
Abstract
Background:
One of the major risks of preterm birth is a history of conization. However, the risk of infection due to this procedure is still not well known. Using next-generation sequencing, we aimed to reveal the influence of conization on vaginal microbiota in the following pregnancy, and their relationship between spontaneous preterm birth (sPTB).
Methods:
We conducted a prospective cohort study including 133 pregnant patients, of whom 25 had conization histories and 108 did not. Vaginal microbiome samples were collected using swabs by an obstetrician upon inclusion in the first trimester and during delivery. V1–V2 of the 16S rRNA gene were amplified and analyzed to identify the bacteria.
Results:
The conization group had a significantly lower delivery week (34 weeks vs. 36 weeks, p = 0.003) and higher sPTB rate (64% vs. 8.3%, p ≤ 0.001) than the control group. In the conization group, alpha (Chao 1, p = 0.02; phylogenetic diversity whole tree, p = 0.04) and beta diversity (permutational multivariate analysis of variance test, p = 0.04) of the vaginal microbiota was significantly higher during delivery in patients who delivered preterm than in those who delivered term. Community-state type IV in the first trimester was significantly associated with sPTB (overall odds ratio 3.80, 95% confidence interval 1.33–10.8, p = 0.01).
Conclusions:
Conization is a risk factor for sPTB. Increased risk of sPTB in patients after conization may belong to the vulnerable defense mechanism, due to the shortened cervix and decreased cervical mucus.
Introduction
Microorganisms, known as vaginal microbiota, broadly colonize the vaginal tract. The spread of next-generation sequencing (NGS) revolutionized our method of detecting the microbiota in each organ and allowed us to elucidate its potential effect on health promotion and disease prevention. Although culture techniques focus on single and often well-characterized organisms, sequencing approaches allow for broad unbiased surveys of bacterial communities. 1
A healthy vaginal microbiota is considered to be dominated by Lactobacillus species to maintain a low vaginal pH by producing lactic acid. 2,3 Many classifications have been proposed using sequencing techniques considering vaginal microbiota, but the classification in most use nowadays is dividing them into five community-state types (CSTs). 4 CST I, II, III, and V are dominated by Lactobacillus, which are L. crispatus, L. gasseri, L. iners, and L. jensenii, respectively. CST IV consists of a higher proportion of bacteria that cause bacterial vaginosis (Gardnerella, Prevotella, Atopobium, Dialister, Megasphaera, and Ureaplasma) with lower proportions of Lactobacillus species.
Many of the CST IV cases are asymptomatic, although considered unhealthy. 5 However, L. iners is reported to have less power to sustain the vaginal microbiota than other Lactobacillus oligotypes due to less and different lactic acid isoform production. 6 Additionally, vaginal microbiota alters not only in individuals, but even in oneself in their life stage by numerous aspects (such as age, hormone levels, menstruation cycle, pregnancy, genital infection, sexual activities, and hygienic practices 6,7 ), implying the undetermined definition of “healthy” vaginal microbiota.
Gynecologic cancer has also been reported to negatively influence the vaginal microbiota. 8 Additionally, one of the known risks of adverse pregnancy outcomes is the history of conization to cervical intraepithelial neoplasia (CIN) or early-stage cervical cancer. Many females who undergo surgical intervention are of reproductive age and require special attention after pregnancy. Female with a history of conization for CIN has shorter cervical lengths during pregnancy. 9 Many studies have shown a significantly increased risk of preterm birth (PTB), perinatal morbidity, and mortality in patients with a history of conization. 10 –12
Berretta et al 13 described a higher risk of spontaneous preterm birth (sPTB) in cases with cervical resection of >1.5 cm. Firichenko et al 14 also reported an association between larger cervical conization size and a short cervix and higher sPTB risk, which was notably increased in nulliparous females. Sozen et al 15 reported that the importance of the volume of excised cervix tissue over height of the excised cervix tissue in perinatal complication assessments. They reported higher risks of sPTB and preterm premature rupture of the membranes in cases with excises cervical tissue volume excision of >2.27 and >3.99 cm3, respectively.
As previously mentioned, most of the previous literature refers to the risk of pregnancy after conization regarding mechanical cervical length shortening or cervical tissue reduction, 13 –15 and not about the potential fragility against bacterial infections.
We hypothesized that the decreased immune substance secreted from the cervical mucus due to loss of endocervical glands would lead to insufficient defense against infectious microorganisms ascending from the vagina, in addition to the physical impact of lost cervical tissues leading to cervical insufficiency. Currently, in the era of NGS, we can detect even a minute proportion of microorganisms that inhabit our body, while environmental microbiologists estimate that <2% of bacteria can be cultured in the laboratory. 16 This study aimed to clarify the vaginal microbiota of pregnant females with a conization history and its relationship with pregnancy outcomes using NGS.
Materials and Methods
Ethical statement
IRB registration number is 1182.
Study design
This prospective cohort study analyzed the vaginal microbiome of females with and without a history of conization. We included patients with singleton pregnancies who were referred to the University of the Ryukyus Hospital and Okinawa Prefectural Nanbu Medical Center and Children's Medical Center during the first trimester from April 2019 to November 2021. We excluded patients with multiple pregnancies, intrauterine fetal deaths, and fetal malformations, as well as patients with premature rupture of membrane before taking vaginal microbiome samples during delivery. Patients who agreed to participate in this study were managed similarly to other patients in each facility, except for the sample collection. Vaginal microbiome samples were collected using swabs by an obstetrician upon inclusion in the first trimester and during delivery. Cervical mucus elastase was collected in the first trimester with the vaginal microbiome swab. We evaluated the vaginal microbiome characteristics of patients with and without conization histories and their relationship with sPTB.
This study complied with the principles of the Declaration of Helsinki (revised in October 2013) and Ethical Guidelines for Medical and Health Research Involving Human Subjects. A written informed consent was obtained from all patients before inclusion. This study was approved by the institutional review board (IRB) of the University of Ryukyus Hospital, Okinawa Prefectural Nanbu Medical Center and Child Medical Center, and Varinos, Inc. (IRB No.1182).
Microbiome analysis
The samples were treated with proteinase K (Roche Applied Science, Penzberg, Germany) containing a 100 mg/mL lysozyme solution (Sigma–Aldrich, MO). Genomic DNA was extracted using MagNA Pure 24 (Roche Diagnostics, Grenzach-Wyhlen, Germany) Pathogen 1000 hp 3.1. Reagents and bacteria derived from the working environment greatly affect ultralow-biomass vaginal samples. Therefore, an experiment was conducted using ultrapure water as a negative control, and the bacteria derived from the reagents or the surrounding environment were monitored. The V1–V2 region of the 16S rRNA gene was amplified using universal primers (27Fmod and 338R).*
*Correction added March 3, 2023 after first online publication of January 31, 2023 to correct the text that was formerly stated as: “The V1–V2 region of the 16S rRNA gene was amplified using 27Fmod and 338R. 5′- The forward primer was CCATCTCATCCCTGCGTGTCTCCGACTCAGNNNNNNNNNNagrgtttgatymtggct 3′, cag- which is contained with the 454 primer A (indicated in capital letters), a 10 bp barcode sequence for each sample indicated in N), and 27Fmod (indicated in lowercase letters), 5′- and the reverse primer was CCTATCCCCTGTGTGC-3′, CTTGGCAGTCTCAGtgctgcctcccgtaggagt which is contained with the 454 primer B (indicated in capital letters) and 338R (indicated in lowercase letters).”
The final library was paired-end sequenced at 2 × 251 bp using a MiSeq Reagent Kit v3 on an Illumina MiSeq platform (Illumina, Inc., San Diego, CA). Operational taxonomic units (OTUs) were constructed after paired-end read quality filtering. OTUs were assigned to taxa by using a previously reported database. 17
Statistical analyses
All the analyses were conducted using R software version 4.1.3. Nonhierarchical clustering of microbiome profiles, samples, and negative controls was performed using weighted UniFrac distance. Samples clustered with the negative controls were not used in subsequent analyses. Hierarchical clustering with microbiome profiles, excluding background-contaminated bacteria, was conducted using the Bray–Curtis distance matrix, and heat maps were generated. Alpha diversity indexes were compared between the conization group and the control group after the rarefaction analysis, including Chao1 richness and phylogenetic diversity (PD) whole tree using microbiome profiles. Beta diversity was analyzed using principal coordinate analysis. Multivariate analysis based on weighted UniFrac distance was conducted to compare the differences in the microbiome between the two groups in the first trimester and during delivery. Beta diversity was analyzed using permutational multivariate analysis of variance (PERMANOVA) test.
We divided the bacterial communities into CSTs and calculated the odds ratio (OR) of sPTB in each CST.
Continuous data were analyzed using the t-test for normally distributed data and the Mann–Whitney test for nonnormally distributed data. Categorical variables were compared using Fisher's exact test. Statistical significance was set at p < 0.05.
The data sets generated during and/or analyzed during the current study are available from the corresponding author on reasonable request. After deidentification, individual participant data that underlie the results reported in this article, study protocol, and informed consent form are available. Data will be available immediately following publication, ending 5 years following publication, to anyone who wishes to access the data on any purpose. Request should be directed to the corresponding author. To gain access, data requestors will be required to sign a data access agreement presented by the corresponding author.
Results
Maternal characteristics and pregnancy outcomes
This study included 133 patients (25 patients in the conization group and 108 patients in the control group). The maternal characteristics and pregnancy outcomes are listed in Table 1. The mean maternal ages were 35 and 34 years in the conization and control groups, respectively (p = 0.37). All participants were Asians. No significant differences were found in parity (p = 0.95), abortion history (44% vs. 36.1%, p = 0.07), PTB history (68% vs. 16.8%, p = 0.08), body mass index before pregnancy (25.2 vs. 25.45, p = 0.13), cervical cerclage (8% vs. 2.8%, p = 0.94), smoking history (56% vs. 78.7%, p = 0.41), or use of immunosuppressant medications (4% vs. 2.8%, p = 0.75). The mean delivery week was 36 weeks (range: 27–41 weeks) in the conization group and 38 weeks (range: 20–41 weeks) in the control group, which was significantly different (p = 0.003). The sPTB rates in the conization and control groups were 64% and 8.3%, respectively (p ≤ 0.001, OR: 6.1875; 95% confidence interval [CI]: 2.13–17.93).
Maternal Characteristics and Pregnancy Outcomes of the Study Population
Data are presented as median (range) or n (%).
BMI, body mass index; CI, confidence interval; OR, odds ratio.
Alpha and beta diversity analysis
As there were no samples classified in the same cluster as the negative control, we analyzed every sample taken. No significant differences were found in alpha (Fig. 1a, b) and beta diversities (Fig. 2a, b) of the vaginal microbiota between the conization and control groups, with respect to timing. No significant differences were found in alpha (Fig. 1c, d) and beta diversities (Fig. 2c, d) between term births and sPTB cases in the control group at either time point. However, there were no significant differences in alpha (Fig. 1e) or beta diversities (Fig. 2e) between term birth and sPTB cases in the first trimester of the conization group, significant differences were found in both alpha (Fig. 1f, Chao 1 p = 0.02, PD whole tree p = 0.04) and beta diversity (Fig. 2f, PERMANOVA test, p = 0.04) at the time of delivery in the conization group.

Alpha diversity in the first trimester and at delivery. Alpha diversity box plots of vaginal microbiota at the first trimester and at delivery. The y-axes indicate the values of bacterial richness as the Chao index and PD whole tree. p-values were calculated by the Mann–Whitney tests.

Beta diversity in the first trimester and at delivery. Beta diversity plots of vaginal microbiota at the first trimester and at delivery. PCoA plot based on unweighted Unifrac distance. Each point corresponds to an individual sample. PERMANOVA test was used for unweighted UniFrac distances to beta diversity comparisons.
CST and its relation with conization and sPTB
A heat map of the relative abundance of the major oligotypes in the vaginal community, divided by the history of conization and timing of birth, is presented in Figure 3.
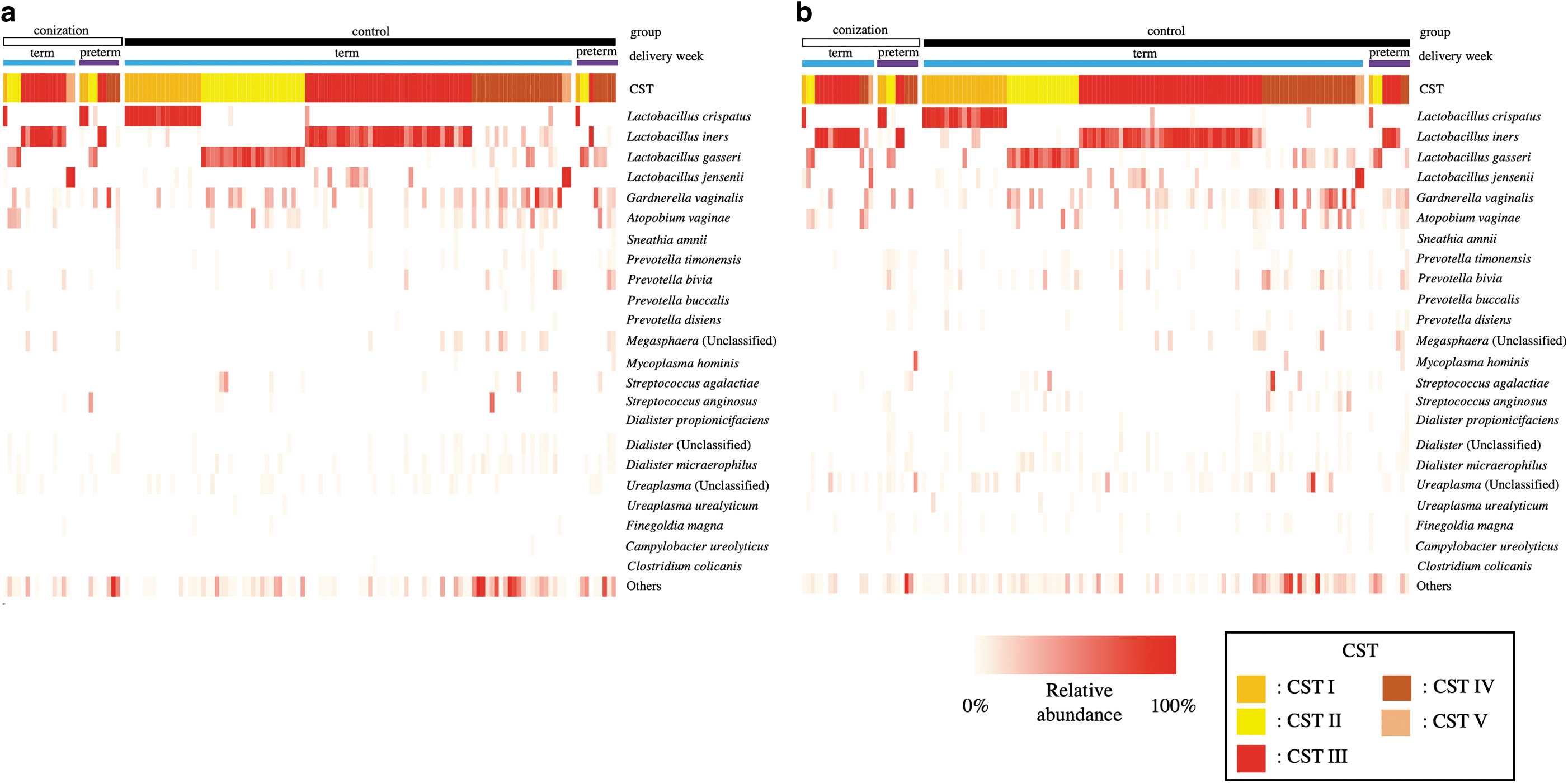
Heat map of percentage abundance of microbial taxa found in the vaginal microbial communities of 133 pregnant women. The upper color bar shows the group (conization or control), the middle color bar shows the delivery weeks (term or preterm), and the lower color bar shows the five CST of each sample.
The CST proportion in each group in the first trimester and the time of delivery are shown in Figure 4a and b. In the control group, CST III increased from 35.1% to 41.6%, CST II decreased from 23.1% to 16.6%, and CST IV slightly decreased from 23.1% to 21.2% from the first trimester to the time of delivery, respectively. In the conization group, CST IV increased from 12% to 20%, whereas CST II and V decreased from 20% to 16% and from 8% to 4%, respectively, from the first trimester to the time of delivery.
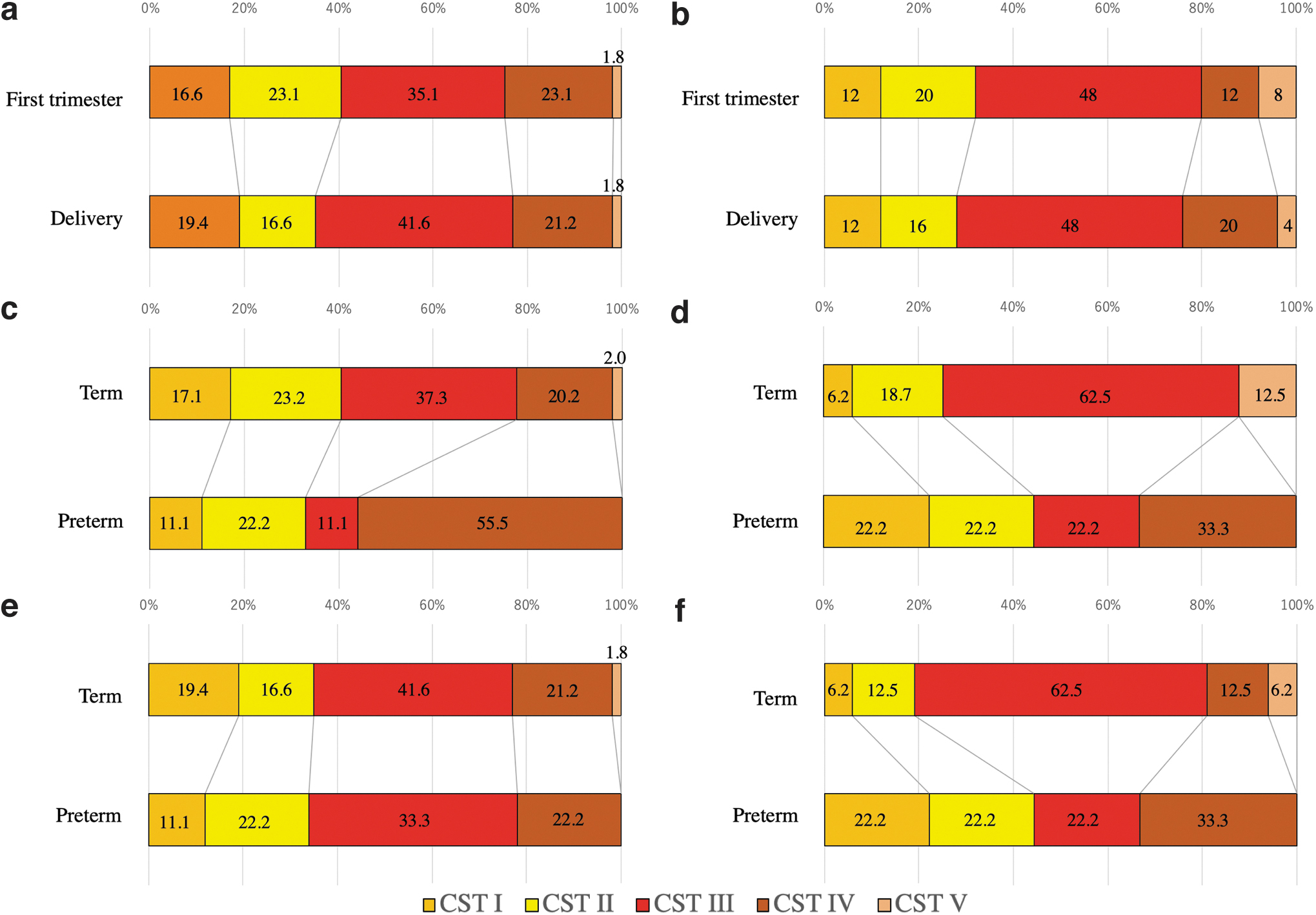
The proportion of each CST in each group (number: %).
The CST proportion in the first trimester of the control and conization groups is shown in Figure 4c and d, respectively. CST IV was greater in sPTB cases than in term cases in both the groups.
The CST of the control and conization groups at the time of delivery is shown in Figure 4e and f, respectively.
Because CST IV tended to increase in sPTB cases compared with term cases in both groups, as shown in Figure 4, we calculated the sPTB rate of each CST in both groups. Figure 5 shows the sPTB rate for each CST. CST IV showed the highest sPTB rate (28.5%) in the first trimester. In the first trimester, CST III had a significantly lower sPTB rate than CST IV, whereas others did not have a significant difference (Fisher's exact test; CST IV vs. CST V, p = 0.54; CST IV vs. CST III, p = 0.01; CST IV vs. CST II, p = 0.20; CST IV vs. CST I, p = 0.30). CST II had the highest sPTB rate at the time of delivery. No specific CST was related to sPTB at the time of delivery.

Preterm delivery rate of each CST.
The OR of sPTB in each CST is shown in Table 2. In the first trimester, CST IV was significantly associated with sPTB (overall OR 3.80, 95% CI 1.33–10.8, p = 0.01). Although not significant, CST III tended to reduce the sPTB rate in the conization group (OR 0.17, 95% CI 0.02–1.11, p = 0.06).
Odds Ratio of Preterm Birth in Each Community-State Types
Data are presented as OR (95% CI, p-value). Significance was determined by Fisher's exact test.
OR cannot be calculated because all the cases were spontaneous preterm birth.
CST, community-state types.
Discussion
This prospective cohort study analyzed the vaginal microbiota of pregnant females with and without conization history and their relationship with sPTB. This study revealed that patients with conization histories who delivered at term had a significantly less diversified vaginal microbiota during delivery.
Initially, the sPTB rate was significantly higher, and the mean delivery week was significantly lower in the conization group, which is consistent with previous reports. 10 –12 In patients with a history of conization, sPTB is said to be caused by cervical insufficiency due to shortened cervix, 13 –15 and few studies mention the possible fragility against infectious bacteria ascending from the vagina due to deteriorated defense mechanisms. Recent studies suggest that increased vaginal microbiota diversity, combined with the reduced relative abundance of Lactobacillus species, is involved in human papillomavirus acquisition, persistence, and development of CIN and cancer. 18,19
Zhang et al 20 reported a decrease in vaginal microbiota diversity 3 months after the loop electrosurgical procedure (LEEP). They revealed a decreased Prevotella-containing microbiota, a lack of consistent dominant species, and an increased abundance of L. iners. Similarly, Wiik et al 21 reported that the vaginal microbiota of patients after LEEP due to CIN had reduced non-Lactobacillus bacterial species at 6 and 12 months after the procedure. This study aimed to clarify the differences in the vaginal microbiota during pregnancy after conization. We found no significant differences in the alpha and beta diversities of the vaginal microbiota between patients with and without conization histories in the first trimester or before delivery. This may be because vaginal dysbiosis was improved by conization, as described above, and showed no difference compared with the control group.
Conversely, we found that patients with a conization history and who delivered at term had significantly lower alpha and beta diversities in their vaginal microbiota at the time of delivery. This finding suggests that a more stable or less diversified vaginal microbiota is necessary for patients who underwent conization to deliver at term. Since neither alpha/beta diversity nor the proportion of CST IV increased in patients with conization history, conization itself does not have a negative influence against vaginal microbiota, as previously reported. 21 However, the classification of CST only reflects the abundance of Lactobacillus species and does not take the diversity of other bacteria into account.
This means that there are cases with high alpha and beta diversity but are classified in CST I, II, III, or V. The fragility against the diversified infectious vaginal bacteria, which may only account for a small abundance in the whole vaginal microbiota and may be classified as CST I, II, III, or V, could be the reason for our results, which shows the increased sPTB rate and the need of less diversified vaginal microbiota to deliver in term in patients after conization. This may be due to a decreased amount of cervical tissue and cervical mucus that makes the patients after conization vulnerable to the even minute proportion of infectious bacteria, which inhabits in the vagina. The influence of decreased cervical mucus following cervical excision has been a concern in fertility and pregnancy in the past. 22,23
Many obstetricians believe that sPTB in patients with a conization history is a result of cervical insufficiency. 24,25 However, if cervical insufficiency is the main reason for sPTB in patients with a conization history, cervical cerclage will likely extend the gestational age; however, this has been controversial because of conflicting results in numerous reports. 26,27 This may be because the vulnerability to infectious bacteria due to decreased cervical tissues and mucus accounts more for sPTB than mechanical cervical weakness. Many studies have reported the relationship between foreign material used in cervical cerclage, 28,29 but only a few have reported that vulnerability to bacteria causes sPTB. 30,31 The findings of our study support this hypothesis. Although not statistically significant, patients with a history of conization tended to have a higher cervical mucus elastase value in the first trimester, which also suggests the influence of infection.
Many studies in the era of culture have shown a correlation between vaginal infections and sPTB. 32,33 In the era of NGS, the correlation between infection and its negative influence has been shown in more detailed conditions. A low abundance of Lactobacillus is associated with sPTB. 34,35 Furthermore, we can now subdivide Lactobacillus species and recognize their ability to stabilize the vaginal microbiota. Vaginal microbiota dominated by L. crispatus, L. jensenii, and L. gasseri have been reported to decrease the risk of sPTB. 36 Our study revealed that CST IV has a significant relationship with sPTB in the first trimester with an OR of 3.80, which is consistent with previous reports. 34,35 This relation was seen in both groups.
Conversely, CST III tended to reduce sPTB in our study. L. iners is said to produce less lactic acid and possesses weak power to stabilize the vaginal microbiota, 6 and some CST III cases in the first trimester did change to CST IV before delivery. However, its influence on increasing sPTB may be minimal, considering that a large proportion of past studies, including this study, showed no difference in the sPTB rate in CST III cases. 37,38 CST I in the first trimester of conization group showed a relatively high OR without significance. Two out of three cases resulted in sPTB in this group, accounting for the high OR. CST I is reported to reduce sPTB, 36 and we could not find any other risk factor of sPTB that these three cases share. We assume that the high proportion of sPTB in CST I cases in the conization group is due to the small sample size, resulting in such deviation. More samples are needed to clarify this result.
To the best of our knowledge, this study is the first study to examine the vaginal microbiota of pregnant women after conization using NGS. However, this study has several limitations. First, the overall sample size was relatively large compared with previous studies, but it was still underpowered to detect the differences in diversities in other data sets, considering the event rate of sPTB. Second, we were not able to control the risk factors for sPTB other than the history of conization before inclusion, due to the expected small sample size. However, known major risks of sPTB did not have a significant difference as shown above. Finally, all participants were Asians, which disabled its generalization worldwide since the study was conducted in one region of Japan. However, previous literature has revealed that the vaginal microbiota of Japanese females resembles that of other racial groups, 39 which is favorable for generalization to different regions and races. Further accumulation of data from various races is required to overcome these limitations.
Conclusions
This study revealed that the risk of sPTB was higher in patients with a history of conization than in those without. CST IV in the first trimester was associated with sPTB in both groups. There was no increase in CST IV cases or microbiota diversity between the patients with and without conization. However, the alpha and beta diversities of the vaginal microbiota were significantly lower in patients with a conization history who delivered at term. These data imply that the increased risk of sPTB in patients after conization belongs to the vulnerable defense mechanism against the diversified vaginal bacteria that may only account for a small part of the whole vaginal microbiota and may be classified as CST I, II, III, or V. This is possibly due to the shortened cervix and decreased cervical mucus after conization. Further accumulation of data in various races is needed to elucidate the exact mechanism by which the risk of sPTB in patients with conization histories is increased and to provide the best treatment to improve pregnancy outcomes.
Footnotes
Acknowledgment
The authors would like to thank Enago for the English language review.
Authors' Contributions
The work presented herein was carried out in collaboration among all authors. S.H.: Formal analysis (equal), investigation (lead), writing—original draft (lead). T.K.: Conceptualization (lead), investigation (support), project administration (lead), writing—review and editing (equal). O.Y., N.Y., Y.K., Y.C., and Y.N.: Investigation (supporting), writing—review and editing (equal). K.A.: Data curation (lead). S.E.T., W.A., M.S., and Y.S.: Formal analysis (equal), resources (equal), writing—review and editing (equal). K.M.: Writing—review and editing (equal). Y.A.: Conceptualization (supporting), project administration (supporting), supervision (lead), writing—review and editing (equal). All authors have read the article and have approved this submission.
Authors' Disclosure Statement
S.E.T, W.A., M.S., and K.A. are employees of Varinos. Y.S. is the CEO and is the shareholder of Varinos. S.H., T.K., O.Y., N.Y., Y.K., Y.C., Y.N., K.M., and Y.A. have no conflict of interest to disclose.
Funding Information
No funding was received for this article.