
Abstract
Purpose:
This study aimed to explore the effects of elevated KDM4D expression and potential therapeutic effects of Lycium barbarum polysaccharide (LBP) on pterygium.
Methods:
The expression levels of KDM4D in the primary pterygium (n = 29) and normal conjunctiva (n = 14) were detected by immunohistochemistry. The effects of KDM4D on pterygium fibroblasts were detected by the CCK-8 assay, liquid chromatography-mass spectrometry assay, flow cytometry, and scratch wound healing assay. The relative expression of KDM4D in pterygium fibroblasts stimulated by interleukin (IL)-1β, IL-6, IL-8, and LBP was detected by quantitative real-time PCR and Western blot. The effects of LBP on pterygium fibroblasts were detected using flow cytometry and scratch wound healing assays.
Results:
The expression level of KDM4D in pterygium was higher than that in normal conjunctiva. KDM4D increased the cell viability of pterygium fibroblasts. The differentially expressed genes identified in the LM-MS assay enriched in “actin filament organization” and “apoptosis.” KDM4D promoted migration and inhibited apoptosis of pterygium fibroblasts in vitro. Inflammatory cytokines, including IL-1β, IL-6, and IL-8, enhanced the expression of KDM4D in pterygium fibroblasts. LBP inhibited the expression of KDM4D in pterygium fibroblasts and decreased their cell viability. Moreover, LBP attenuated the KDM4D effects on migration and apoptosis of pterygium fibroblasts.
Conclusions:
Elevated KDM4D expression is a risk factor for pterygium formation. LBP inhibits the expression of KDM4D in pterygium fibroblasts and may be a potential drug for delaying pterygium development.
Introduction
As a common ocular surface disease, pterygium is characterized by the proliferation and invasion of fibrovascular lesions into the cornea. 1 Later pterygium leads to a serious reduction in vision acuity, and surgical excision is the only treatment for pterygium. Although considerable progress has been made in etiology of pterygium, its pathogenesis and therapeutic strategies have not been fully clarified. 2 Threrfore, it is necessary to gain a deeper understanding of pterygium development.
Several studies have demonstrated that aberrant DNA methylation plays a crucial role in pterygium formation. Compared with normal conjunctiva, the transcription initiation of transglutaminase-2 and large tumor suppressor kinase 1/2 was significantly hypermethylated, whereas the metalloproteinase 2 and CD24 promoter genes were significantly hypomethylated in pterygium. 3 N6-methyladenosine (m6A) and methyltransferase 3 (METTL3), a key m6A methyltransferase, were downregulated in pterygium. 4
Histone lysine demethylase subfamily 4D (KDM4D), also known as JmjD2D, is highly conserved in placental mammals. KDM4D removes the methyl moieties from H3K9 and H1.4K26. 5 Many studies have proved that KDM4D upregulation is associated with tumorigenesis. For example, the overexpression of KDM4D promoted gastrointestinal stromal tumor progression, renal cancer angiogenesis, and acute myeloid leukemia development.6–8
As the main component of lyceum barbarum, Lycium barbarum polysaccharide (LBP) is a natural Traditional Chinese Medicine. LBP exhibits antifibrotic effects in treating corneal injury and does not irritate the eyes.9,10 As LBP can be administered topically, it is a potential ocular surface drug. 11
In this study, we investigate the role of KDM4D in pterygium formation and the effect of LBP on pterygium fibroblasts. We found that KDM4D expression was higher in pterygium and elevated KDM4D level promoted migration and inhibited apoptosis of pterygium fibroblasts. IL-1β, IL-6, and IL-8 enhanced the expression of KDM4D in pterygium fibroblasts. Moreover, the effects of KDM4D on pterygium fibroblasts were partially attenuated by LBP.
Methods
Patient samples preparation
Thirty-three pterygia were obtained from patients and informed consent was obtained. For immunohistochemistry, 29 pterygium samples served as the experimental tissue, and 14 residua conjunctivas from donors served as the control tissue. The excluded symptoms included other ocular infectious and noninfectious diseases, ocular trauma within the past 6 months, systemic immune diseases, corticosteroid use, and diabetes. For fibroblasts culture, the 4 pterygium samples were immediately digested and cultured. The remaining tissues were stored in liquid nitrogen until further use.
Immunohistochemistry
Samples were immediately immobilized in 4% formalin. Immunohistochemical staining was performed using Ventana BenchMark GX (Roche Diagnostics, Basel, Switzerland). In brief, antigen retrieval was performed using CC1 antigen retrieval solution (Roche Diagnostics) for 60 min. Specimens were incubated with anti-KDM4D/JMJD2D antibody (ab93694; Abcam) for 32 min, followed by visualization using the Ultra View Universal DAB Detection Kit (Roche Diagnostics) for 12 min. The sections were counterstained with hematoxylin, dehydrated, and covered with coverslips. The IHC results are shown as the percentage of KDM4D positive cells, and 600 cells were counted.
Pterygium fibroblasts culture
The experiment was performed as previously described. 12 In brief, the pterygium samples were cut into 1 mm3 pieces on a bacteria-free workbench, and digested with 0.25% trypsin-EDTA (25200-056; Gibco Life Technologies) at 37°C for 15 min. The digested solution was diluted with 1 mL culture medium.
Then the solution was filtered through 70 μm cell strainers (BD Falcon, Franklin Lakes, NJ) and centrifuged (200 g, 5 min) to obtain the cell sediment. Three milliliters DMEM/F-12 (C11330500BT; Gibco Life Technologies) supplemented with 10% fetal bovine serum (FBS, 10270-106; Gibco Life Technologies), 100 U/mL penicillin and streptomycin (15140-122; Gibco Life Technologies) was resuspended in the cell sediment and cultured (5% CO2, 37°C). The medium was changed every 2 days. Fibroblasts at passages 3 to 5 were used.
CCK-8 assay
Pterygium fibroblasts were seeded in a 96-well plate. When the cell density reached 70% confluency, the cells were treated with 100 ng/mL recombinant human KDM4D/JMJD2D protein (ab196398; Abcam) or 50 μg/mL LBP together in medium with 2% FBS for 24 h. Finally, 10 μL of CCK-8 reagent (CK04; Dojindo) was added and incubated for 2 h. A microplate reader (BioTek, Winooski, USA) was used to measure the absorbance value at a wavelength of 450 nm (OD450).
Quantitative real-time PCR
RNA extraction was performed using an RNA simple Total RNA Kit (DP419; TIANGEN, China). cDNA synthesis was performed using M-MLV reverse transcriptase (M1701; Promega, USA). The relative expression of KDM4D was detected by Quantitative PCR using TB Green Premix EX Taq II (RR820A; Taraka, Japan). GAPDH was used as an endogenous control.
Western blot
Pterygium fibroblasts were seeded in 6-well plates. When the cell density reached 70% confluence, the cells were treated with 10 ng/mL inflammation cytokines or LBP for 24 h. The cells were collected, lysed with RIPA buffer, 20 μg protein was electrophoresed by 12% SDS-polyacrylamide gels after mixing with loading buffer, and transferred onto polyvinylidene difluoride membranes. The membranes were blocked with milk and incubated with anti-KDM4D/JMJD2D antibody (ab93694; Abcam) and anti-β-actin antibody (ab8226; Abcam) at 4°C overnight, then incubated with secondary antibodies for 1 h at room temperature and stained with enhanced chemiluminescence (ECL) reagent β-actin was used as an endogenous control.
Liquid chromatography-mass spectrometry
Pterygium fibroblasts were seeded in 6-well plates. When the cell density reached 70% confluence, cells were treated with 100 ng/mL recombinant human KDM4D/JMJD2D protein for 24 h. The cells were lysed with M-PER Mammalian protein extraction reagent (78501; ThermoFisher). In total, 200 μg protein was added in 100 μL 50 mM NH4HCO3, then 10 mM DTT was added for 2.5 h at 37°C, alkylated with 10 mM iodoacetamide for 30 min at room temperature in the dark, centrifuged to obtain the proteins with Pierce™ Protein Concentrators PES (10K MWCO, 0.5 mL, 88513; ThermoFisher), then the proteins were digested with Pierce™ Trypsin Protease, MS Grade (90057; ThermoFisher) at enzyme–protein ratio 1:100, at 37°C overnight.
MS analyses were performed using a Q Exactive Plus MS system (Thermo Fisher Scientific). Raw MS data files were processed using Proteome Discoverer (Thermo Fisher Scientific). The analysis of differentially expressed genes (DEGs) and Gene Ontology enrichment was performed with LIMMA R package (P value <0.05, fold change >2).
Scratch wound healing assay
Pterygium fibroblasts were seeded in scratch culture inserts (80209; ibidi) in a 24-well plate. When the cell density reached 90% confluence, the scratch culture inserts were removed. The cells were treated with 100 ng/mL recombinant human KDM4D/JMJD2D protein (ab196398; Abcam) or 50 μg/mL LBP in a medium with 2% FBS. Wounds were photographed every 6 h from 0 to 48 h using a Cytation 5 cell imaging multimode reader (BioTek). The cellular migration area in each image was measured using Image J software. The area of the wound at 0 h was considered 100%.
Flow cytometry
For the apoptosis assay, pterygium fibroblasts were seeded in 6-well plates. When the cell density reached 80% confluence, cells were treated with 100 ng/mL recombinant human KDM4D/JMJD2D protein for 6 h, added 50 μg/mL LBP or not for another 6 h, then added 100 μM H2O2 for 1 h to induce cell apoptosis. Apoptosis was detected using an Annexin V, FITC apoptosis detection kit (AD10; Dojindo).
Statistical analysis
GraphPad Prism 8.0 was used for statistical analyses with unpaired 2-tailed Student's t-test. P < 0.05 was considered significant (*P < 0.05; **P < 0.01; ***P < 0.001).
Results
The expression of KDM4D in pterygium was higher than that in conjunctiva
We collected 29 pterygium and 14 conjunctiva samples and detected KDM4D expression using immunohistochemistry. Red arrows pointed KDM4D positive cells in the conjunctiva and pterygium (Fig. 1A). Further analysis revealed that the percentage of KDM4D positive cells in pterygium samples was significantly higher than that in conjunctiva samples (Fig. 1B).
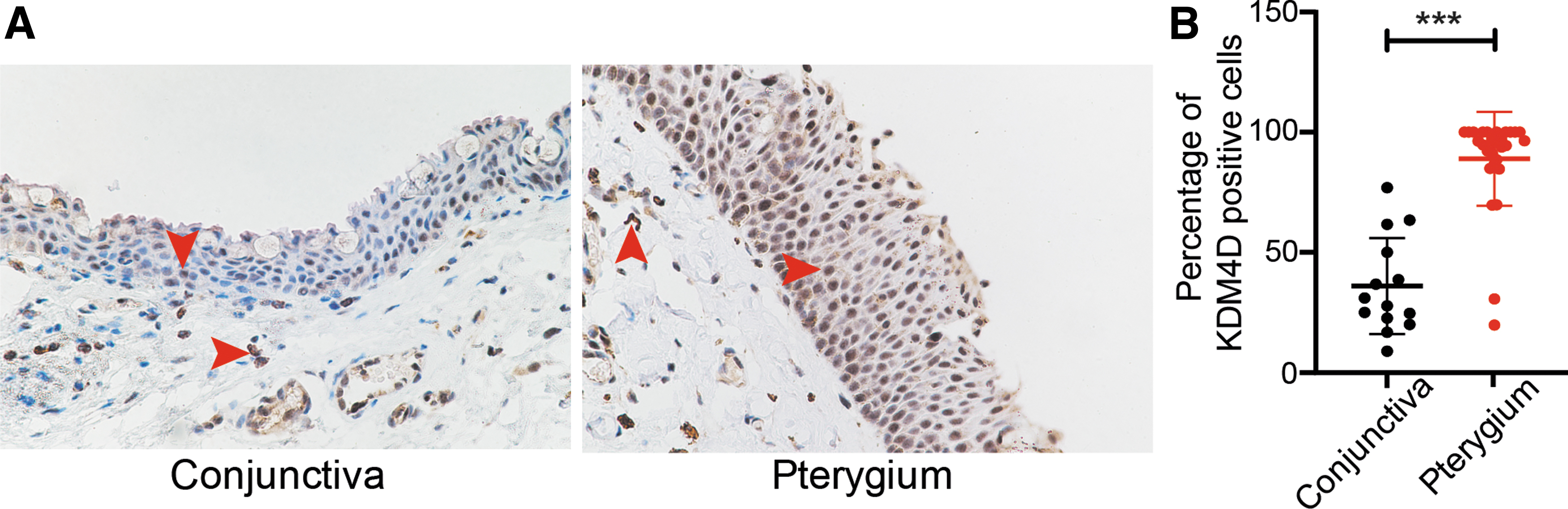
KDM4D expression in pterygium was higher than that in conjunctiva.
KDM4D promoted migration and inhibited apoptosis of pterygium fibroblasts
The proliferation and invasion of pterygium fibroblasts are the main mechanism of pterygium pathogenesis, 13 we then collected the pterygium samples and cultured the pterygium fibroblasts 12 to explore the effect of KDM4D on pterygium fibroblasts. Pterygium fibroblasts were treated with KDM4D (100 ng/mL) for 24 h. We analyzed the cell viability of pterygium fibroblasts affected by KDM4D using the CCK-8 assay. The results showed that the KDM4D group presented a higher OD450 value than the control group (Fig. 2A), indicating that KDM4D increased the viability of pterygium fibroblasts.
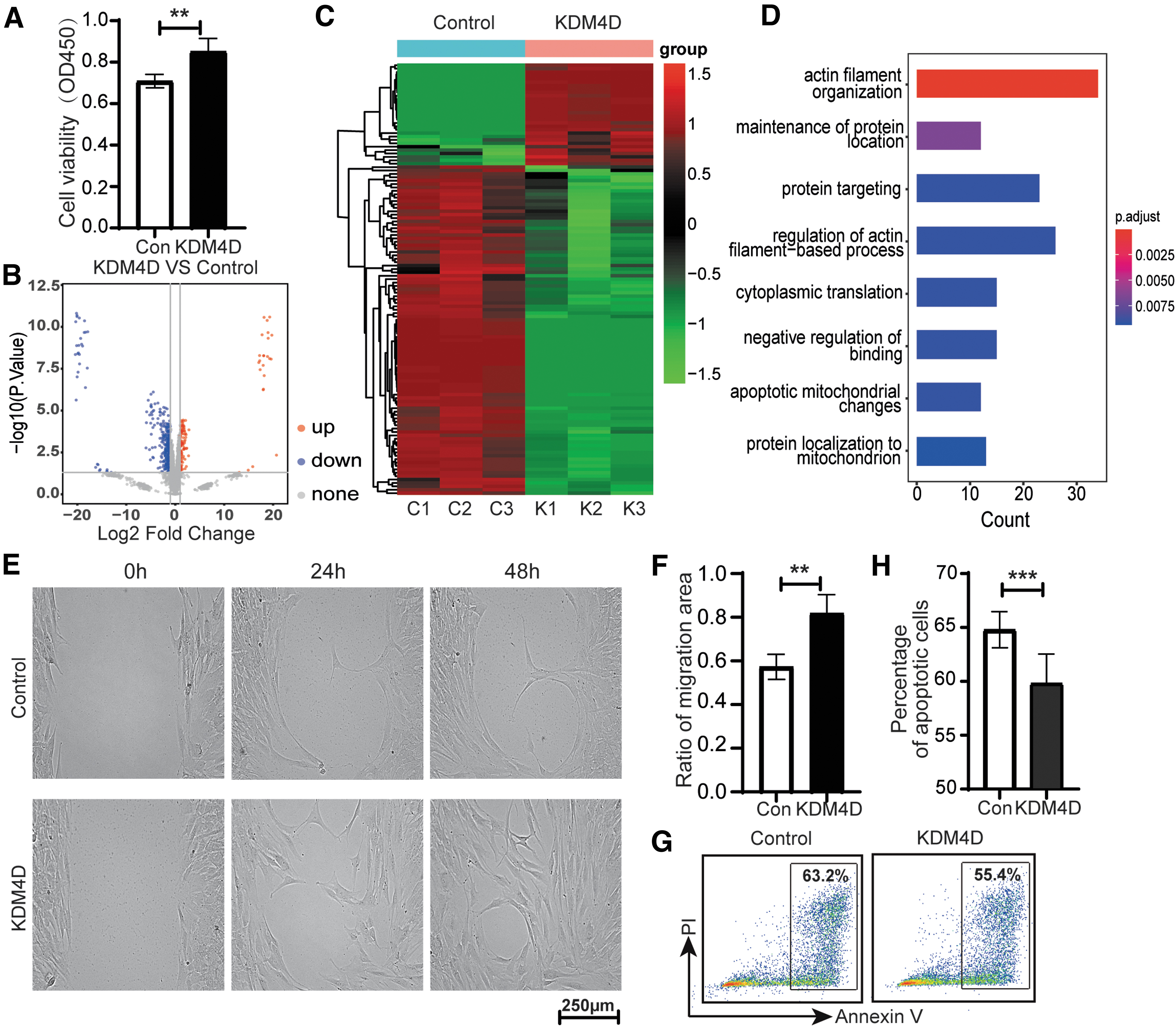
KDM4D promoted the migration and inhibited the apoptosis of pterygium fibroblasts.
To understand the mechanism, we analyzed the proteomics of pterygium fibroblasts after exposure to KDM4D by liquid chromatography-mass spectrometry assay. We identified a total of 4141 proteins. There were 112 upregulated and 341 downregulated proteins in pterygium fibroblasts after exposure to KDM4D (Fig. 2B, C). We then performed an enrichment analysis of the DEGs derived from the proteomics data. The results showed that the DEGs enriched in “actin filament organization” and “apoptosis” (Fig. 2D).
Since actin plays a key role in cell migration, 14 we first examined the migration of pterygium fibroblasts after exposure to KDM4D using a scratch assay. The results showed that the migration rate of pterygium fibroblasts was higher in the KDM4D group than in the control group (Fig. 2E, F). We examined apoptosis of pterygium fibroblasts after exposure to KDM4D using flow cytometry. We found that the percentage of apoptotic cells in pterygium fibroblasts was lower in the KDM4D group than in the control group (Fig. 2G, H). The results illustrated that KDM4D promoted the migration and inhibited the apoptosis of pterygium fibroblasts, in turn advancing the cell vigor of pterygium fibroblasts and promoting the progression of pterygium.
Inflammatory cytokines enhanced the expression of KDM4D in pterygium fibroblasts
Inflammatory factors can promote the disease progression by inducing KDM4D expression. 15 In pterygium, the genes associated with cytokine signaling in the immune system were upregulated compared with those in the conjunctiva (Fig. 3A, GSE183153 16 ). We then explored the relationship between the inflammatory cytokines and KDM4D expression in pterygium fibroblasts using IL-1β, IL-6, and IL-8 treatments. We found that the relative expression (including gene and protein expression) of KDM4D in pterygium fibroblasts was significantly increased after exposure to 10 ng/mL IL-1β, IL-6, or IL-8 for 24 h, respectively (Fig. 3B, C). These results confirmed that inflammatory cytokines enhanced the expression of KDM4D in pterygium fibroblasts.
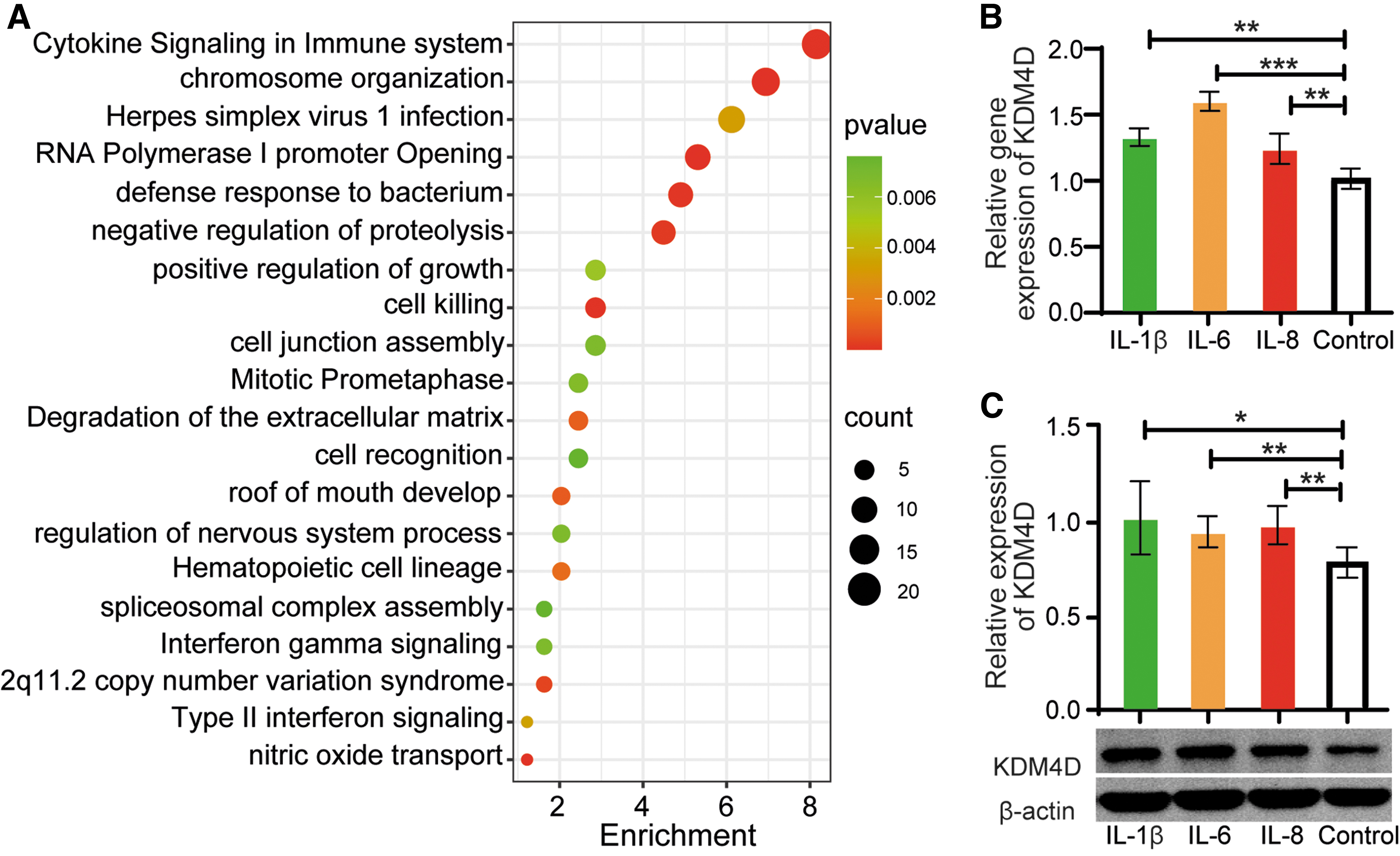
Inflammatory cytokines enhanced the expression of KDM4D in pterygium fibroblasts.
LBP inhibited the expression and attenuated the effect of KDM4D in pterygium fibroblasts
LBP is a natural Traditional Chinese Medicine that can reduce the inflammatory response in corneal disease. 11 So we speculated that LBP had an effect on the expression of KDM4D in pterygium. The results showed that the relative expression (including gene and protein expression) of KDM4D in pterygium fibroblasts was significantly decreased after exposure to 50 μg/mL LBP for 24 h (Fig. 4A, B). We then tested whether LBP plays a functional role in pterygium fibroblasts. We stimulated pterygium fibroblasts with LBP and KDM4D, and found that LBP reduced the viability of pterygium fibroblasts and partially attenuated the effect of KDM4D (Fig. 4C).
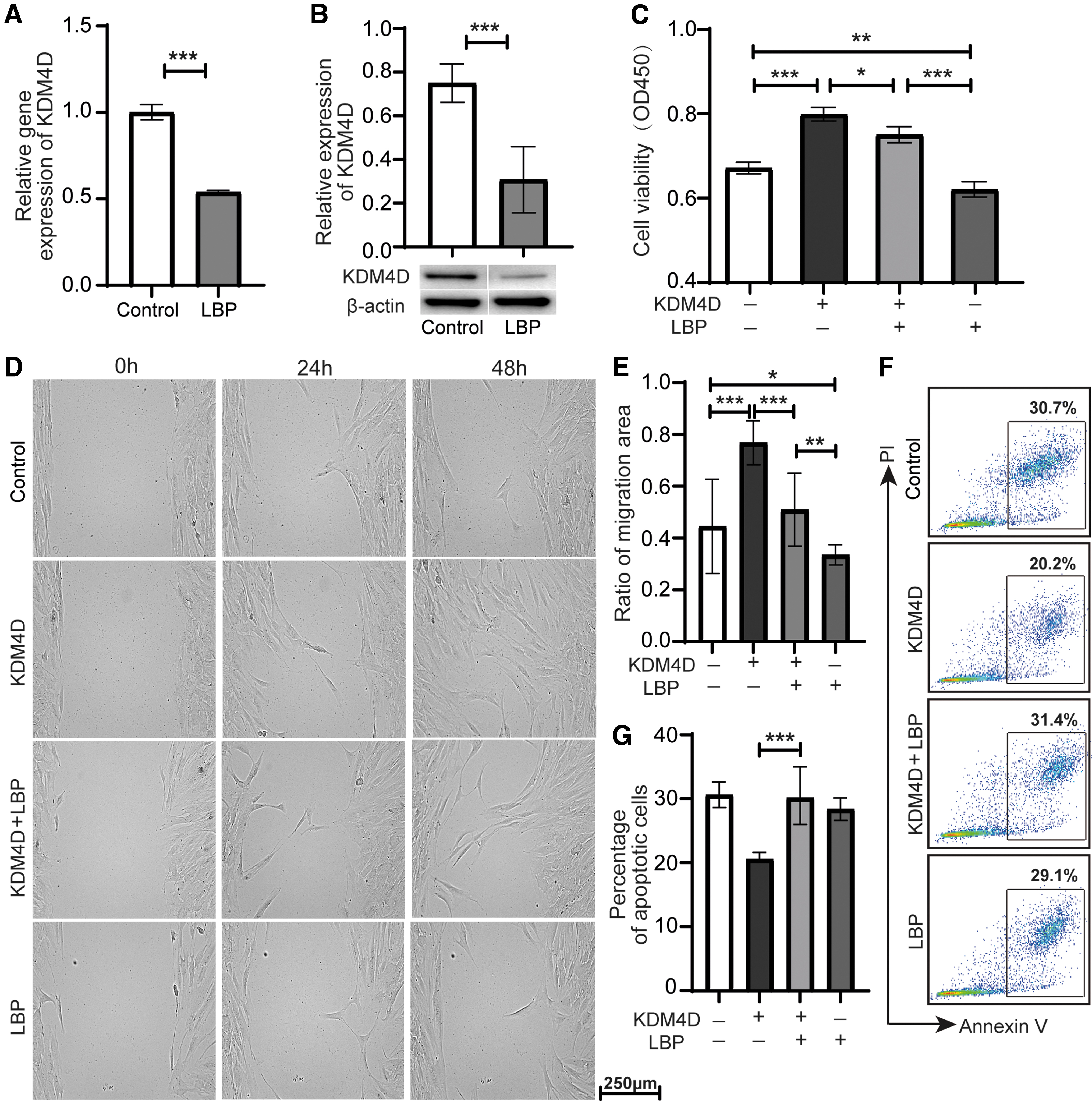
LBP inhibited the expression and attenuated the effect of KDM4D in pterygium fibroblasts.
Furthermore, we examined the effect of LBP on the migration and apoptosis of pterygium fibroblasts. The scratch wound assay showed that LBP inhibited the migration rate of pterygium fibroblasts and reduced the migration rate of pterygium fibroblasts induced by KDM4D (Fig. 4D, E). Moreover, LBP increased apoptosis of pterygium fibroblasts inhibited by KDM4D (Fig. 4F, G). These results indicated that LBP may be a potential drug for inhibiting pterygium development.
Discussion
Aberrant DNA methylation is associated with pterygium development. 17 Therapies targeting aberrant methylation are potential strategies for pterygium treatment. As a histone lysine demethylase, the high level of KDM4D may lead to aberrant DNA methylation. We found that the expression of KDM4D was higher in the pterygium than that in the conjunctiva. For privacy reasons, we did not include age and gender when processing the data. Inflammation of ocular surface plays a pivotal role in pterygium pathogenesis.
IL-1β, IL-6, and IL-8 are the major inflammatory cytokines in the tears from pterygium patients. 1 Our results showed that inflammatory cytokines, such as IL-1β, IL-6, and IL-8, enhanced the expression of KDM4D in pterygium fibroblasts. Therefore, we hypothesized that the high expression of KDM4D in pterygium is mainly the result of an abnormal ocular surface immune response, whereas age and gender are not the major factors.
KDM4D is a pro-proliferative molecule in many cancers.6–8 Overexpression of KDM4D in cardiac myocytes can increase the cell cycle by binding to the cell cycle gene promoter and increasing cell cycle gene expression. 18 Many studies have illustrated that the proliferation and invasion of pterygium fibroblasts are the main mechanisms of pterygium pathogenesis, and the recurrence of pterygium is associated with the fleshiness of stroma. 13 The proteomics assay showed that KDM4D is involved in the actin filament-associated process and apoptosis of pterygium fibroblasts. Actin plays a critical role in cell motility. 14 The scratch assay showed that KDM4D promoted the migration of pterygium fibroblasts. Thus, the effect of KDM4D on actin may play a key role in this process.
Moreover, the effect on apoptosis of KDM4D was further demonstrated by flow cytometry. Inducing apoptosis is an effective way to control disease progression. 19 Knockdown of KDM4D increases the expression of proapoptotic gene PUMA in liver cancer cells. 20 KDM4D may promote the viability of pterygium fibroblasts mainly by inhibiting apoptosis. We suggest that KDM4D is a candidate target for inhibiting the growth and recurrence of pterygium through the pathways associated with migration and apoptosis.
LBP inhibits the expression of profibrotic genes, including collagen, vimentin, and alpha-smooth muscle actin, in human corneal fibroblasts in vitro.9,10 Dry eye was ameliorated by topical application of LBP in mice. Moreover, LBP inhibited the infiltration of inflammatory cells and decreased the release of inflammatory cytokines in dry eye and vitiligo.11,21 Our results showed that LBP inhibited the viability and migration of pterygium fibroblasts, indicating that LBP could be a potential drug for treating pterygium. Remarkably, LBP itself had no effect on the apoptosis of pterygium fibroblasts, but it increased the apoptotic rate of pterygium fibroblasts inhibited by KDM4D. We speculate that there is an interaction between KDM4D and LBP and the molecular mechanism between KDM4D and LBP needs to be further studied.
The limitations of this study include the following aspects. First, the sample size is relatively small. Second, this study did not track the prognosis and recurrence of the patients. Thus, a prospective study with a larger sample size is warranted in the future. Nonetheless, our study provides evidence that elevated KDM4D expression in pterygium affects the migration and apoptosis of pterygium fibroblasts and that LBP partially attenuates the effect of KDM4D on pterygium fibroblasts. Therefore, the KDM4D expression should be concerned in pterygium and LBP plays a positive role in the inhibition of pterygium development.
Above all, overexpression of KDM4D in pterygium may be a risk factor for pterygium development, and LBP may be a potential drug to delay the development of pterygium.
Footnotes
Acknowledgments
The authors thank Dr. Lin Ma, Weiwei Li, Ying Li, Rui Wang, and Yi Dong for their help with the tissue collection.
Ethical Approval
The study was performed on the basis of the principles of the Declaration of Helsinki and approved by the ethics committee of Tianjin Eye Hospital.
Authors' Contributions
J.G. conceived and designed the study, performed the experiments, analyzed the data, and wrote the article. Y.W., R.H., J.L., P.H., and J.L. performed the experiments. X.C. performed the LM-MS assay. L.J., L.W., and Y.M. performed the Immunohistochemistry. L.C. and X.L. interpreted the results and oversaw the project.
Data access statement
All relevant data are within the article and its supporting information files.
Author Disclosure Statement
The authors declare that they have no affiliations with or involvement in any organization or entity with any financial interest in the subject matter or materials discussed in this article.
Funding information
This study was supported by the Tianjin Eye Hospital Youth Grant (YKQN2007, YKQN2005), the Tianjin Key Medical Discipline (Specialty) Construction Project (TJYXZDXK-016A), the Tianjin Health Research Project (TJWJ2023QN080), the Tianjin Technical attack project (15KG120), and the Beijing Bethune nonprofit foundation (BJ-GY2021019J).