
Abstract
Abstract
Background:
Although it is generally accepted that exercise accelerates lymph flow, no study has directly measured lymph flow as a function of exercise intensity. In this study, we have measured flow in the thoracic lymph duct of five instrumented dogs while they ran on a treadmill.
Methods and Results:
Dogs were surgically instrumented with an ultrasonic flow transducer on the thoracic lymph duct and a catheter in the descending thoracic aorta. After recovery from surgery, the dogs ran on a treadmill at speeds which varied stepwise from 0 to 10 mph and from 10 to 0 mph. Dogs ran for 1 min at each speed with 15 min rest between each exercise. Heart rate increased significantly during exercise, whereas mean aortic pressure did not change. Resting lymph flow was 1.7 ± 0.2 ml/min. Exercise at 1.5 mph significantly increased lymph flow to 3.9 ± 0.6 ml/min (P < 0.01), 121% higher than resting flow. Lymph flow was further elevated at higher treadmill speeds, reaching 9.0 ± 1.6 ml/min (P < 0.01) at 10 mph, 419% higher than resting flow. Regression analysis demonstrated a linear relationship between treadmill speed and the percent increase in lymph flow. Lymph flow returned to the resting rate 1-2 min post-exercise.
Conclusion:
Lymph flow in the thoracic duct is positively correlated with exercise intensity.
Introduction
Methods
Subjects
This study was approved by the Institutional Animal Care and Use Committee and was conducted in accordance with the Guide for the Care and Use of Laboratory Animals. Five adult mongrel dogs, 3 males and 2 females, free of clinically evident disease were used for this study. The average weight of the dogs was 30.2 ± 2.5 kg.
Surgical preparation and instrumentation
The day before surgery, the dog's neck was shaved, and a fentanyl (1.5–3 μg/kg) patch was applied. The dog was fasted overnight. On the day of surgery, a preanesthetic, acepromazine maleate (0.3 mg), was injected subcutaneously. Anesthesia was induced 30 min later with thiopental sodium (5 mg/kg) administered through an intravenous line inserted into an antecubital vein. Endotracheal intubation was followed by mechanical ventilation with isoflurane (1%–3%) and oxygen to maintain a surgical plane of anesthesia.
A left lateral thoracotomy was performed in the fourth intercostal space using sterile technique. A pressure monitoring catheter (model LAP17, 17ga; Edward Lifesciences, Irvine, CA) was introduced in the descending aorta. This catheter was connected to a pressure transducer, and arterial blood pressure was monitored (Hewlett–Packard Pressure Monitor, 78354A) for the remainder of the operation. Blood samples were periodically collected from the aortic catheter for assessment of arterial blood gases and pH (GEM Premier 3000 Blood Gas/Electrolyte Analyzer, Model 5700, Instrumentation Laboratory, Lexington, MA) during surgery. An ultrasonic flow transducer (model 2SB/2.5SB; Transonic Systems Inc., Ithaca, NY) was implanted on the thoracic lymph duct and secured to the surrounding tissue with sutures.
The aortic catheter and the flow transducer cable were passed through subcutaneous tunnels and exteriorized between the scapulae. A chest tube was placed to evacuate the pneumothorax; this tube was removed on the second postoperative day. Ribs were approximated and the chest was closed in layers. Bupivacaine was infiltrated around the incision, and buprenorphine was injected to alleviate postoperative pain. Wounds were treated with Triple Antibiotic Cream (Bacitracin, Neomycin & Polymixin B) to combat infection. Antibiotics (cephalosporin, SMZ/TMP, fluoroquinolones) were given postoperatively for 10 days to prevent infection. The aortic catheter and the flow transducer cable were wrapped in gauze, and the chest of the dog was wrapped with veterinary wrap bandage, which also covered the catheter and transducer cable. This bandage was covered with a nylon canine jacket (Alice King Chatham, Medical Arts, Hawthorne, CA). The aortic catheter was flushed daily with heparinized saline (1000 U/ml) to prevent clotting.
Experimental protocols and data collection
After 10–12 days of recovery from surgery, data were collected with the dog standing on a canine treadmill, and while the dog ran at specified speeds on a treadmill at 0% grade. At rest and during exercise, aortic blood pressure and thoracic duct lymph flow were measured continuously. The aortic catheter was attached to a pressure transducer (ISOTECTM Disposable pressure transducer, Quest Medical Inc, Allen, Texas) positioned at the mid-chest level, and the flow transducer was connected to a flowmeter (T206, Small Animal Blood Flowmeter, Transonic Systems Inc.). Pressure and flow data were recorded on a multichannel chart recorder (Grass Model 7D Polygraph, Quincy, MA) and on a data acquisition system (EMKA 2.2.17.19, EMKA Technologies USA, Falls Church, VA). The data acquisition system computed mean aortic blood pressure and heart rate from the aortic pressure signal and mean lymph flow for the resting baseline condition and for each period of exercise and for postexercise periods.
Two experimental protocols were used. In one protocol, the speed of the treadmill increased from 0 to 1.5, 3, 4, 5, 6, 7, 8, 9, and 10 mph. The dog ran for 1 min at each speed, and rested for 15 min between each run. The other protocol was similar, except that the initial treadmill speed was 10 mph, and then decreased to 9, 8, 7, 6, 5, 4, 3, and 1.5 mph. These protocols were repeated at least once, except one dog did not run at 10 mph, and the descending speed protocol was not performed on this dog. For the other dogs, there were no significant differences between lymph flow values at respective treadmill speeds whether the exercise began at 1.5 mph or at 10 mph. Thus, for each dog, data from corresponding treadmill speeds were averaged.
At the conclusion of this study, the dogs were assigned to another research project.
Statistical analysis
Data are presented as mean ± standard error. Data describing hemodynamic variables (Table 1) and lymph flow as a function of treadmill speed (see Fig. 1) were analyzed with a repeated measures, one factor analysis of variance (ANOVA). Data used to compare responses to treadmill speeds of 3 and 9 mph at varying times (see Fig. 3) were analyzed with a repeated measures, two factor ANOVA. Differences among means were considered significant if P < 0.05. In that case, a Student–Newman–Keuls multiple comparison test was performed to identify specific differences among means. Linear regression was used to examine relationships between heart rate and treadmill speed and between percent change in thoracic duct lymph flow and treadmill speed (see Fig. 2). All statistical analyses were performed by GB-stat statistical software, version 10.

Lymph flow in the thoracic duct is plotted at rest and during exercise at treadmill speeds from 1.5 to 10 mph (n = 5). Dogs exercised for 1 min at each speed with 15 min of rest between each exercise. Values are mean ± SE, plotted at the midpoint of each data collection interval. **P < 0.01 vs. rest.
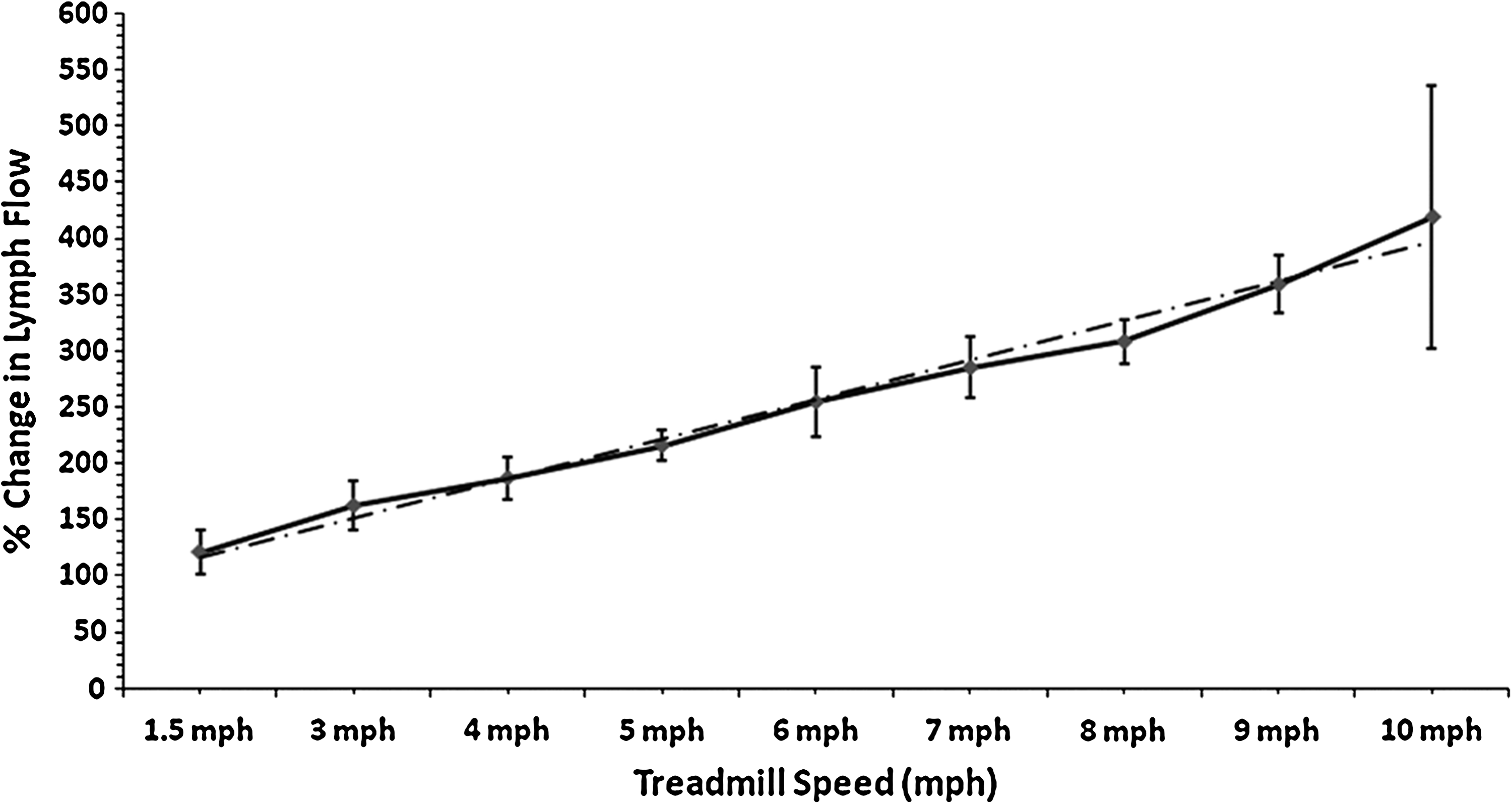
Percent change in thoracic duct lymph flow relative to flow at rest is plotted for treadmill speeds from 1.5 to 10 mph (n = 5). Values are mean ± SE, plotted at the midpoint of each data collection interval. The interrupted line demonstrates a positive linear relationship between percent change in lymph flow and treadmill speed (P < 0.001): Percent change in lymph flow (%) = (56.8 ± 11.3 %) + (33.7 ± 1.7%/mph) × treadmill speed (mph).
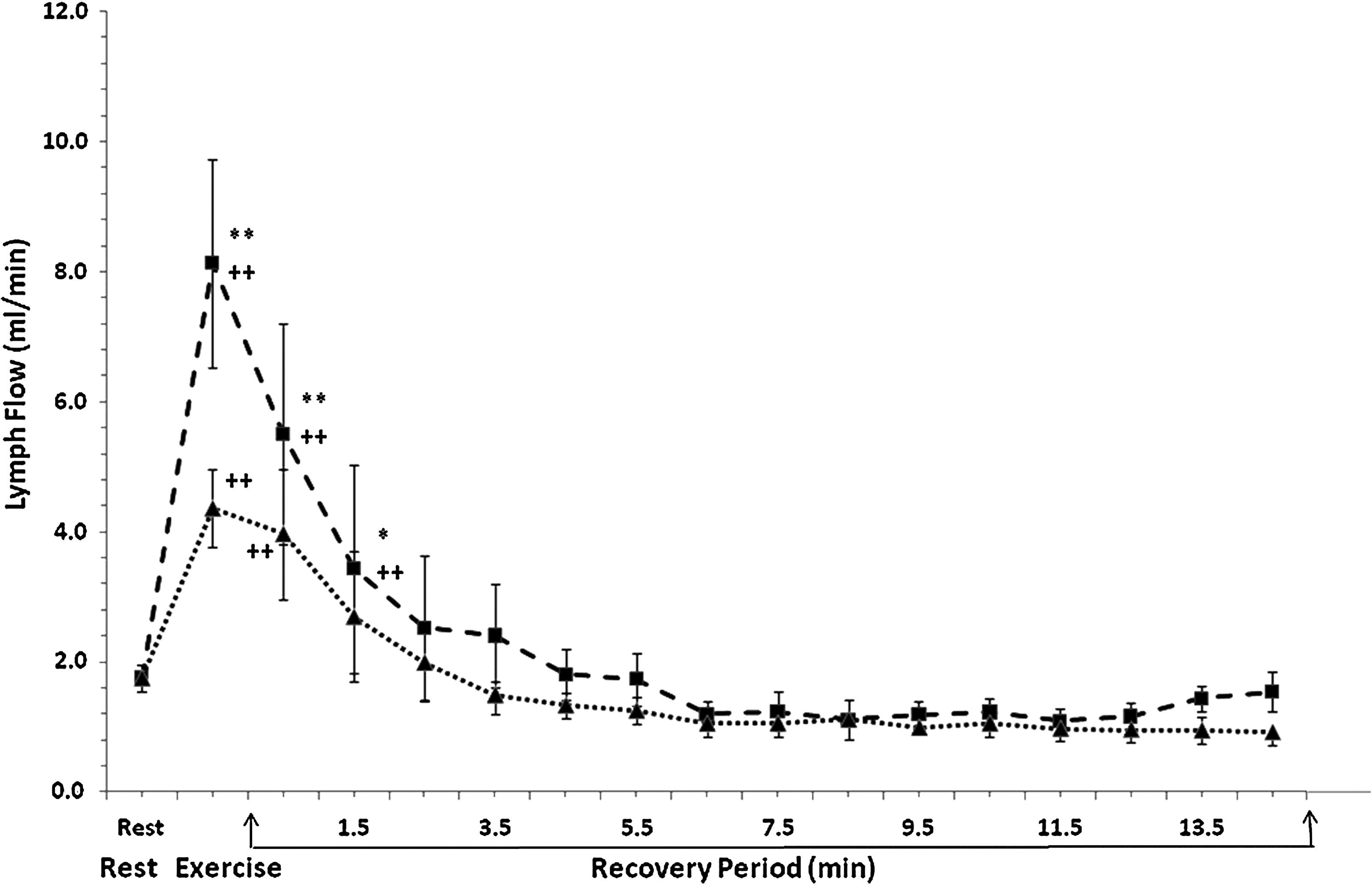
Lymph flow in the thoracic duct is plotted for the resting condition, 1-min exercise, and during 15 min postexercise at treadmill speeds 3 mph (n = 4) and 9 mph (n = 4). Values are mean ± SE, plotted at the midpoint of each data collection interval. **P < 0.01 vs. treadmill speed @ 3 mph, *P < 0.05 vs. treadmill speed @ 3 mph, ++P < 0.01 vs. rest for respective speed.
Dogs (n = 5) exercised for 1 min at each speed with 15 min rest between each exercise.
Values are mean ± SE. *P < 0.05 vs. rest, **P < 0.01 vs. rest.
Results
Effect of exercise on hemodynamic variables
Hemodynamic data for the resting, baseline condition and for 1-min exercise periods at increasing treadmill speeds are presented as mean ± SE in Table 1. At rest, heart rate was 127 ± 7 beats per min (bpm). Heart rate increased to 159 ± 5 bpm when the dog ran at 1.5 mph (P < 0.05), and the heart rate continued to increase reaching 255 ± 36 bpm at a speed of 10 mph (P < 0.01, compared to rest). The relationship between heart rate and treadmill speed was assessed by linear regression, which produced the equation: Heart Rate (bpm) = (139 ± 4 bpm) + (11.0 ± 0.6 bpm/mph) ×Treadmill Speed (mph) (P < 0.0001). Mean aortic pressure for baseline condition was 120 ± 8 mmHg. Mean aortic pressure did not significantly change at any treadmill speed.
Effect of exercise on lymph flow in the thoracic duct
Figure 1 presents mean values of thoracic duct lymph flow at the resting baseline condition and for each 1-min exercise period at increasing treadmill speeds. Figure 2 shows percent changes in the thoracic duct flow during exercise compared to flow at rest. Baseline thoracic duct lymph flow was 1.7 ± 0.2 ml/min. Exercise at 1.5 mph significantly raised the lymph flow to 3.9 ± 0.6 ml/min (P < 0.01), which was 121% higher than resting flow. Lymph flow further increased as treadmill speed was increased, reaching 9.0 ±1.6 ml/min (P < 0.01) at a speed of 10 mph (419% higher than resting flow).
The relationship between percent change in lymph flow and treadmill speed (Fig. 2) was analyzed by linear regression, which yielded the following equation: Percent Change in Lymph Flow (%) = (56.8 ± 11.3 %) + (33.7 ± 1.7 %/mph) ×Treadmill Speed (mph). Regression analysis demonstrated a positive relationship between treadmill speed and thoracic lymph flow (P < 0.0001).
Thoracic duct lymph flow during the 15-min recovery period after exercise was studied in detail for four dogs. In Figure 3, lymph flow is plotted as a function of time for A) the resting condition, B) 1 min treadmill exercise, and C) 1-min intervals during 15 min postexercise. This figure shows lymph flow data collected before, during, and after exercise at 3 mph (dotted line) and 9 mph (dashed line). Consistent with Figures 1 and 2, lymph flow was greater during and immediately following exercise at 9 mph compared to exercise at 3 mph. For exercise at 3 mph, lymph flow was significantly elevated at 1 min postexercise and returned to the resting value by 2 min postexercise. For exercise at 9 mph, lymph flow remained elevated for 2 min postexercise.
Discussion
While it is generally accepted that contraction of skeletal muscles compresses lymph vessels and accelerates lymph flow, 1 relatively little research has been reported on this important topic. 5 In 1953, Elkins et al. chronically catheterized the thoracic lymph duct of dogs, and then measured lymph flow by timed collection of lymph continuously diverted from the duct. For one dog, they reported lymph flows of 0.3 ml/min at rest and 0.8 ml/min during walking. 2 In 1977, Schad and Brechtelsbauer implanted an extracorporeal shunt between the thoracic lymph duct and the superior vena cava of splenectomized, heparinized dogs. 3 With this shunt, thoracic duct flow could be measured by intermittently diverting lymph from the shunt and then re-infusing the lymph. When these dogs walked at approximately 6 km/h (∼3.7 mph), lymph flow increased by about 100%. Also using an extracorporeal shunt, Lindena et al. reported that in dogs performing treadmill exercise, also at 6 km/h, thoracic duct lymph flow increased to 4.9 ml/min. 4 Several studies, using indirect methods, have demonstrated also that exercise increases regional lymph flow in exercising muscles,6,7 finger webs, 8 axilla and forearm, 9 the hind limb, 10 and lungs10–12 of animals and/or humans. However, only recently have the effects of exercise on thoracic duct lymph flow been studied in normal, intact, instrumented animals. Knott et al. reported that treadmill exercise at 3 mph increased lymph flow in the thoracic duct of intact instrumented dogs from 1.5 to 5.8 ml/min. 13 Recently, we examined thoracic duct lymph flow during a more prolonged period of exercise, 8 min at a treadmill speed of 3 mph. 14 This study showed that thoracic duct flow remained significantly higher than resting flow for the initial 4 min of exercise. Furthermore, this exercise-induced increase in lymph flow was greatly augmented by expanding the extracellular volume. The current study extends understanding of the effects of exercise on lymph flow by examining thoracic duct lymph flow over a range of treadmill speeds from 1.5 to 10 mph.
The major finding of the current investigation was a nearly linear increase in thoracic duct lymph flow as treadmill exercise increased to 10 mph, reaching a flow of 9.0 ± 1.6 ml/min. At a treadmill speed of 4 mph, we observed mean lymph flow 5.0 ml/min. This is similar to the thoracic duct lymph flows reported by Schad and Brechtelsbauer and by Lindena et al. for dogs walking at ∼4 mph. At the most intensive exercise we studied, 10 mph, mean lymph flow had not reached a plateau, although one dog refused to run this fast, and in another dog, lymph flow was lower at 10 mph than at 9 mph. While the dogs in this study had been taught to run on a treadmill prior to instrumentation, they were not exercise conditioned. Thus, it is likely that dogs conditioned to run faster than 10 mph would have higher lymph flows than those observed in this study.
In these experiments, the duration of exercise was limited to 1 min. In a previous study, we observed that exercise at 3 mph could elevate lymph flow significantly for not more than 4 min. 14 Thus, it is likely that the flows reported here for speeds greater than 3 mph would not have been sustained for as long as 4 min. Limiting the duration of exercise to 1 min in this study lessened the possibility of depleting sources of lymph, and was less tiring for the animals.
Similar lymph flows were observed when the exercise protocol was begun at low speed or when the protocol was begun at high speed. Since lymph flow was greatest at high speeds, depletion of lymph sources would have resulted in lesser lymph flow at subsequent lower speeds when speed was varied from 10 mph to 1.5 mph compared to when exercise was begun at 1.5 mph. Conversely, depletion of lymph sources would have resulted in lesser lymph at high speeds when exercise was begun at 1.5 mph compared to when exercise was begun at 10 mph. Since lymph flows were similar for both protocols, the brief 1-min periods of exercise separated by 15 min appear not to have depleted sources of thoracic duct lymph sufficiently to impact lymph flow measures at subsequent treadmill speeds.
The effect of exercise on lymph flow deserves further comment. The lymphatic system transports interstitial fluid and its contents to the central circulation without the aid of a distinct pump (i.e., a heart). 1 Interstitial fluid enters the terminal lymphatic capillaries, which are tethered by anchoring filaments to the surrounding connective tissue. At rest, these filaments maintain open junctions between adjacent endothelial cells, so fluid can enter the lymphatic capillary. When the tissue is compressed, as when nearby muscles contract, the anchoring filaments become slack, tight junctions form at adjacent endothelial cells, and fluid cannot escape from the capillary lumen. Thus, increased tissue pressure causes lymph to flow from the terminal lymphatic capillaries, toward the major lymph vessels, and ultimately to the central blood circulation. Lymph flow is unidirectional due to presence of numerous valves in the lymph vessels. As centrally flowing lymph distends upstream vessels, the smooth muscle of these lymphatic vessels is activated. 1 This myogenic contraction further propels lymph centrally.
Lymph flow reflects the difference in capillary filtration and absorption of fluid. As metabolism increases in exercising muscle, arterioles dilate to increase blood flow and the number of perfused capillaries. 15 This, along with increased venous pressure, 16 elevates capillary blood pressure and increases filtration while impeding absorption.1,17–19 Arteriolar dilation is tightly controlled by the intensity of exercise, 15 so greater filtration would occur with more intensive exercise and produce more fluid for transport by the lymphatic circulation. This exercise-induced hyperemia persists for some time after likely cessation of exercise, with its duration directly proportional to the exercise intensity. 20 This explains, at least in part, the persistence of elevated lymph flow we observed following exercise, which persisted longer with more intensive exercise (Fig. 3).
During exercise, sympathetic nerve activity increases in all tissues, serving to increase cardiac activity and to preferentially distribute the rising cardiac output to contracting skeletal muscles while decreasing visceral flow. 21 Thus, an adequate arterial pressure is maintained to perfuse the dilated vascular beds of exercising muscles, where sympathetic vasoconstriction is counterbalanced by metabolic vasodilatation.22,23 Since the thoracic duct lymph flow measured in this investigation originated from visceral organs as well as from exercising muscle, the increase in lymph flow we observed during exercise likely underestimated the effects of increased filtration and compression in active muscle, since lymph flow from the viscera would likely have decreased during exercise along with blood flow to these organs. Future research should delineate the specific contributions of these various sources to the elevated thoracic duct lymph flow produced by exercise.
We previously reported that application of abdominal lymphatic pump treatment, as performed in osteopathic medical practice, increases thoracic duct lymph flow13,14 and may enhance immune function by mobilizing large numbers of leukocytes via the thoracic lymph duct. 24 In the current study, we have demonstrated that increasing the intensity of physical activity increases lymph flow in the thoracic duct. It is tempting to speculate that lymphatic mobilization of leukocytes would also be positively correlated with exercise intensity. Clearly this important question merits investigation.
Footnotes
Acknowledgments
The expert technical assistance of Linda Howard is gratefully acknowledged.
Author Disclosure Statement
The authors have no conflicts of interest or financial ties to disclose.
This research was supported by National Institutes of Health Grants U19 AT002023 and RN0035. A portion of Dr. Downey's salary was funded by the Osteopathic Research Center.