
Abstract
Purpose:
Transforming growth factor-β2 (TGFβ2) and Toll-like receptor 4 (TLR4) crosstalk have been implicated in extracellular matrix regulation in the trabecular meshwork (TM) and ocular hypertension in mice. We investigated TLR4 expression in normal and glaucomatous human trabecular meshwork (HTM) sections and utilized a human perfusion organ culture model to determine TGFβ2-TLR4 signaling crosstalk in glaucoma.
Methods:
Expression of TLR4 was determined in TM of normal and glaucomatous human eyes. Anterior segments of paired human eyes were perfused at a constant flow rate (2.5 μL/min) for 4 days to acquire stable baseline intraocular pressures (IOPs). We treated paired eyes with two different treatment paradigms: (1) TGFβ2 in one eye and vehicle control in the paired eye, (2) TGFβ2 in one eye and TGFβ2 + TLR4 inhibitor TAK-242 in the paired eye. Perfusate and TM tissue were collected and analyzed for fibronectin (FN) and collagen IV (COLIV) expression.
Results:
We observed increased TLR4 expression in glaucomatous HTM sections compared to normal (age-matched) (P < 0.05). Significant elevation of IOP was detected in 47% of TGFβ2-treated anterior segments (P < 0.01) compared to control, and in TGFβ2 treated compared with co-treatment with TGFβ2 + TLR4 inhibitor (P < 0.0001). An increase in FN and COLIV expression was observed after TGFβ2 treatment, and inhibition of TLR4 signaling decreased TGFβ2-induced FN and COLIV expression in perfusate (P < 0.05).
Conclusions:
These studies identify TGFβ2-TLR4 crosstalk as a novel pathway in glaucoma. They provide a potential new target to lower IOP and explore glaucoma pathogenesis.
Introduction
Elevated intraocular pressure (IOP) is a major risk factor in glaucoma. The trabecular meshwork (TM) is a crucial tissue in aqueous humor (AH) drainage. 1 A fine balance regulates IOP through AH production from the ciliary epithelium and outflow through the TM. 2 In primary open angle glaucoma (POAG), the increase in IOP is mainly because of the dysregulation of outflow facility causing pathological changes in the TM. The changes can be brought about by multiple factors but one key factor that has been implicated in POAG has been transforming growth factor-β2 (TGFβ2). The cytokine has been shown to be elevated in the AH of POAG patients.3,4 It can alter the extracellular matrix (ECM) composition and induce crosslinking within the TM.5–7 Various studies have shown TGFβ2 to elevate IOP in anterior segment perfusion organ culture systems.8,9 These reports, in conjunction with expression data from human trabecular meshwork (HTM) cells, implicate TGFβ2 in TM fibrosis and elevation of IOP.
Toll-like receptor 4 (TLR4), a member of the TLR family, has also been implicated in fibrosis. 1 It was originally identified as a specific receptor for lipopolysaccharide.10,11 Other endogenous ligands, such as damage-associated molecular patterns (DAMPs) expressed as a result of injury, cell damage, ECM remodeling, and oxidative stress also activate TLR4 signaling.12,13 Previous reports have linked DAMP-activated TLR4 signaling with the regulation of ECM and fibrosis. 14 DAMPs can activate TLR4 to augment TGFβ2 signaling for downstream pathogenic fibrotic responses as observed in hepatic fibrosis, renal fibrosis, and in scleroderma patients.14–16 It has also been shown that the C3H/HeJ (Tlr4 mutant) mice have a missense point mutation in the Tlr4 gene rendering the receptor nonfunctional. These Tlr4 mutant mice develop less interstitial fibrosis in contrast to wild-type mice. 17 In addition, specific single nucleotide polymorphism alleles in Tlr4 have been shown to delay progression of fibrosis in liver disease and confer protection. 18 Recently, we also discovered TLR4 signaling as an important regulator of ocular hypertension and TM damage in mice and primary HTM cell cultures. 1 These data implicate TLR4 signaling in ECM regulation.
A potential antifibrotic approach for selective therapeutic targeting of TLR4 signaling is TAK-242. It is a novel cyclohexene derivative that specifically suppresses TLR4 signaling, inhibiting production of nitric oxide and proinflammatory cytokines. 19 It blocks the intracellular signaling domain of TLR4 and potently suppresses both ligand-dependent and -independent signaling of TLR4. 20 In preclinical fibrosis models and in fibroblasts, TAK-242 treatment has been shown to reduce the expression of several profibrotic mediators. 18 Previously, we reported that TLR4 inhibitor, TAK-242, inhibits the expression of ECM markers, laminin, collagen I, and fibronectin (FN) in TM cells treated with TGFβ2. 1 We have established that the TLR4 signaling is an important pathway to target the pathogenesis involving TLR4 in the TM. Small molecule TLR4 signaling inhibitors could potentially provide new therapies for glaucoma.
Human anterior segment perfusion organ culture systems provide a unique ex vivo model to study the pathophysiology of elevated IOP observed in glaucoma pathogenesis. We studied the effects of TGFβ2 and TLR4 crosstalk in normal and glaucomatous eyes and within the perfused human eyes in the anterior segment organ culture model. We wanted to determine whether TGFβ2 elevates IOP in this experimental model and if a selective TLR4 inhibitor could rescue the elevation in IOP and reduce the secretion of FN. Our studies will determine this crucial crosstalk role of TLR4 signaling in controlling IOP and provide a potential pharmacological agent that can significantly reduce the effect of TGFβ2 in human anterior segment perfusion organ cultures. Identification of a unique target will provide a novel therapy to lower IOP in patients with POAG.
Methods
Immunohistochemistry of HTM sections
All human donor eyes were obtained according to the provisions of the Declaration of Helsinki for research involving human tissue. Donors with a history of ocular disorder other than glaucoma were not included in the study. Anterior segments from glaucomatous and normal human donor eyes were formalin-fixed within 6-h postmortem and embedded in paraffin to produce sagittal TM tissue planes. Sections were masked for primary data analysis of immunohistochemical staining. Paraffin-embedded segments were deparaffinized (100%, xylene 95% ethanol, and 50% ethanol) and rinsed using phosphate-based saline buffer and blocked with SuperBlock™ T20 (TBS) Blocking Buffer for an hour at room temperature. Tissue sections were incubated overnight at room temperature with primary antibody (anti-TLR4, ab13556, 1:100) dissolved in SuperBlock (Catalog No. 37515; ThermoScientific, Rockford, IL). After incubation, tissues were washed three times with phosphate-buffered saline (PBS). Secondary antibody was applied for an hour to the sections at room temperature (Alexa Fluor 488 goat anti-rabbit, A11008, 1:500). After incubation, tissues were rinsed with PBS and cover-slipped with Prolong Gold Antifade Reagent containing DAPI (4’,6-diamidino-2-phenylindole) nuclear stain (Catalog No. P36931; Life Technologies, Carlsbad, CA).
Immunohistochemistry imaging was performed using the Keyence microscope (Catalog No. BZ-X710; Keyence, Itasca, IL) in a masked fashion. TM images were captured with a 20 × objective lens. To evaluate the effect of TLR4 in the glaucomatous TM, we excluded any human donor eye section that highly expressed TLR4 in other ocular tissues such as the cornea and ciliary body. This allowed the ability to remove any potential donors that had ocular inflammation owing to other unidentified means. The staining of TLR4 was quantified in a masked manner using ImageJ analysis to identify the range of florescence intensity/area for each TM section from normal and glaucomatous eyes.
Anterior segment perfusion culture
Donors with a known a history of glaucoma or other ocular disorders were not included in the study. All eyes were obtained according to the provisions of the Declaration of Helsinki for research involving human tissue. Human eye bank eyes that were aged matched (Lions Eye Institute for Transplant and Research, Tampa, FL) were obtained within 12 h of death, dissected in half, and placed in culture medium before being shipped overnight on ice. Within 24 h of death, the anterior segments were cultured in the human anterior segment perfusion culture system. Perfusion culture was setup according to previously published protocols.2,21
In brief, under sterile conditions, anterior segments of human eyes were placed on the perfusion model culture dishes (ethylene oxide sterilized) with a tight-fitting ring to seal the scleral rim to an inner island to create a “closed eye” system using bolts. The infusion of medium was through a 30 mL sterile syringe connected to a 3-way stop cock, a 2.5 μL micropore syringe filter, a 15 mm luer lock stub, and polyethylene tubing connected to an infusion port on the chamber dish. A second port was fitted with tubing and a pressure transducer to constantly measure IOP. Before connecting the chamber dish to a pressure transducer and Powerlab system (ADInstruments, Inc., Colorado Springs, CO), an initial supply of medium was injected into the eye and trapped air removed through the “push-pull” method using the above described 30 mL sterile syringes.
Once the eye was sealed in place and all air bubbles removed, the eye was scored with an artificial slit 3 mm posterior to the limbus to sever aqueous veins to prevent impedance of aqueous flow. The infusion syringes were then fitted on the Harvard Apparatus microinfusion pump (Harvard Apparatus, Cambridge, MA). Medium (serum-free high glucose Dulbecco's Modified Eagle Medium containing 1% penicillin + streptomycin and 1%
Baseline pressures were documented for 4 days. Treatment was performed in paired eyes through the perfusion medium. Each paired eye was utilized for either experimental condition (1) No treatment control versus active recombinant human TGFβ2 (5 ng/mL) 9 (Catalog No. 302-B2-010; R&D Systems, Minneapolis, MN) for 4 days or (2) TGFβ2 (5 ng/mL) versus TGFβ2 (5 ng/mL) + TLR4 inhibitor (TAK-242, 15 μM) (Catalog No. CLI-095; Lifespan Biosciences, Seattle, WA) for 8 days. Change in mean daily (average of 24 h) IOP was calculated by normalizing to average baseline pressure. An average area under the curve (AUC) was measured for each experimental condition.
Western blot analysis of conditioned medium
The FN protein secreted by TM cells was determined by Western immunoblot analysis. Conditioned medium (10 mL) was collected from each human anterior segment perfusion organ culture after treatment with (1) control or TGFβ2 (5 ng/mL) and (2) after TGFβ2 (5 ng/mL) or co-treatment with TGFβ2 (5 ng/mL) + TLR4 inhibitor (TAK-242, 15 μM). One milliliter of conditioned medium was concentrated using 10 μL of StrataClean Resin (Catalog No. 400714-61; Agilent, Santa Clara, CA) from each collected conditioned medium sample. Concentrated protein was resolved on Novex® 4–20% Tris-glycine precast gels (Life Technologies, Grand Island, NY) and transferred to Immun-Blot PVDF membranes (BioRad, Hercules, CA). Blocking and antibody incubations were performed in SuperBlock (Life Technologies, Grand Island, NY). Membranes were incubated overnight at room temperature with anti-FN (AB1945, 1:1,000) or anti-COLIV (NB120-6586, 1:1,000). Primary antibody was identified using horseradish peroxidase (HRP)-conjugated secondary antibody (goat anti-rabbit, 7074S, 1:1,000). Detection of HRP-labeled secondary antibody was performed with ECL SuperSignal West Femto Maximum Sensitivity Substrate (Pierce Biotechnology, Rockford, IL). The bands were visualized using the BioRad VersaDoc multi-wavelength imaging system (BioRad) and Quantity One Imaging software (BioRad). Immunoreactivity was quantified using the ImageJ spot densitometry software.
Immunohistochemistry of perfusion TM sections
Half of the anterior eye segments postperfusion were fixed in formalin and paraffin embedded. Paraffin-embedded segments were deparaffinized (100%, xylene 95% ethanol, and 50% ethanol). Slides were washed with PBS for 10 min and blocked with SuperBlock Blocking Buffer at room temperature for 1 h. Sections were subsequently labeled with primary antibodies (anti-FN, AB1945, 1:100 and anti-TLR4, ab22048, 1:100). Primary antibodies were detected using Alexa Fluor secondary antibodies (Alexa Fluor 488 goat anti-rabbit, A11008, 1:500 and Alexa Fluor 594 donkey anti-mouse, A21203, 1:500). Cell nuclei were counterstained using DAPI. Stained sections were imaged using Keyence microscope (Catalog No. BZ-X710; Keyence). TM images were captured with a 20 × objective lens. Immunoreactivity was quantified using the ImageJ spot densitometry software. Sections that were outliers were removed from the quantification analysis.
Hematoxylin and eosin staining
Anterior segments of human eyes were formalin fixed and embedded in paraffin. The eyes were sectioned in produce sagittal TM tissue planes. They were processed an automated system (LEICA ST 5020, Buffalo Grove, IL) for deparaffinization (100%, xylene 95% ethanol, and 50% ethanol) and stained with hematoxylin and eosin.
The images were captured with the 20 × objective lens using a Keyence microscope (Catalog No. BZ-X710; Keyence) using the bright-field light source.
Statistical analysis
Grading scale was compared using the Mann–Whitney statistical analysis. Paired and unpaired Student's t-test was used to compare between treatment groups. Two-way analysis of variance was used to compare between samples of different treatment groups and intergroup differences between time points. GraphPad Prism 6 (San Diego, CA) was used to perform all statistical analyses. Data are presented as mean ± standard error of the mean and P < 0.05 was considered statistically significant.
Results
Increased expression of TLR4 in glaucomatous HTM
Previously, our group and others have reported expression of TLR4 in HTM and primary HTM cell cultures.1,23 Here, we demonstrate that TLR4 expression is elevated within the TM region of human glaucomatous donor samples compared with normal (age-matched) (Fig. 1A–F). The expression of TLR4 of glaucomatous samples was compared with normal and quantified. Graphical representation of the data exhibited significantly higher TLR4 expression in TM of glaucoma eyes compared with controls (P < 0.05, n = 13, 14) (Fig. 1G).
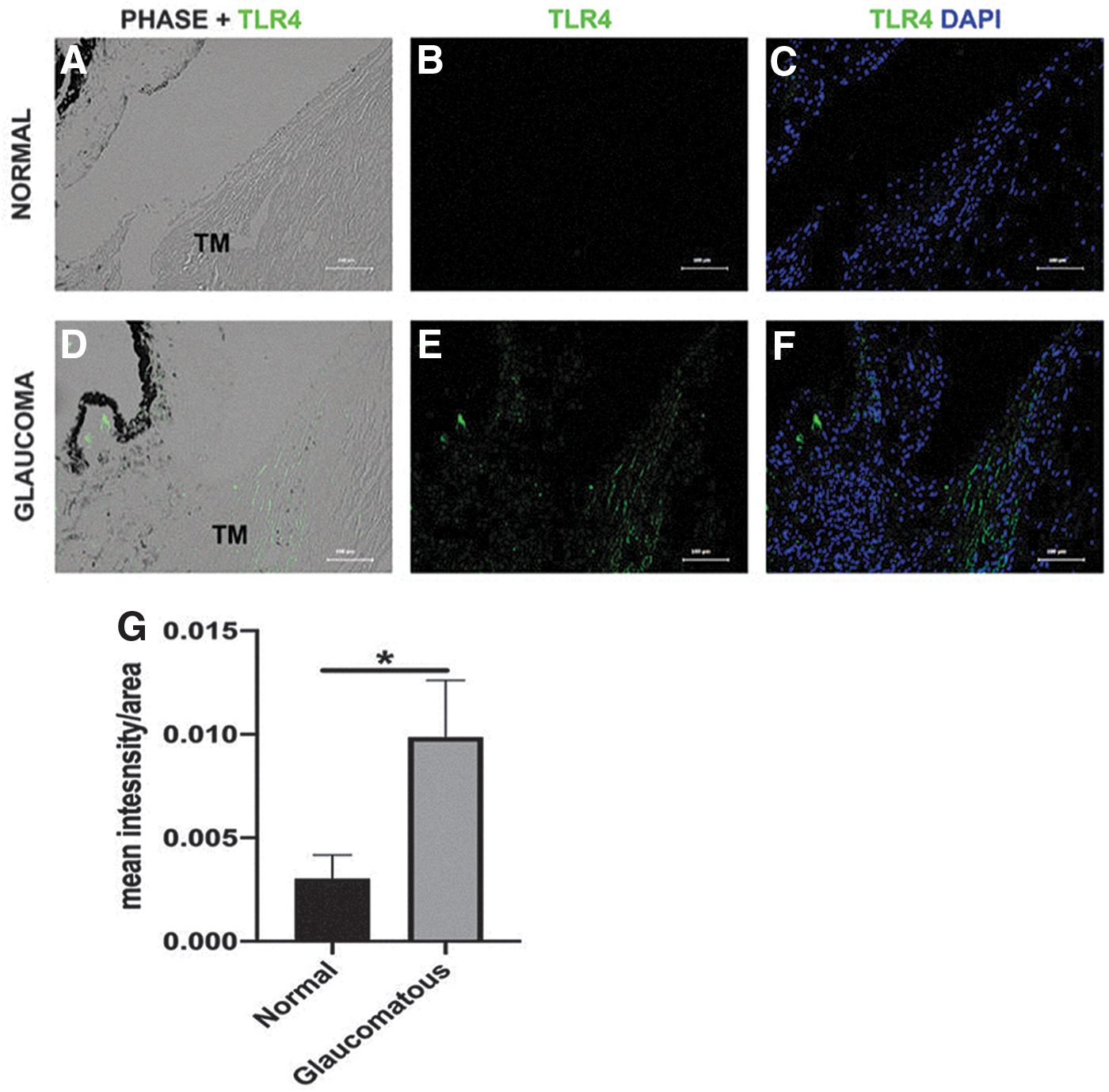
TLR4 expression in normal and glaucomatous HTM sections. Representative cross-sections from
Elevated IOP controlled through TLR4 inhibitor in human anterior segment perfusion cultures
It is well reported that TGFβ2 induces ECM production in the TM5,24–27 and elevates IOP in perfusion cultures8,9 and within our established mouse model of ocular hypertension using an Ad5.TGFβ2 virus.1,28,29 In addition, we previously demonstrated that a selective inhibitor, TAK-242, can block TGFβ2-induced TM damage in primary TM cell culture, and TAK-242 treatment alone (15 μM) had no effect on TM cells. 1 As previously reported, we also demonstrate as a positive control that in paired anterior segment perfusion cultures, TGFβ2 induces elevated IOP (Fig. 2A, B). 23 Of importance, we show that this elevation can be blocked by inhibition of TLR4 signaling through the selective inhibitor, TAK-242 (15 μM) (Fig. 2C, D, Supplementary Table S1). After 4 days post-treatment with TGFβ2 (5 ng/mL), there was significant increase in IOP (P < 0.01, n = 4) compared with control (Fig. 2A). The largest difference between means of both groups (TGFβ2 vs. control) by 4 days was 2.946 ± 0.7536 Δ in mmHg. The cumulative AUC for all IOP change through the time-course for TGFβ2 was significantly higher than control (P < 0.05, n = 4, AUC mean difference = 9.785 ± 3.031 Δ in mmHg) (Fig. 2B, Supplementary Table S2).

Elevation and control of IOP through constant perfusion of paired human anterior segments with various treatments. Graphical presentation of Δ in mmHg over baseline for human eyes treated with paired control and TGFβ2 (5 ng/mL)
For the TLR4 inhibitor, TAK-242, in co-treatment with TGFβ2, we extended the time course from 5 to 8 days to demonstrate a larger effect of TGFβ2 and highlight that co-treatment of TAK-242 + TGFβ2 blocked the elevation of IOP observed with TGFβ2 alone (P < 0.0001, n = 5) (Fig. 2C). The means between both groups (TGFβ2 vs. TGFβ2 + TLR4i) was −6.983 ± 1.936 Δ in mmHg. For the eyes that were 8 days in perfusion, the TLR4 inhibitor showed significantly decreased cumulative AUC IOP compared with TGFβ2 (P < 0.01, n = 3, AUC mean difference = −45.53 ± 8.024 Δ in mmHg) (Fig. 2D).
Inhibition of TLR4 signaling decreases TGFβ2-induced FN and COLIV expression in perfusate from human anterior segment perfusion organ cultures
Elevated IOP was observed in 47.4% (9/19) of all eyes that were treated with TGFβ2 and these eyes were termed as responders. For proof of concept and to confirm previous perfusion studies, TGFβ2 perfusate from responders of TGFβ2 and control samples that were not treated with TGFβ2 were analyzed for soluble FN and collagen IV expression. It has been well established that FN and collagen IV is increased in mouse TM, supernatant from cultured HTM cells, and perfusate from anterior segment perfusion cultures after TGFβ2 treatment (5 ng/mL).1,9,30,31
Consistent with these results, in the perfusate of responder eyes from our perfusion organ culture system, TGFβ2 induced an increase in FN (Fig. 3A) and collagen IV (Fig. 3B) expression compared with controls (n = 3). Images of all perfusate from responder eyes (n = 3) depicting expression of FN are given in Supplementary Fig. S1A and collagen IV in Supplementary Fig. S2A. Quantification of FN (Fig. 3A) and collagen IV (Fig. 3B) expression in responder eyes showed significantly elevated levels with TGFβ2 compared with control (P < 0.05, n = 3).
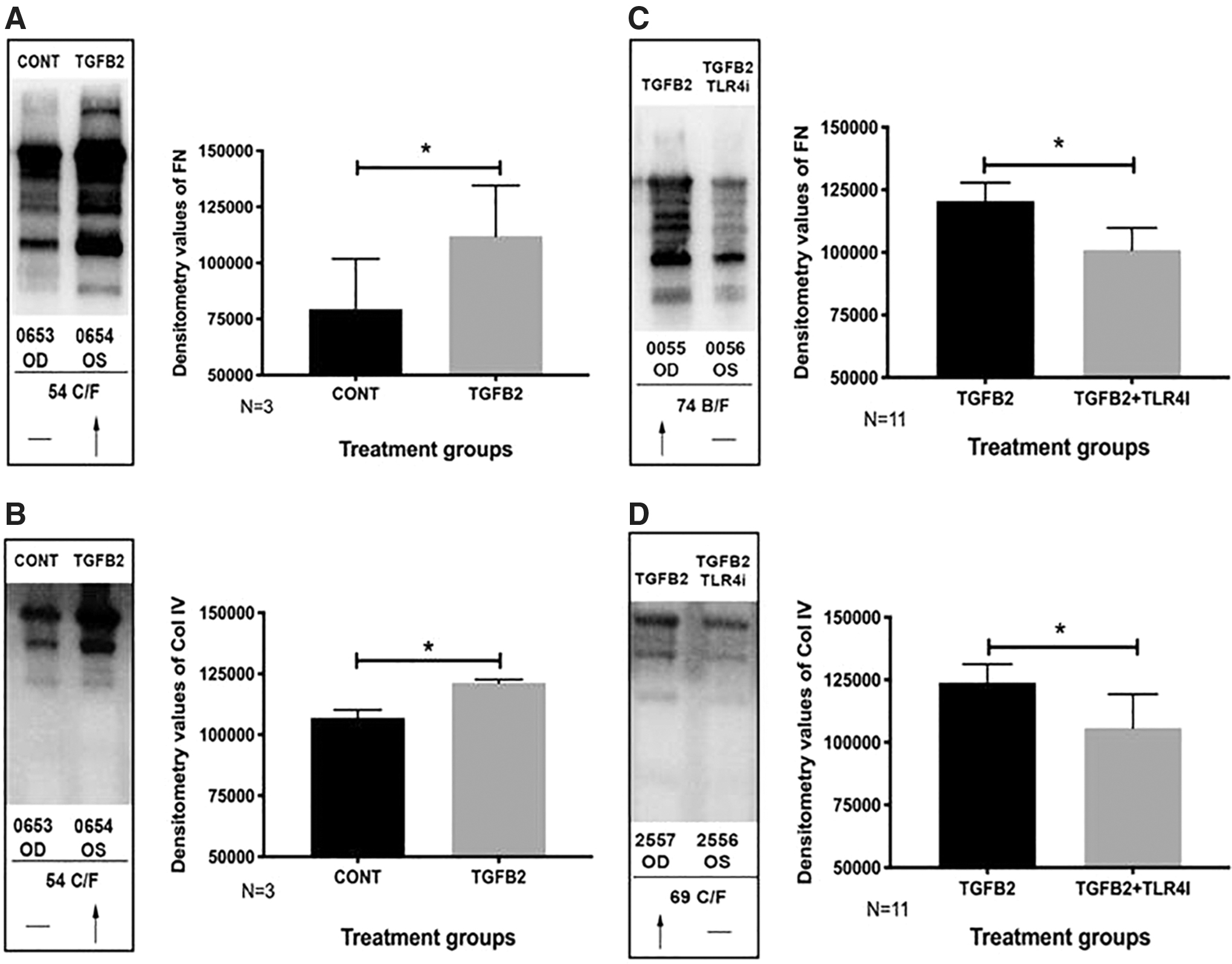
Inhibition of TLR4 signaling decreases TGFβ2-induced FN and COLIV expression in perfusate from paired anterior segments in human perfusion organ culture system. Western immunoblot of perfusate of responder eyes after TGFβ2 (5 ng/mL) treatment compared with control shows increased representative expression and significant densitometric analysis of
In contrast to eyes that responded to TGFβ2, there were also eyes that were nonresponders. This could potentially be because of the fact that the study was not extended beyond 8 days and the possibility of genetic variability in human samples. Not every eye responds in a similar manner and the study time-course may not have been sufficient to induce a substantial fibrotic response that could impede outflow facility and increase IOP in all eyes. Therefore, to understand the effects of TLR4 inhibition in terms of ECM deposition, we wanted to study the expression of FN and collagen IV on both responder and nonresponder eyes. The TGFβ2-induced elevation in FN (Fig. 3C) and collagen IV (Fig. 3D) level was significantly downregulated by TLR4 inhibitor, TAK-242 (15 μM), in both responder and nonresponder eyes (P < 0.05) (n = 11). Expression of FN and collagen IV in all perfusates after TGFβ2 treatment or in conjunction with TLR4 inhibition for responder and nonresponder eyes is given in Supplementary Fig. S1B and C and Supplementary Fig. S2A and C, respectively.
Decreased expression of FN in HTM after inhibition of TLR4 signaling
FN expression was identified within the TM of representative cross-sections from anterior segments collected after 4 days of control (Fig. 4A) or TGFβ2 treatment (Fig. 4B). Upon quantification of the FN levels, a trend of elevated expression was observed in all cross-sections for the responder group compared with the control (n = 3) (Fig. 4C). Comparable with the perfusate expression analysis between the TGFβ2 (Fig. 4D) and TGFβ2 + TAK-242 co-treatment group (Fig. 4E), FN levels showed a decreased trend in the TLR4 inhibitor (TAK-242) group (n = 8) (Fig. 4F). We only observed a trend and not a significant result for both quantifications as there is the possibility of segmental flow areas within the TM of these eyes and because of that there could potentially be FN differences and variability in expression.

Expression of FN after inhibition of TLR4 signaling and TGFβ2 treatment in paired anterior segments cultured through human perfusion organ system. Cross-sections of HTM show FN expression after
Discussion
We present an original report that depicts increased TLR4 expression in the TM of glaucoma patients. Our results distinguish the contributing role of TLR4 in ECM regulation and fibrosis within TM cells. In concurrence with identification of TLR4 in HTM fibrosis, we determined the important crosstalk between TGFβ2 and TLR4 signaling pathways in human glaucoma patient cohorts. Our data implicate TLR4 as a unique molecular mechanism that contributes to glaucomatous pathogenesis. Activation of TLR4 induces the formation of a complex and recruits adaptor proteins including MyD88. 32 The activation of MyD88-dependent pathway leads to translocation of nuclear factor kappa B (NF-κB) into the nucleus. 33 The nuclear NF-κB functions to regulate various genes including cytokines, chemokines, and proteins.32,33 It can activate ECM proteins and DAMPs. In various fibrotic diseases, DAMPs including cellular FN extra domain A (EDA), low-molecular-weight hyaluronan, and tenascin C have been shown to activate TLR4 and augment TGFβ signaling to activate downstream fibrotic responses15,33,34 (Fig. 5). Our discovery that inhibition of TLR4 decreases TGFβ2-mediated ECM accumulation to downregulate elevated IOP provides a novel therapeutic target that could target glaucomatous damage occurring within the TM.

Pathway of TGFβ2-TLR4 crosstalk within the glaucomatous TM. TLR4 is activated by endogenous ligands called DAMPs and signals through the MyD88/NF-κB signaling pathway to augment TGFβ2 signaling. The increased TGFβ2 signaling leads to an elevation in DAMP and ECM production to further activate TLR4 and downregulate BAMBI. This feed-forward loop and inhibition of BAMBI induced by the crosstalk between TGFβ2-TLR4 signaling causes progressive fibrosis within the TM. BAMBI, BMP, and activin membrane bound inhibitor; DAMPs, damage-associated molecular patterns; ECM, extracellular matrix; NF-κB, nuclear factor kappa B.
Similar to previous established reports, we demonstrated that TGFβ2 elevates IOP in anterior segment perfusion organ culture systems.8,9 It was interesting to note in our study that only 47.4% (9/19) of eyes treated with TGFβ2 induced an elevation in IOP. This response observed for TGFβ2 is a phenomenon that has also been observed in patients treated with glucocorticoids. The individuals displaying elevated IOP in response to glucocorticoids are referred to as steroid “responders” and those that are not as “nonresponders.”34,35 There have also been similar nonresponder and responder differences observed with IL-19 and IL-24 treatments in POC experiments. 36
Likewise, we observed that only a certain subset of donors was responsive to TGFβ2. Multiple factors could possibly be contributing to this unique finding. There is the possibility that because the study was not extended beyond 8 days and because of the genetic variability of human samples the study time-course may not have been sufficient to induce a substantial fibrotic response that could impede outflow facility and increase IOP in all eyes. This could also be attributed to genetic variability in samples, other external factors including incomplete TM rings because of removal of ciliary body, segmental flow patterns between the paired eyes, and potential epigenetic factors. Although we did not detect an elevation in IOP in the nonresponder eyes, it was still observed that certain nonresponders still showed elevation in FN and collagen IV expression with TGFβ2 treatment owing to the above-mentioned contributions. We wanted to emphasize this distinction and thus quantified all the responder and nonresponder eyes for the TGFβ2 versus TGFβ2 + TLR4i experimental cohort.
In addition, in our immunohistochemical analysis we identified that ∼23% of normal samples expressed TLR4. This could potentially implicate another mechanistic property of TLR4 that may be resistive to glaucomatous degeneration or the activation of TLR4 through a protective endogenous ligand. Furthermore, the donors may not have had examination to identify glaucoma phenotype and could possibly be undiagnosed glaucoma patients. There could also be underlying protective genetic factors that may be playing a role in reducing pathogenesis. On the contrary, ∼36% of glaucoma donor eyes did not depict any TLR4 expression. It has been shown that C3H/HeJ (Tlr4 mutant) mice have decreased fibrosis in contrast to wild-type mice. 17 There is a possibility that these donors may have TLR4 mutations that could potentially be rendering the receptor nonfunctional.
Several reports have shown that TGFβ2 induces FN expression in the TM.5,7,27 In addition, it has been reported that human perfused eyes decrease outflow facility and increase accumulation of ECM molecules.2,8,21 Similarly, in our study we observe comparable results of time-dependent elevated IOP after constant perfusion of ex vivo anterior segments with TGFβ2. Significant elevation in IOP is seen as early as 3 days post-TGFβ2 perfusion. As expected, we observed that inhibition of TLR4 signaling through the selective inhibitor TAK-242 produced a significant decline of TGFβ2-mediated IOP elevation. Furthermore, the decrease in FN expression after treatment with TAK-242 depicts the crucial role that TLR4 signaling plays in fibrosis and ECM deposition within the TM.
In conclusion, we successfully demonstrated that the anterior segment perfusion organ culture segments respond to TGFβ2 with concomitant increases in IOP and FN secretion. Modulation of TGFβ2-induced ECM production was performed by a selective TLR4 inhibitor to decrease elevated IOP. These results provide a novel and viable approach to manage glaucoma.
Footnotes
Acknowledgments
The authors thank Dr. Abbot F. Clark and Dr. Weiming Mao for their technical assistance with the perfusion organ culture model. The authors also thank the Lions Eye Institute for Transplant and Research (Tampa, FL) for providing the human donor eyes.
Author Disclosure Statement
The authors of the article have no potential conflicts of interest to disclose.
Funding Information
This project was funded by NIH R01EY026529 (C.M.M.). This study was supported in part by an unrestricted grant from Research to Prevent Blindness, Inc. to the UW Madison Department of Ophthalmology and Visual Sciences.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.