
Abstract
Abstract
Purpose:
To investigate retinal gene expression of tetramethylpyrazine (TMP) eye drop-treated endotoxin-induced uveitis (EIU) in mice and to explore the mechanisms.
Methods:
The inflammatory signs of the anterior segment were evaluated, and clinical scores were graded. The retinal transcriptome from the TMP eye drop-treated and the untreated mice was identified by RNA sequencing (RNA-seq) strategy. Differentially expressed genes (DEGs) were validated by real-time PCR. The protein–protein interaction was analyzed using the STRING software.
Results:
Compared with the TMP-treated group, the inflammatory responses of the untreated control group were much severe and clinical score was remarkably higher (P < 0.001) at 24 h after lipopolysaccharide administration. RNA-seq assay identified 407 DEGs, among which 356 were upregulated and 51 were downregulated. There were 12 upregulated gene ontology terms enriched and 27 upregulated pathways. Seven DEGs, including inflammation-related, complement system-related, and interferon-related genes, were validated using quantitative PCR.
Conclusions:
TMP exerted anti-inflammatory effect in EIU. Local application of TMP inhibited retinal inflammatory response by regulating the inflammation-related genes, suggesting that TMP may be a potential novel therapeutic drug for ocular inflammation.
Introduction
Uveitis is an inflammatory ocular disease that mainly affects the iris, ciliary body, and choroid. 1 It is sight threatening and even causes vision loss within the working population worldwide. 2 The infections and systemic autoimmune disorders may play important roles in the pathogenesis of uveitis, but the specific mechanism is still unclear. An endotoxin-induced uveitis (EIU) mouse model with the injection of lipopolysaccharide (LPS) systemically or locally serves as a classic animal model to study human acute infectious uveitis, in which the inflammation involves both anterior and posterior segments of the eyes. 3 Even though the conventional treatment for uveitis, including corticosteroids, immunosuppressive agents, and nonsteroidal anti-inflammatory drugs, is efficacious to many patients, there are still limitations. Some patients do not respond well to these drugs. Furthermore, long-term application of these drugs may cause numerous undesirable side effects.4,5 Therefore, it is highly desirable to search novel and effective therapeutic drugs with less side effects.
Tetramethylpyrazine (TMP), also known as ligustrazine, is the main active ingredient extracted from Ligusticum wallichii (Chuanxiong), which is used in traditional Chinese medicine. 6 It has been extensively applied in the treatment of cardiovascular and cerebrovascular diseases for its antioxidative and vascular protection functions.7,8 It has also been discovered to exert protective effects on multiple organs and systems.9,10 TMP attenuated LPS-induced pulmonary vascular permeability and decreased the release of inflammatory cytokines in Sprague-Dawley rats. 11 In addition, TMP inhibited the production of reactive oxygen species in microglia cells challenged by LPS. 12
Previous studies also showed that TMP played neuroprotective and antioxidative roles in a number of animal models of ocular diseases, entailing oxygen-induced retinopathy and diabetic retinopathy.13,14 However, the role of TMP in anti-inflammatory effect in ocular tissue is not clear. In addition, the mechanisms underlying the beneficial effects of TMP in inflammation, especially in ocular inflammation, remain unknown.
The transcriptome is the complete aggregate of transcripts for a specific type of cells or tissues in a certain developmental stage or physiological condition. 15 Transcriptomic analysis can not only reveal the organism's gene expressing level and structural variation but also discover new genes.16,17 As a new transcriptome research method, RNA-sequencing (RNA-seq) with next-generation sequencing can detect the sequence of each transcription fragment directly and provide an abundance of transcripts in a given specimen. 18 Besides, compared with hybridization, RNA-seq showed little background noise and cross reaction, giving a relatively unambiguous representation of transcriptome. Moreover, RNA-seq is exceedingly reproducible and can cover a much greater dynamic range than microarray analysis. 16 Therefore, this high-throughput sequencing proved to be one of the quantitative and powerful methods to distinguish the variations of diseases and reveal their pathogenesis.
In this study, retinal transcriptome analysis of the protective effects by local treatment with TMP on EIU mice was performed using RNA-seq to investigate a comprehensive and comparative profile of gene expression changes mediated by TMP treatment. We found TMP attenuated the inflammation in EIU mice and it may become a novel therapeutic drug for ocular inflammation.
Methods
Animals and experimental procedures
Healthy adult female BALB/c mice (6–8 weeks old) were purchased from the Jackson Laboratories (Bar Harbor, ME) and maintained in an air-conditioned room with dim cyclic light (12 h on/off) in the Laboratory Animal Center of Chongqing Medical University (Chongqing, China). Food and water were supplied ad libitum. The experimental protocols, approved by the Ethics Committee of the First Affiliated Hospital of Chongqing Medical University, adhered to the ARVO Statement for the Use of Animals in Ophthalmic and Vision Research. Every effort was made to minimize animal discomfort and stress. The 6- to 8-week-old mice were randomly divided into LPS group and TMP+LPS groups. The TMP+LPS group: mice were anesthetized and 125 ng of LPS dissolved in 1 μL sterile phosphate-buffered saline (PBS) was injected intravitreally into each mouse eye to induce EIU, as previously described. 19 After injection, 10 μL of 0.1% TMP (Sigma-Aldrich, St. Louis, MO) eye drops prepared in sterile PBS was given to both eyes six times. The LPS group: 125 ng of LPS was injected intravitreally into each mouse eye when mice were anesthetized, and 10 μL of sterile PBS was given six times. Twenty-four hours after LPS injection, the retinas were dissected for further assays.
Inflammation evaluation and clinical score
Twenty-four hours after LPS injection, mice were subjected for an evaluation of the clinical manifestation in EIU by a slit-lamp microscopy (Shangbang, Chongqing, China) and clinical scores were evaluated at the same time by two independent observers in accordance with the previously described criteria. 19
Library construction and RNA-seq
Total RNA was isolated from the retinas by TRIzol reagent (Invitrogen). The contaminating genomic DNA was eliminated by RNase-free DNase I (New England Biolabs, Ipswich, MA). Each sample contained 3 μg RNA. We created the library under the guidance of standard Illumina protocols. Sequencing libraries were constructed using the NEBNext® Ultra™ RNA Library Prep Kit for Illumina (New England Biolabs), following the manufacturer's instruction. Double-stranded complementary DNAs (cDNAs) were synthesized and fragmented, and the RNA-seq libraries were established according to the standard Illumina protocol. The quality of the library was evaluated by an Agilent 2100 Bioanalyzer (Agilent Technologies, Palo Alto, CA). An Illumina Hiseq2500 device was used to sequence the libraries. The RNA-seq was performed by Novogene (Beijing, China). Base calls were carried out by CASAVA to analyze the data. The split read aligner TopHat (v2.0.12) was used to align the reads to the genome with default parameters. Meanwhile, the mismatches of the genome were set at 2. The RNA-seq data have been uploaded to NCBI database under the BioProject accession PRJNA509762.
RNA-seq data analysis
DESeq R package (1.10.1) was used to analyze the differentially expressed genes (DEGs) of LPS group and TMP+LPS group (three biological replicates for each group). The Benjamini and Hochberg's approach was applied to adjust the P values and to control the false discovery rate. The adjusted P < 0.05 genes were assigned as differentially expressed.
Gene ontology and Kyoto Encyclopedia of Genes and Genomes pathway enrichment analysis of DEGs
The gene ontology (GO)seq R package, ensuring the correction of gene length bias, was used to carry out the GO enrichment analysis of DEGs. Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway analysis and functional annotation for differential genes were performed using KOBAS software (v2.0), which is an efficient examination software to evaluate the significance of enrichment of pathways. An absolute threshold corrected P < 0.05 was used to select significantly enriched GO terms and pathways.
Protein–protein interaction analysis of DEGs
The protein–protein interaction (PPI) network was constructed using the STRING software v10.0 (www.string-db.org) and was visualized with Cytoscape software (http://cytoscape.org/) to reveal the connection among the DEGs. The subnetwork with P value <0.05 was assigned as significant.
Real-time quantitative PCR
The samples used in the real-time PCR validation experiments were independent from the RNA-seq. Total RNA was isolated from mice retinas using the TRIzol reagent (Invitrogen) according to the manufacturer's instructions. cDNA was synthesized using the PrimeScript RT Reagent Kit (Takara Biotechnology, Dalian, China). Real-time PCR validation of chosen DEGs was performed according to the manufacturer's instructions of ABI Prism 7500 system (Applied Biosystems, Foster City, CA). PCR amplification was conducted in a volume of 20 μL using SYBR Premix Ex TaqTM II (Takara Biotechnology). The cycling protocol consisted of one cycle of 10 min at 95°C followed by 40 cycles of 95°C for 15 s and 60°C for 1 min. Each sample was tested in duplicate and the endogenous reference gene GAPDH was used to normalize the messenger RNA expression. Relative quantification was achieved by the average cycle threshold values as previously described. 19 The primer sequences and accession numbers are shown in Table 1.
Sequences of the Primers for Real-Time PCR
Statistical analysis
Data are shown as mean ± SEM. Real-time PCR data were analyzed by SPSS 22.0 software (SPSS, Inc., Chicago, IL) and GraphPad Prism 5 software (GraphPad Software, Inc., San Diego, CA). The experimental data were analyzed by independent-sample t-test. The P < 0.05 was seen as significantly different.
Results
TMP exerted anti-inflammatory effect in EIU
Twenty-four hours after LPS injection, inflammation responses in the anterior chamber were observed under a slit lamp. The clinical scores were assessed according to the standard described previously. 19 The LPS group manifested typical signs of uveitis, such as remarkable iris hyperemia and hypopyon. The inflammation was notably alleviated in the TMP-treated EIU eyes. The clinical scores showed a remarkable reduction in the TMP+LPS group (2.67 ± 0.78, mean ± standard deviation [SD]) compared with the LPS group (3.83 ± 0.75, mean ± SD, P < 0.001) at 24 h after injecting LPS (Fig. 1).

TMP reduced the anterior chamber inflammation and lowered clinical scores at 24 h after injecting LPS.
Analysis of DEGs
To reveal the changes of retinal gene expression between the TMP-treated and the untreated EIU mice, computational analysis of the DEGs was performed. Genes with corrected P < 0.05 were considered DEGs. The results showed that 407 genes transformed expression in the LPS group compared with TMP+LPS group. Within the DEGs, 356 (87.5%) genes were upregulated (Fig. 2, red dots) and 51 genes (12.5%) were downregulated (Fig. 2, green dots) in the TMP-untreated EIU mice compared with the TMP-treated EIU mice. In our previous study, we showed that the expression of 478 genes altered in the retinas of the EIU mice versus the control mice. 16 In this study, we found that TMP alleviated the expressions of 21 (4.4%) genes among these 478 DEGs, which were regulated by LPS. The 21 genes included Slc19a1, Hif3a, Cdt1, Tc2n, Fkbp5, Lamc3, Pgpep1l, Usp2, Prr5, Slc6a20a, 8430408g22rik, Lrrc8c, B4galt6, Kl, E230001n04rik, Zbtb16, Selenbp1, Selenbp2, Gm807, Gm10807, and Ac156953.1.

Volcano plot of the up- and downregulated DEGs. The number of the differential genes between the TMP-treated and nontreated group was shown in the volcano plots. The horizontal axis is log2-fold change, and the vertical axis represents the P value after the negative logarithm conversion. The red dots indicate the upregulated genes, whereas the green dots represent the downregulated genes. The corrected P < 0.05 was an absolute threshold used to select DEGs. DEGs, differentially expressed genes.
GO enrichment analysis of DEGs between TMP-treated and untreated EIU mice
The GO annotations of the DEGs in the nontreated group compared with the TMP-treated group were involved in three major categories: biological processes, cellular components, and molecular function. In addition, the top 30 enriched up- and downregulated GO terms in the LPS group are shown in Fig. 3. An absolute threshold of the corrected P < 0.05 was applied to select 12 remarkably upregulated GO terms, including immune system process, immune response, antigen processing and presentation, GTPase activity, GTP binding, guanyl ribonucleotide binding, guanyl nucleotide binding, major histocompatibility complex (MHC) protein complex, MHC class I protein complex, chemokine activity, chemokine receptor binding, and transition metal ion transport (Table 2). These significantly upregulated GO terms of LPS group were mainly related to immune response process-related GO terms, manifesting that TMP may play an anti-inflammatory role by downregulating the genes from these immune response process-related GO terms with the treatment of TMP. No significantly downregulated GO terms have been enriched (Fig. 3).

GO enrichment of DEGs. The top 30 up- and downregulated GO enriched terms in the TMP-treated mice compared with untreated EIU mice. The horizontal axis is the enriched GO term and the vertical axis represents the number of significant genes in the term. Green marks biological process, orange indicates cellular component, and purple represents molecular function. The corrected P < 0.05 was an absolute threshold used to select significantly enriched GO terms (*adjusted P < 0.05). GO, gene ontology.
The Significantly Enriched Gene Ontology Terms
GO, gene ontology; MHC, major histocompatibility complex.
KEGG pathway analysis of DEGs between TMP-treated and untreated EIU mice
KEGG pathway was enriched to identify the possible pathways involved in the protective effect of TMP. The top 20 upregulated pathways of LPS group compared with TMP+LPS group are shown in Fig. 4. The results showed that 27 significantly upregulated enriched pathways in the aggregate were found in the LPS group compared with TMP+LPS group, including phagosome, antigen processing and presentation, allograft rejection, herpes simplex infection, graft-versus-host disease, type I diabetes mellitus, autoimmune thyroid disease, tuberculosis, Staphylococcus aureus infection, viral myocarditis, cell adhesion molecules (CAMs), leishmaniasis, human T-cell lymphotropic virus type I (HTLV-I) infection, rheumatoid arthritis, influenza A, intestinal immune network for immunoglobulin A (IgA) production, malaria, viral carcinogenesis, cytosolic DNA-sensing pathway, asthma, measles, natural killer cell-mediated cytotoxicity, osteoclast differentiation, pertussis, inflammatory bowel disease (IBD), toxoplasmosis and Epstein–Barr virus infection. No downregulated pathway has been enriched (Table 3).
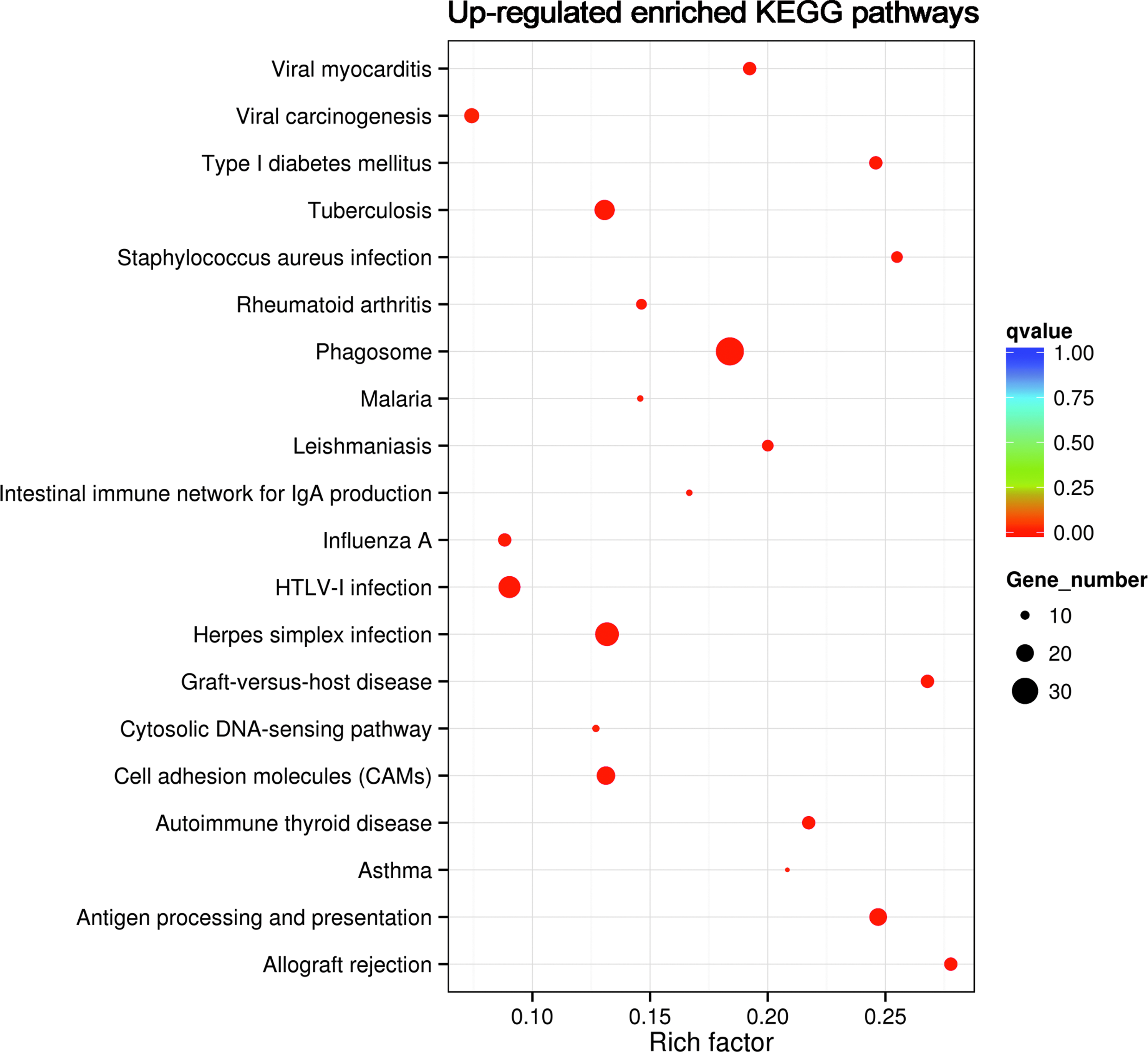
KEGG pathway analysis of DEGs. The top 20 upregulated KEGG pathways associated with the identified DEGs between the LPS group and the TMP+LPS group. The horizontal axis and the vertical axis indicate the log (corrected P value) and the function descriptions of the enriched pathways, respectively. The corrected P < 0.05 was an absolute threshold used to select significantly enriched pathways (*adjusted P value <0.05). KEGG, Kyoto Encyclopedia of Genes and Genomes.
The Most Enriched Kyoto Encyclopedia of Genes and Genomes Pathways
CAMs, cell adhesion molecules; HTLV-I; IBD, inflammatory bowel disease; IgA, immunoglobulin A.
PPI network for DEGs
The PPI network of the DEGs in LPS group compared with TMP+LPS group was constructed by Cytoscape based on STRING database. The constructed PPI network consisted of 386 nodes and 2,393 edges (Fig. 5A). Among the 42 networks, the top 3 significant subnetworks with P value <0.05 are shown in Fig. 5B–D. The top 10 high degree genes, including Cxcl10 (degree = 75), Irf7 (degree = 69), Oasl2 (degree = 63), Tlr2 (degree = 59), Oas2 (degree = 58), Ifit1 (degree = 57), Isg15 (degree = 56), Cxcl9 (degree = 56), Ifit3 (degree = 56), and Irf1 (degree = 55), were the hub genes (Table 4).
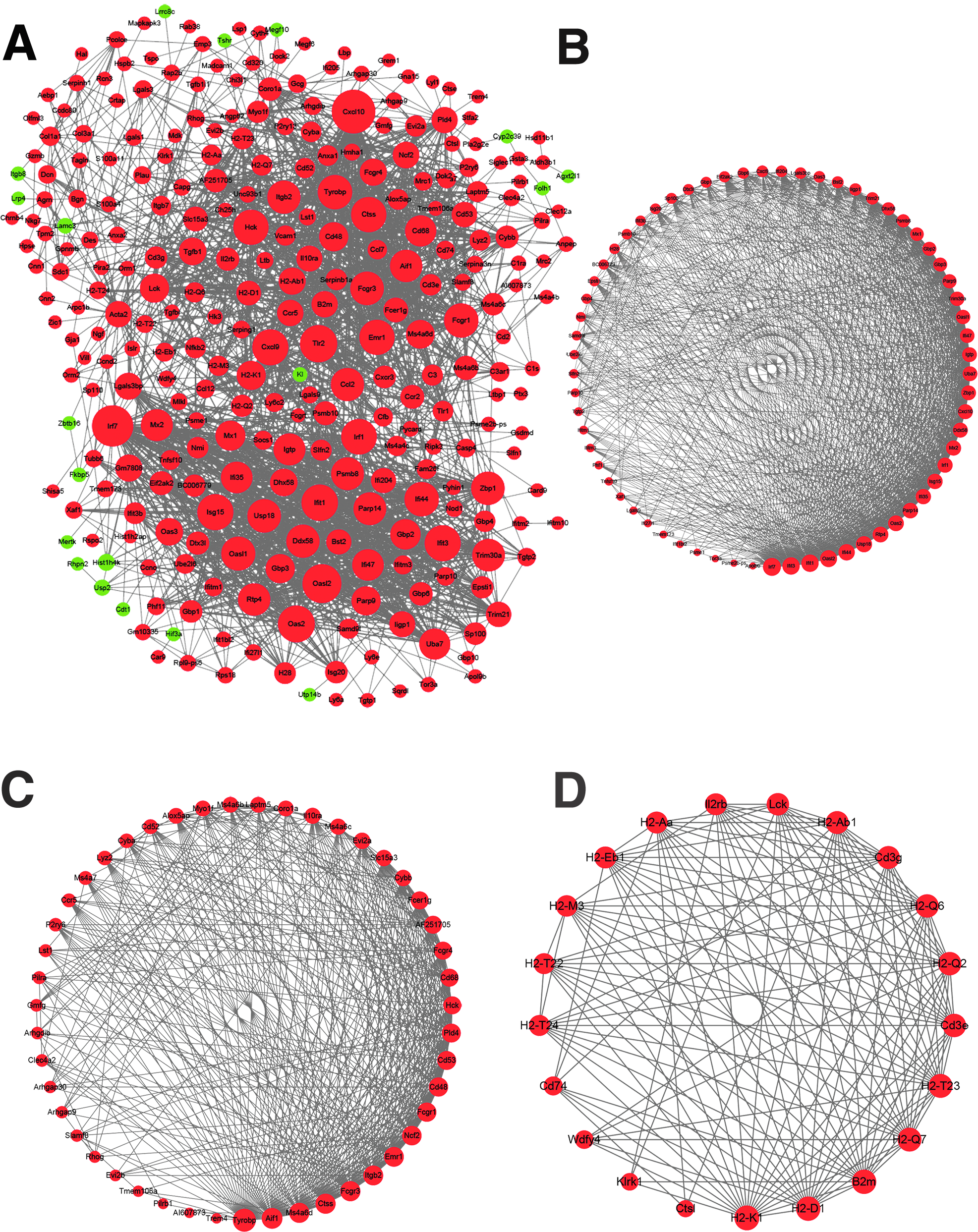
The PPI networks of DEGs.
The Top 10 High-Degree Genes of Protein–Protein Interaction Network
Real-time PCR validation of the DEGs
To confirm the differential gene expression results from transcriptome analysis, seven genes (log2-fold change >2 and adjusted P value <0.05) through GO and KEGG analyses were selected to be validated by real-time PCR (Table 5). Since there were no significantly enriched downregulated GO term and KEGG pathway, all of the seven genes were chosen from the upregulated genes in TMP-untreated mice compared to TMP-treated EIU mice. No DEGs met the criteria log2-fold change >2 and adjusted P value <0.05 in the significantly downregulated GO terms and no downregulated pathways were enriched. The seven chosen genes can be divided into three types: inflammation-related genes: Ccl12, Ccl2, and Gzmb, the complement system-related gene: C1ra, and interferon (IFNs)-related genes: Siglec1, Zbp1, and Ifit1. The results showed that real-time quantitative PCR-measured genes exhibited a similar pattern as the RNA-seq analyses. Compared with the TMP+LPS group, whole validated genes, including Ccl12, Ccl2, Gzmb, C1ra, Siglec1, Zbp1, as well as Ifit1, were remarkably upregulated in the LPS group (P < 0.05, P < 0.01) (Fig. 6).

Validation of DEGs with real-time quantitative PCR. All the genes to be validated were selected from the remarkably enriched upregulated GO terms and KEGG pathways. Compared with the LPS group, the mRNA expressions of inflammation-related genes Ccl12, Ccl2, Gzmb
Real-Time PCR Validation of Differentially Expressed Genes
Discussion
We demonstrated an array of DEGs in the TMP-treated and untreated EIU mice. Meanwhile, we identified the relevant molecular pathways to explore the possible molecular mechanism by which TMP eye drops protect against EIU. The results showed that the inflammatory response, the complement system, as well as the IFN-related genes are associated with the potential protective mechanisms of TMP eye drops' protective effect. The TMP treatment targeting these genes may exert a commendably therapeutic effect in uveitis.
The Ccl2, Ccl12, Gzmb, C1ra, Siglec1, Zbp1, and Ifit1 are the top seven genes with the greatest fold changes after TMP treatment. Chemokine (C–C motif) ligand 2 encoded by the Ccl2 gene, also known as the monocyte chemoattractant protein-1, is an important inflammatory mediator of uveitis. It attracts various inflammatory cells, such as monocytes, neutrophils, memory T cells, and natural killer cells, to the site of inflammation. 20 Chemokine (C–C motif) ligand 12 (Ccl12) is a proinflammatory cytokine responsible for fibrocyte recruitment and intensifies fibrotic responses. It prolongs inflammation and inhibits fibroblasts, and prevents initiation of the reparative response. 21 Granzyme B (Gzmb) is a cytotoxic mediator that induces target cell death 22 and plays an important role in several chronic inflammatory diseases.23,24 Complement component 1r subcomponent A (C1ra) is the homologue of human C1r, and C1r is a highly specific serine protease that initiates the classical pathway of complement activation during immune and inflammation responses.25–27 Sialic acid binding Ig-like lectin 1, sialo adhesin (Siglec1), is a key IFN factor of uveitis in animal models. 28 This factor combines the ability to attach and internalize certain viruses and endocytose sialylated bacteria such as Neisseria meningitidis. 28 Z-DNA binding protein 1 (Zbp1), identified as an IFN-induced cytosolic DNA sensor, 29 can trigger cell death and inflammatory responses during virus infection. 30 IFN-induced protein with tetratricopeptide repeats 1 (Ifit1) serves as a viral restriction factor through regulating virus replication 31 and inhibiting steps in translation initiation during responses to viral infections.32,33 In the present study, we found that these seven genes were remarkably decreased in the TMP-treated mice compared with the untreated EIU mice, indicating that the therapeutic effect of TMP eye drops may be associated with these genes.
TMP alleviated the inflammatory responses and the severity of clinical signs of EIU mice. Through the RNA-seq analysis, the GO functional classification, and enrichment analysis, as well as the KEGG pathway enrichment analysis, we found that three types of genes were remarkably downregulated in the TMP-treated group, including the inflammation-related genes (Ccl12, Ccl2, and Gzmb), complement system-related gene (C1ra), and IFN-related genes (Siglec1, Zbp1, and Ifit1). Previous studies showed that TMP exerted an anti-inflammatory effect in multiple organs and systems,34–36 and reduced the release of reactive oxygen species production induced by IFN-γ. 37 In parallel with previous studies, we showed a significant decrease in the inflammation-related genes (Ccl12, Ccl2, and Gzmb) and IFN-related genes (Siglec1, Zbp1, and Ifit1).37,38 Nevertheless, the present study is the first to show that TMP may exert an anti-inflammatory effect in ocular inflammation by downregulating the complement system-related genes. It may shed light on finding a novel possible molecular mechanism by which TMP protects against ocular inflammation.
Protein–protein interaction network evaluation assisted to identify the Cxcl10 (degree = 75), Irf7 (degree = 69), Oasl2 (degree = 63), Tlr2 (degree = 59), Oas2 (degree = 58), Ifit1 (degree = 57), Isg15 (degree = 56), Cxcl9 (degree = 56), Ifit3 (degree = 56), and Irf1 (degree = 55), the hub genes. The three significant subnetworks, shown in Fig. 5B–D, were related to the GO terms (immune system process, immune response, antigen processing and presentation, GTP binding, and MHC class I protein complex) and KEGG pathways (phagosome, antigen processing and presentation, allograft rejection, herpes simplex infection, graft-versus-host disease, type I diabetes mellitus, autoimmune thyroid disease, tuberculosis, S. aureus infection, viral myocarditis, CAMs, leishmaniasis, HTLV-I infection, rheumatoid arthritis, influenza A, intestinal immune network for IgA production, malaria, viral carcinogenesis, asthma, measles, IBD, toxoplasmosis, and Epstein–Barr virus infection).
Our study defined a potential novel therapeutic medicine for ocular inflammation. However, we still noticed some limitations in our study that need to be addressed in the future. First, the transcriptional changes of retinas were analyzed 24 h after injecting LPS, when the clinical and histological scores are the most obvious. The samples taken at a single time point could not reveal the dynamic changes mediated by TMP. Therefore, studies, including more time points, would provide detailed information. Second, the transcriptome profile also included the afflux of hemocytes from the posterior segment. Thus, the transcripts also incorporate these cells, which is more relevant to uveitis in human patients. However, it may be useful to eliminate the influence of these infiltrated cells by perfusion of the retina before collecting the samples.
In summary, RNA-seq was a complete and accurate method for a comprehensive and comparative analysis of the retinal gene expression changes in probing the mechanisms of therapeutic effect in EIU. Local application of TMP inhibited retinal inflammatory responses and downregulated the expression of several inflammation-related genes, suggesting that TMP may be a promising potential therapeutic medicine for ocular inflammation.
Footnotes
Acknowledgments
The authors thank the National Natural Science Foundation of China for financial support from grants (81770949 and 81470621), the Henan Science and Technology Bureau (182102310145), and the National Key Clinical Specialties Construction Program of China. The authors alone are responsible for the content and writing of the article.
Authors' Contributions
L.Y., Y.Q., B.H., and B.L. conceived the idea and designed the experiments. L.Y., Y.Q., R.L., P.Y., and X.F. performed the experiments. L.Y., J.L., B.H., and Y.Q. analyzed the data. L.Y. and Y.Q. wrote the article. B.L. reviewed and revised the article. All authors read and approved the final version of the article.
Author Disclosure Statement
The authors declare that they have no conflicts of interest.