
Abstract
Abstract
Endothelial nitric oxide synthase (eNOS) and heat shock protein 90 (Hsp90) have been reported to contribute to angiogenesis and lymphangiogenesis. However, the functions of these proteins during lymphangiogenesis are unclear. In the present study, we first observed the cord formation pattern of human dermal microvascular lymphatic endothelial cells (HMVEC-dLy) on Matrigel over 2 to 8 h. The length of cord formation increased, peaked at 4 h, and then started to decline after 6 to 8 h of incubation. siRNA-targeted NOS3 significantly reduced the cord formation ability of HMVEC-dLy cells by 27% relative to control. This result confirmed the importance of eNOS in cord formation by human lymphatic endothelial cells. In addition, immunoprecipitation and Western blotting indicated that the interaction between eNOS and Hsp90 was maximal at 4 h, and then the proteins dissociated. This interaction correlated with the observation of cord formation of human lymphatic endothelial cells on Matrigel. Moreover, we found that the eNOS level decreased as the eNOS and Hsp90 complex disassociated during the late stage of cord formation. An Hsp90 inhibitor, 17-DMAG, was able to inhibit the eNOS and Hsp90 interaction, decrease the level of eNOS, and significantly inhibit cord formation to 38% of the level observed in the control. For the first time, we report that the interaction between eNOS and Hsp90 plays an important role in determining eNOS levels and in regulating cord formation of human lymphatic endothelial cells in vitro.
Introduction
Endothelial nitric oxide synthase (eNOS or NOS3) synthesizes nitric oxide (NO) from L-arginine, NADPH, and oxygen. 8 Several angiogenic factors, such as shear stress and vascular endothelial growth factors (VEGFs), stimulate eNOS-derived NO production. 9 NO mediates angiogenesis by triggering the S-nitrosylation 10 and sGC-cGMP pathways, which promote DNA synthesis, 11 endothelial cell migration, 12 and proliferation, 13 and inhibit apoptosis. 14 NO also enhances endogenous pro-angiogenic factors, such as hypoxia-inducible factor 1α (HIF1α), which upregulates VEGF15,16 and inhibits endogenous anti-angiogenic factors, such as thrombospondin-1 (TSP-1). 17 eNOS can be activated in several steps, and heat shock protein 90 (Hsp90) plays a role in this process. The binding of Hsp90 to eNOS disrupts the eNOS-caveolin-1 inhibitor complex, allowing calcium/calmodulin to bind eNOS and expose a key Ser residue for Akt-dependent phosphorylation, which enhances NO production. 18
Lymphatic endothelial cells express eNOS and Hsp90 in a manner that is similar to blood vascular endothelial cells.19–21 eNOS plays a causal role in VEGF-C-induced lymphangiogenesis, lymphatic hyperplasia, and lymphatic metastasis in vivo. 7 Hsp90 plays a role in angiogenesis; 22 however, the details of its function and how eNOS associates with Hsp90 during lymphangiogenesis remain unknown.
Here, we studied the role of eNOS and Hsp90 in cord formation by human lymphatic endothelial cells in vitro. First, we assessed the cord formation pattern of human dermal lymphatic microvascular endothelial cells (HMVEC-dLy) on Matrigel. Next, we confirmed the role of eNOS in cord formation of human lymphatic endothelial cells using siRNA and determined the levels of eNOS and Hsp90 and the complex during cord formation. Finally, we examined the importance of the eNOS-Hsp90 interaction in regulating the eNOS level and cord formation using an Hsp90 inhibitor (17-DMAG).
Our results provide the first evidence that the interaction between eNOS and Hsp90 is important for eNOS levels and in cord formation of human lymphatic endothelial cells in vitro.
Materials and Methods
Materials
The Hsp90 inhibitor dimethyl 17-[2-(dimethylamino)ethyl]amino-17-demethoxygeldanamycin (17-DMAG) was purchased from BIOMOL International, L.P. (Plymouth Meeting, PA). 3,3′-Dithiobispropionimidateċ2 HCl (DTBP) was purchased from Pierce Biotechnology (Rockford, IL). Dynabeads protein G was purchased from Invitrogen (Oslo, Norway). siNOS3 and the antibodies against eNOS, Hsp90, and PCNA were purchased from Santa Cruz Biotechnology, Inc. (Santa Cruz, CA)
Culture of endothelial cells
Normal human dermal lymphatic microvascular endothelial cells (HMVEC-dLy) were obtained from Takara Bio Inc. (Shiga, Japan) and maintained in an EGM-2MV BulletKit (Takara Bio) at 37°C and 5%CO2.
siRNA transfection
To inhibit NOS3 in human endothelial cells, HMVEC-dLy cells were transfected with NOS3 siRNA (Santa Cruz Biotechnology, Inc.) at a final concentration of 6 nM using the Lipofectamine RNAiMAX reagents from Invitrogen Corporation (Carlsbad, CA). After transfection, the cells were grown for 18 h at 37°C in 5% CO2, trypsinized, and seeded on Matrigel. After the incubation, the cells were used for cord formation analysis on Matrigel, RT-PCR, immunoprecipitation, and Western blotting.
Cord formation on Matrigel
Ninety-six-well plates were coated with 60 μl of Matrigel per well (10 mg/ml), which polymerized after 30 min at 37°C. HMVEC-dLy cells (8 × 103 cells/well) were seeded on the Matrigel and incubated at 37°C. At each time point, the cells were fixed with 4% paraformaldehyde and stained using Mayer's hematoxylin (Muto Pure Chemical Co., Ltd., Japan). The cord formation network was photographed, and the capillary length was measured using an Angiogenesis Image Analyzer (Kurabo, Japan).
Proliferation assay
To find a noncytotoxic dose of the Hsp90 inhibitor, 50 μl of endothelial cells (1.5 × 104 cells/well) was seeded in a 96-well plate and allowed to adhere for 2 h. Fifty microliters of medium containing the Hsp90 inhibitor was added to each well. After incubation, 10 μl of WST-1 reagent from a cell counting kit (DOUJINDO, Kumamoto, Japan) was added, and the plate was incubated for 2 h at 37°C in 5% CO2. The absorbance was measured at 450 nM to determine the percentage viable cells.
Protein extraction from cells on Matrigel, immunoprecipitation, and Western blotting
For the immunoprecipitation of eNOS and Western blotting of Hsp90 to characterize their interaction during cord formation on Matrigel, we used DTBP as a protein cross-linker, as described previously. 23 Endothelial cells that had been cultured on Matrigel were washed with phosphate buffered saline (PBS) (pH 8.0) and incubated with 5 mM DTBP in PBS for 30 min. After three washes with PBS (pH 8.0) containing 0.5 mM PMSF, the cells were incubated in a whole cell lysis buffer [25 mM HEPES (pH 7.7), 0.3 M NaCl, 1.5 mM MgCl2, 0.2 mM EDTA, 0.1% Triton X-100, 20 mM β-glycerophosphate, 1 mM sodium orthovanadate, 1 mM phenylmethylsulfonyl fluoride (PMSF), 1 mM dithiothreitol (DTT), 10 μg/ml aprotinin, and 10 μg/ml leupeptin]. The cells (including the Matrigel) were then scrubbed. The collected samples were vigorously vortexed, and the cytoplasmic extracts were collected from the supernatants by centrifugation at 14,000 rpm for 10 min. Immunoprecipitation was carried out by incubating the lysates with an eNOS polyclonal antibody for 16 h at 4°C and then incubating for 4 h with Dynabeads protein G (Invitrogen). The immunoprecipitates were washed with lysis buffer, resuspended in loading buffer, boiled for 3 min, and subjected to SDS-PAGE on 7.5% polyacrylamide gels, which were then transferred to a PVDF membrane. The primary antibodies used were anti-eNOS and anti-Hsp90. The membrane was then washed and incubated with secondary antibodies coupled to horseradish peroxidase. For detection of the Hsp90 level alone, the cell lysate was subjected to Western blotting using an anti-Hsp90 antibody and processed as described above. PCNA was used as a loading control.
Statistical analysis
The statistical analysis was performed using Dunett's method. P values below 0.05 were considered to be significant.
Results
Cord formation pattern in vitro
To study the characteristics of cord formation by human lymphatic endothelial cells in vitro, HMVEC-dLy cells were seeded on Matrigel and observed after incubation for 2–8 h. At each time point, the cord formation was photographed (Fig. 1A) and the lengths of the cords were measured (Fig. 1B). The graph indicates that endothelial cells started to form networks within 2 h, reached an optimal length at 4 h, and then rapidly decreased. The cord formation of HMVEC-dLy cells on Matrigel usually followed a similar pattern (data not shown). The time course of cord formation was compared with the levels of eNOS, Hsp90, and the eNOS-Hsp90 complex (Fig. 2).

Cord formation patterns of HMVEC-dLy cells seeded on Matrigel for 2–8 h.
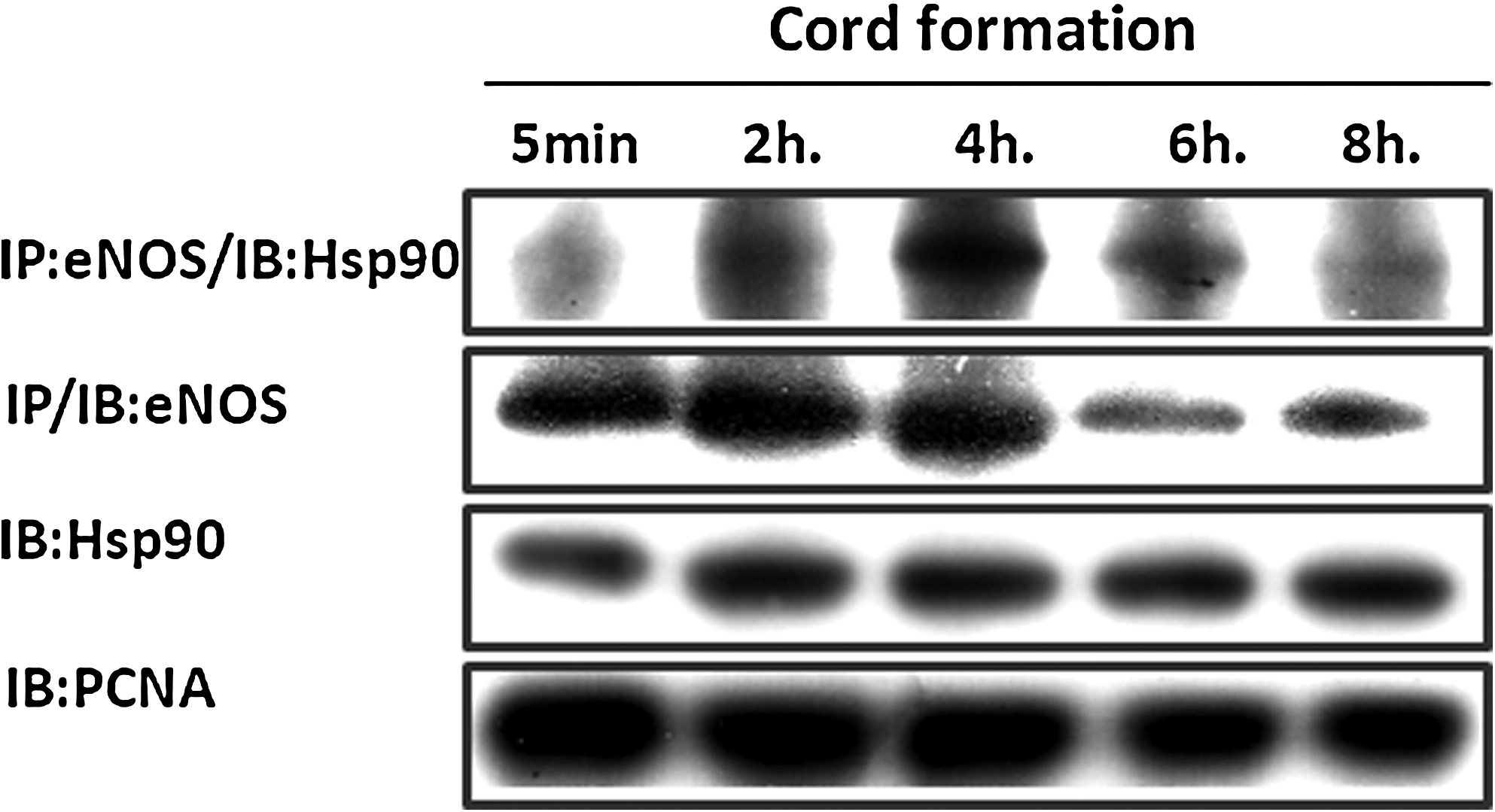
The levels of eNOS, Hsp90 and the complex of eNOS and Hsp90 during cord formation of HMVEC-dLy cells on Matrigel from 5 min to 8 h. Endothelial cells were seeded on Matrigel at 37°C and 5% CO2. After 5 min to 8 h of incubation, the cells were harvested to perform immunoprecipitation (IP) and Western blot analysis (IB) for detection of the eNOS–Hsp90 complex, eNOS, and Hsp90 levels, as described in the Materials and Methods. PCNA was employed as the loading control. Similar results were obtained in three independent experiments.
Influence of eNOS on cord formation in vitro
eNOS has been reported to play a crucial role in lymphangiogenesis in vivo. 7 The present study confirmed that eNOS affects cord formation by human lymphatic endothelial cells in vitro using siNOS3 transfection (Fig. 3). siNOS3-transfected endothelial cells were seeded on Matrigel, and cord formation was examined at 4 h, which is when the maximum numbers of cords were observed. The results indicate that the complete knockdown of the eNOS gene and protein (Fig. 3A) significantly decreased the length of cord formation in human lymphatic endothelial cells (HMVEC-dLy) by 27% compared with the control (Figs. 3B and 3C). The complete suppression of the eNOS and eNOS levels (Fig. 3A) indicates that siRNA transfection was successful.
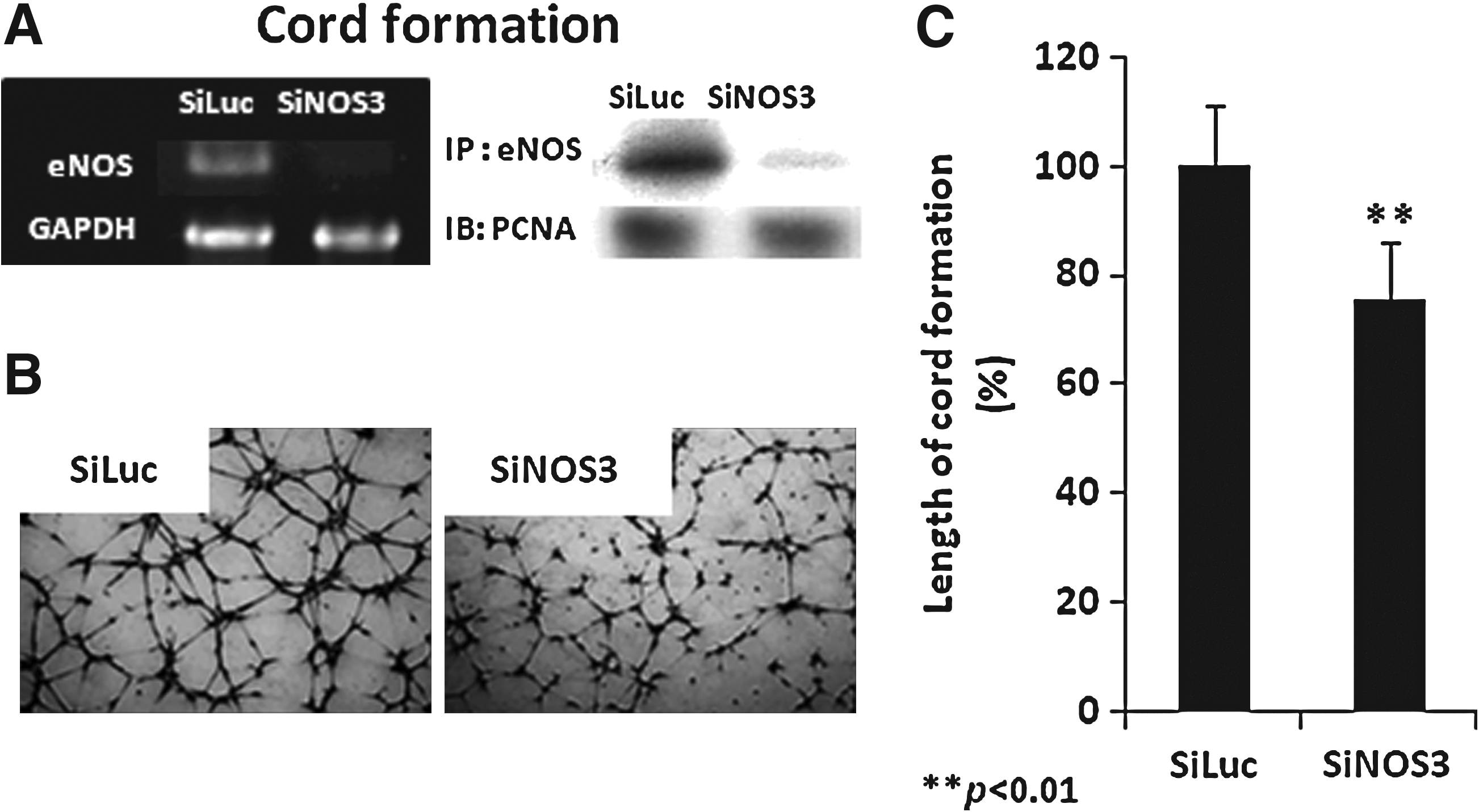
Inhibition of cord formations by NOS3 knockdown. Transfected HMVEC-dLy cells were cultured on Matrigel for 4 h at 37°C and 5% CO2.
The eNOS and Hsp90 interaction correlated with the eNOS level and cord formation by human lymphatic endothelial cells in vitro
eNOS is activated through interaction with Hsp90 and phosphorylation by Akt in blood endothelial cells. 18 However, the role of eNOS and Hsp90 levels, and their interaction in cord formation by human lymphatic endothelial cells, is unclear. In this study, immunoprecipitation and Western blotting were performed to determine the protein and complex levels during cord formation on Matrigel (5 min to 8 h) (Fig. 2). The interaction between eNOS and Hsp90 during cord formation peaked at 4 h and then decreased at 6–8 h. This pattern correlated with relative lengths of cord formation in HMVEC-dLy cells (Fig. 1B). Interestingly, in the late stage of cord formation (6 to 8 h), the eNOS level was decreased as the eNOS and Hsp90 interaction declined (Fig. 2). However, the level of Hsp90 did not change (Fig. 2).
The eNOS and Hsp90 complex is necessary for the eNOS level and cord formation by human lymphatic endothelial cells in vitro
The previous findings (Figs. 2 and 3) allowed us to design an experiment to determine whether the interaction between eNOS and Hsp90 is important for the eNOS level and cord formation by human lymphatic endothelial cells in vitro. The Hsp90 inhibitor 17-DMAG (nontoxic dose, Fig. 4D) was used in this study. Cord formation on Matrigel at 4 h, which represents the maximal level of eNOS–Hsp90 interaction, was chosen to analyze cord formation on Matrigel, immunoprecipitate the complex, and determine protein levels by Western blot in response to the Hsp90 inhibitor. Figure 4A indicates that pretreatment with 350 nM 17-DMAG disrupted the interaction of eNOS with Hsp90, which led to a decrease in the amount of eNOS relative to the control DMSO, whereas the Hsp90 level did not change (Fig. 4A). 17-DMAG also significantly reduced the length of cord formation in HMLVEC-dLy cells to 38% of the control (Figs. 4B and 4C).
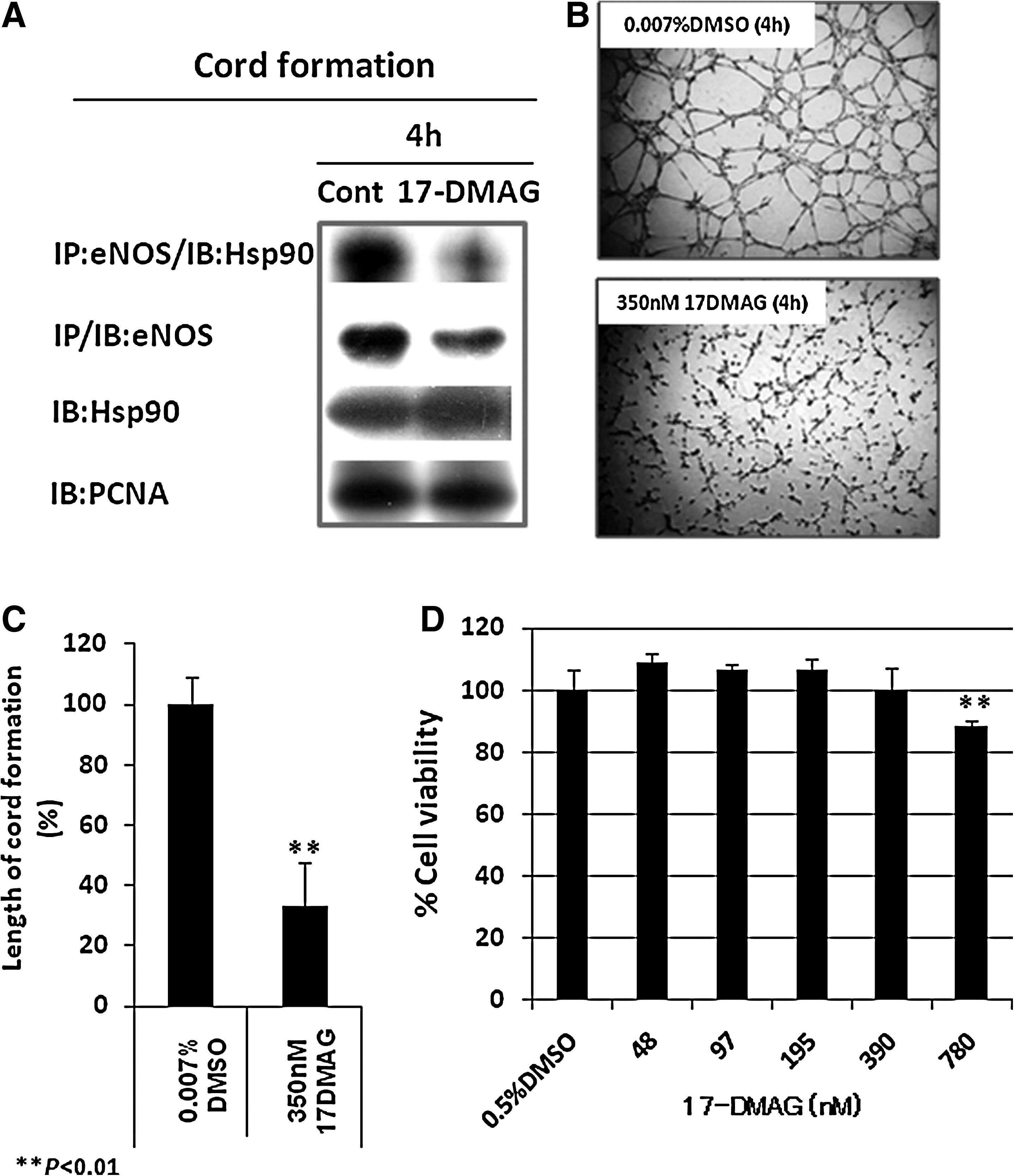
Effect of an Hsp90 inhibitor (17-DMAG) on eNOS and Hsp90 levels, the eNOS-Hsp90 complex
Additional inhibitory effect of Hsp90 on cord formation by human lymphatic endothelial cells in vitro
To determine whether Hsp90 also has an additional effect on cord formation independent of eNOS. siLuc- and siNOS3-transfected HMVEC-dLy cells were preincubated with 350 nM 17-DMAG or control DMSO for 24 h. The cells were trypsinized and seeded on Matrigel, and cord formation on Matrigel was observed after 4 h of incubation. The siNOS3-transfected cells treated with 17-DMAG exhibited cord formation that was significantly reduced to 37% of the control, and there was also the additional effect of a significant inhibition of 11% compared with the siLuc-transfected cells treated with 17-DMAG (Fig. 5). However, cord formation in the eNOS knocked down cells was not completely inhibited by 17-DMAG (Fig. 5).
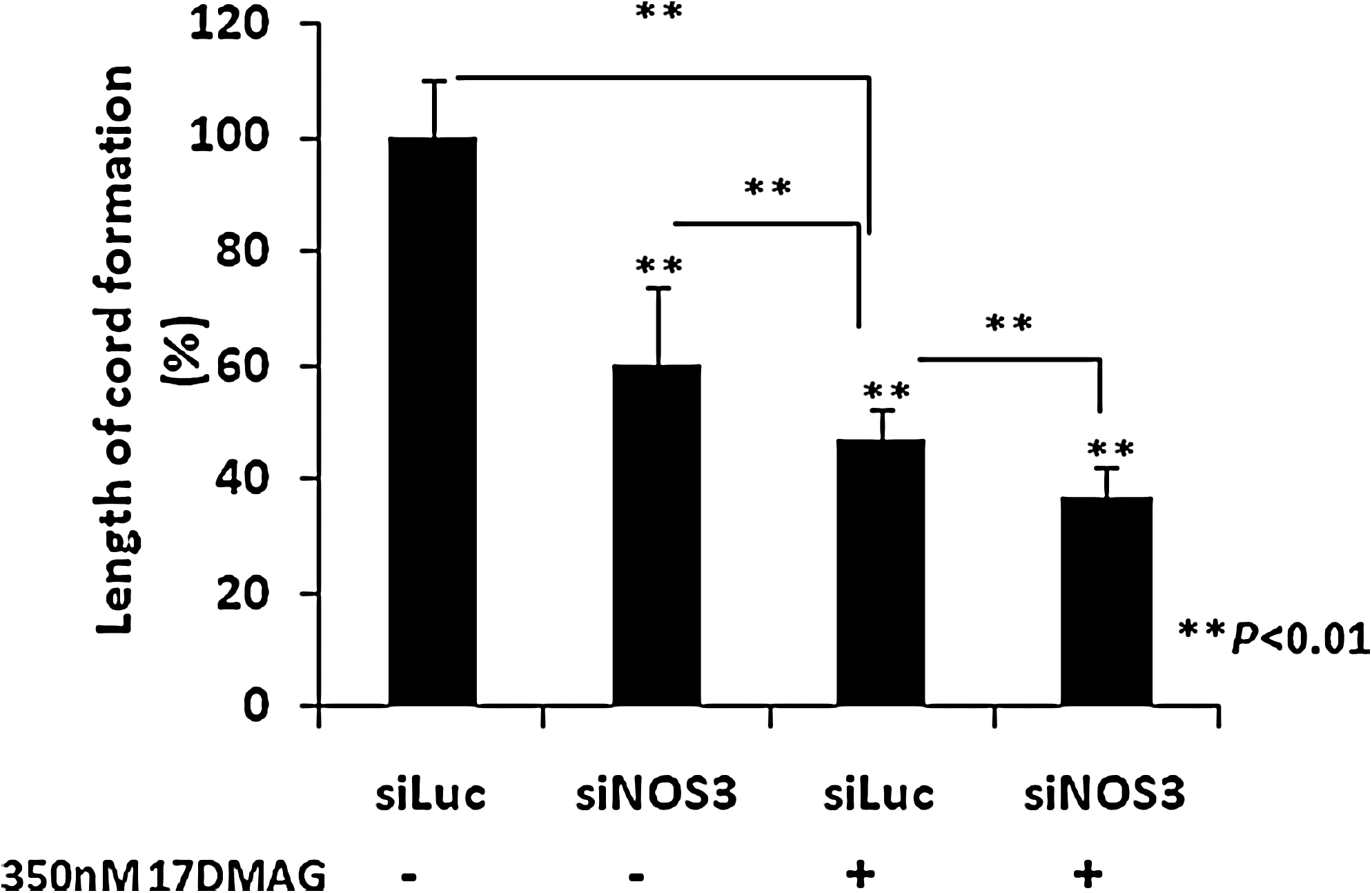
Additional inhibitory effect of Hsp90 on cord formation. siLuc and siNOS3-transfected HMVEC-dLy cells were preincubated with 350 nM 17-DMAG or control DMSO for 24 h. The cells were trypsinized and seeded on Matrigel. Cord formation on Matrigel was measured after 4 h of incubation. The bar graph indicates the lengths of the cords formed in siLuc- and siNOS3-transfected HMVEC-dLy cells pre-treated with or without 17-DMAG, which were measured with the Angiogenesis Image Analyzer. Similar results were obtained in three independent experiments. Data are presented as mean + S.D. (n = 3); **p < 0.01.
Discussion
In the present study, we identified for the first time the roles of eNOS, Hsp90, and the interaction between them during cord formation by human lymphatic endothelial cells in vitro. We found that the interaction between eNOS and Hsp90 is important for the eNOS level and for cord formation in lymphatic endothelial cells.
We modeled the characteristics of cord formation on Matrigel using human lymphatic endothelial cells incubated for 2–8 h (Fig. 1). Cords started to form at 2 h, increased to the maximal length at 4 h, and then declined (Fig. 1B). This pattern of cord formation was used to compare with the levels of eNOS, Hsp90, and the complex of the two molecules (Fig. 2).
It has been reported that eNOS plays a crucial role in lymphangiogenesis in vivo by mediating VEGF-C signaling and lymphatic metastasis. 7 In our study, we confirmed the role of eNOS in cord formation by human lymphatic endothelial cells in vitro using siRNA transfection. Complete knockdown of the eNOS gene (Fig. 3A) led to a significant 27% decreased in cord formation in human lymphatic endothelial cells (HMVEC-dLy) relative to the control. This study indicated that eNOS-mediated cord formation by human lymphatic endothelial cells can be modeled in vitro. eNOS is an enzyme that produces nitric oxide (NO) from L-arginine and is known to participate in angiogenesis. 8 The interaction between eNOS and Hsp90 promotes the activation of eNOS and the release of NO. Human eNOS interacts with Hsp90 at Cys597, which is located in a three-stranded β-sheet that is close to the C-terminal dimeric interface of Hsp90. 24 The function of Hsp90 depends on ATP binding and hydrolysis at the N-terminal domain, which is coupled to a conformational cycle of opening and closing dimeric molecular clamp that is formed for the capture of client proteins such as eNOS. 25
Our study indicated that eNOS and Hsp90 interacted during cord formation by human lymphatic endothelial cells on Matrigel (Fig. 2). We found that the pattern of eNOS and Hsp90 interaction in human lymphatic endothelial cells (Fig. 2) directly correlated with the length of cord formation (Fig. 1) on Matrigel, which was maximal at 4 h and decreased at 6 to 8 h. We confirmed that the interaction between eNOS and Hsp90 is important for the formation of cords by endothelial cells using an Hsp90 inhibitor (17-DMAG). This inhibitor binds to the N-terminal domain ATP-binding site of Hsp90 and inhibits its chaperone activity. 26 After preincubation of lymphatic endothelial cells with a nontoxic dose of 17-DMAG and seeding on the Matrigel, the interaction of eNOS with Hsp90 was inhibited (Fig. 4A), leading to cord formation in the lymphatic endothelial cells that was significantly decreased to 38% of the control (Fig. 4C). This result indicated that the eNOS and Hsp90 complex is important for cord formation by human lymphatic endothelial cells in vitro. This is the first study to indicate that an Hsp90 inhibitor (17-DMAG) inhibited both the eNOS and Hsp90 interaction and the formation of cords in human lymphatic endothelial cells (Fig. 4).
In addition, Figure 5 indicates the significant decrease in siNOS3-transfected cells pretreated with 17-DMAG relative to siLuc-transfected cells pretreated with 17-DMAG. This result indicated that, at least in part, the eNOS/Hsp90 complex participated in cord formation of human lymphatic endothelial cells on Matrigel. Moreover, the significant decrease in 17-DMAG pretreatment with siLuc relative to siNOS3 transfection alone indicated that 17-DMAG also inhibits cord formation through a non-eNOS mechanism.
It has been reported that 17-DMAG has antiangiogenic properties in vitro and in vivo.27,34 In vitro treatment of HUVEC with 17-DMAG significantly inhibited migration, extracellular matrix invasion, and cord formation on Matrigel. 17-DMAG inhibited VEGF- and FGF-2-induced HUVEC proliferation and resulted in apoptosis. In addition, this compound also inhibited the expression of protein kinases involved in the angiogenesis pathways of endothelial cells, which are also known to be Hsp90 client proteins (pAkt and c-Raf-1) or their downstream substrates (pERK). 27 In human lymphatic endothelial cells, 17-DMAG inhibited VEGFR-3, which is a major receptor in lymphatic endothelial cells that signals downstream induction of lymphatic neovascularization. 21 The additional inhibitory effect of 17-DMAG on cord formation of eNOS knocked down cells (Fig. 5) may have been influenced by these factors.21,27,34
In this study, we also found that the eNOS level decreased at the same time as the eNOS and Hsp90 complex declined during the late stage of cord formation (Fig. 3). The use of 17-DMAG indicated that the interaction between eNOS and Hsp90 was necessary for regulation of the eNOS level in lymphatic endothelial cells (Fig. 4A). This inhibitor induced dissociation of eNOS from Hsp90, and the eNOS level was decreased (Fig. 4A). The decrease in eNOS levels in the late stage of cord formation in Figure 3 can be explained by the inhibition of Hsp90 function that causes the eNOS and Hsp90 complex to dissociate. It has been reported that eNOS can be degraded by calpain or by the ubiquitination system.28,29 The disruption of the folding of Hsp90 client proteins leads to ubiquitination and subsequent proteolytic degradation by the proteasome. 29 In addition, the inhibition of Hsp90 and eNOS complex by geldanamycin (Hsp90 inhibitor) leads to eNOS proteolysis by calpain in a cell-free system. 30 However, the mechanism of ubiquitination and calpain activation during cord formation by human lymphatic endothelial cells in vitro was not examined in this study.
Dysregulation of Hsp90 during the late stage of cord formation (Fig. 2) may result from an increase in ROS. 31 The recruitment of the eNOS-Hsp90 complex during the initial stage of cord formation (2–4 h) (Fig. 2) could generate NO, which promotes oxidative stress and damages cells. 32 Hsp90 acts as a molecular sensor of the local NO concentration and adjusts NO production. 31 NO can react with Cys597 of Hsp90 (S-nitrosylation), which inhibits Hsp90 ATPase activity, disrupts the eNOS–Hsp90 complex, and reduces eNOS activation. 33 Then, during the late stage of cord formation at 6–8 h, the decrease in Hsp90 function leads to a dissociation of the eNOS–Hsp90 complex (Fig. 2), which decreases the eNOS level (Fig. 2) and cord formation (Fig. 1). Next, we are planning to investigate NO production, ROS generation and S-nitrosylation during cord formation by human lymphatic endothelial cells in vitro.
In summary, we have provided evidence that the interaction of eNOS with Hsp90 plays an important role in the regulation of the eNOS level and in cord formation by human lymphatic endothelial cells in vitro. These results provide novel clues for lymphatic biology and targets for anti-lymphangiogenic therapy.
Footnotes
Acknowledgments
This work was supported in part by Grants-in-Aid for Scientific Research (C) (No. 22501042 and 22591561) from the Japan Society for the Promotion of Science (JSPS).
Author Disclosure Statement
No competing financial interests exist.
O. Prangsaengtong and K. Koizumi contributed to this work equally.