445>RS-102221>>M-100907. The higher potencies exhibited by RS-127445 and RS-102221 indicate that a predominant population of 5-HT2B and 5-HT2C receptors may mediate the contractile response of the isolated bovine ciliary muscle to serotonin. It is feasible that while 5-HT2A receptors may also play a role in these contractions, their overall contribution may be limited based on the low potency exhibited by both M-100907 (5-HT2A antagonist) and R-DOI (5-HT2A agonist) in the bovine ciliary muscle. It is pertinent to note that signal transduction and receptor binding studies with selective 5-HT2 agonists and antagonists in isolated human ciliary muscle cells revealed the 5-HT2A receptor subtype as having the highest functional activity in these cells. In summary, it appears that species differences may exist in the contribution of 5-HT2 receptor subtypes in mediating contraction of mammalian ciliary muscles.
Taken together, our results provide strong pharmacological evidence that stimulation of 5-HT2B and 5-HT2C receptors induces contractions in bovine ciliary muscles and these contractions can be inhibited by antagonists of the respective 5-HT2 receptor subtypes. The ability of agonists at 5-HT receptor sites to regulate aqueous humor dynamics and IOP in experimental animals is well documented (as reviewed by Sharif 13 ). Indeed, the importance of 5-HT receptors in the pathway and processes involved in the regulation of IOP is underscored by the fact that cabergoline, a dopaminergic agonist with serotonergic activity, reduced IOP in ocular normotensive rats and in ocular hypertensive cynomolgus monkeys presumably using 5-HT2 receptors. 17 The implications and potential clinical significance of the findings from the present study cannot be fully explained without additional studies to confirm the physiological role of serotonin and related agents in the bovine ciliary muscle. A major relevance of data obtained in the present study is the pharmacological characterization of the subtype of 5-HT2 receptors in the bovine ciliary muscle that may contribute to the IOP-lowering action of serotonergics in this specie. It is feasible that the ability of 5-HT2B/2C receptors to mediate contraction of the ciliary muscle could account, at least in part, to the ocular hypotensive action of serotonin. The ciliary muscle is integrally involved in modulating intraocular pressure by facilitating aqueous humor outflow. 33 Contraction of ciliary muscles may reduce outflow resistance through the trabecular meshwork.34,35 Agents such as the muscarinic agonist pilocarpine, which induces contraction of the ciliary muscle, can facilitate aqueous humor drainage, thereby decreasing intraocular pressure.36–38 The involvement of 5-HT2 receptors present on ciliary muscles in the regulation of IOP appears to be an important component of the pharmacological action of serotonin on aqueous humor dynamics. Indeed, these receptors have been hypothesized to play a role in regulation of intraocular pressure in humans.13–15,22
We conclude that based on the pharmacological profile of activity of selective agonists and antagonists, the contractile response to serotonin in isolated bovine ciliary muscles are mediated primarily by 5-HT2B and 5-HT2C receptor subtypes. The observed presence of these excitatory serotonin receptor subtypes in bovine ciliary muscle supports their role in the regulation of aqueous humor dynamics and IOP.
Footnotes
Author Disclosure Statement
No competing financial interests exist.