
Abstract
Abstract
Background:
Lymphatic endothelial cells from primary lymphedema skin have never been cultured nor characterized. A subgroup of patients with primary lymphedema undergo surgery to bring about an improvement in their quality of life. The aim of this study was to culture and characterize LECs from the skin of these patients.
Methods and Results:
Lymphatic endothelial cells were isolated and cultured from the skin of patients with primary lymphedema and from normal skin. The isolated cells were compared in their ability to form microvascular networks in a three-dimensional culture medium, and in their response to treatment with vascular endothelial growth factors A, C, and D. Whole tissue transcriptional profiling was carried out on two pools of isolated lymphatic endothelial cells—one from primary lymphedema skin and the other from normal skin. Lymphatic endothelial cells from primary lymphedema skin form tubule-like structures when cultured in three-dimensional media. They respond in a similar fashion to stimulation with the vascular endothelial growth factors A, C, and D. Comparative analysis between lymphedema tissue and normal tissue (fold change >2) showed differential expression of 2793 genes (5% of all transcripts), 2184 upregulated, and 609 downregulated. Genes involved in cellular apoptosis (vascular endothelial growth inhibitor, zinc finger protein), extracellular matrix turnover (matrix metalloproteinase inhibitor-16), and type IV collagen deposition were upregulated. Various pro-inflammatory genes (interleukin-6, interleukin-8, interleukin-32, E-selectin) were downregulated.
Conclusion:
Cellular adhesion, apoptosis, and increased extracellular matrix turnover play a more prominent role in primary lymphedema than previously thought. In addition, the acute inflammatory response is attenuated as evidenced by the downregulation of various pro-inflammatory genes.This sheds further light on the interplay of the various pathological processes taking place in primary lymphedema.
Introduction
Nonne, in 1891, was the first to describe congenital lymphedema of the lower limbs. 2 Milroy, in 1892, described the same condition affecting 26 persons in a single family, spanning six generations. 3 This form of lymphedema, characterized by unilateral or bilateral lower limb swelling, with a family history, was classified as primary or congenital lymphedema. Other forms of heritable lymphedema have since been described, including Meige's disease and lymphedema distichiasis. Meige's disease is primary lymphedema of the lower limbs, presenting in puberty, with a genetic predisposition in 30 percent of cases. 4 The lymphedema is usually mild, rarely extends above the knee and is generally bilateral. Lymphedema distichiasis is congenital lymphedema associated with developmental abnormalities of the eyelids. 5 Milroy disease 6 and lymphedema distichiasis 7 have been linked to specific gene mutations on chromosomes five (5q34–q35) and 16 (16q24.3), respectively. Defective vascular endothelial growth factor receptor three (VEGFR-3) signaling has been identified as the cause of congenital hereditary lymphedema in Milroy disease. 8 The VEGFR-3 receptor binds the ligands vascular endothelial growth factor (VEGF)-C and VEGF-D. Lymphedema distichiasis, on the other hand, is caused by a mutation in the gene expressing the transcription factor FOXC2. 9
Lymphatic disease is quite prevalent, and often not well clinically characterized.10,11 Apart from lymphedema, other human diseases directly or indirectly affect or alter lymphatic structure and function, with a wide array of clinical signs and symptoms. 12 Progress in our understanding of the molecular mechanisms controlling the lymphatic system has lead to new insights in the role of lymphatic endothelium in inflammation, vascular disorders, and cancer dissemination.13–15
This study focused on a small subgroup of patients with severe lower limb or genital primary lymphedema. These patients with massive limb swelling or genital lymphedema are often offered surgery, after an initial trial of conservative management. Limb and genital reduction surgery has been shown, in patients with severe primary lymphedema, to provide symptomatic relief, and an improvement in quality of life. 16 Lymphatic endothelial cells have been isolated from animals and normal human skin, using antibodies to lymphatic endothelial cell markers.17–23 The hyaluronan receptor LYVE-1 and the transmembrane glycoprotein podoplanin are expressed by LECs in vitro17–21 and antibodies to these cell markers have been used to isolate LECs from normal human and bovine skin.
The aims of this study were to isolate and characterize LECs from the skin of patients with primary lymphedema. The ability of these LECs to form tubules, their response to stimulation with growth factors, and the gene profile expressed by these cells was then compared to that of LECs isolated from normal skin.
Patients, Materials and Methods
The Academic Department of Surgery, St. Thomas Hospital, London, is a tertiary referral center for patients being considered for lymphedema surgery. Patients with primary lymphedema that were referred to the surgical outpatient clinic for consideration for surgery fell into two main groups:
those with massive unilateral or bilateral limb swelling. those with genital swelling.
Control skin was obtained from patients undergoing cosmetic plastic surgical procedures, such as breast reduction surgery or abdominoplasty procedures (excision of redundant abdominal wall skin). Normal breast and abdominal skin is not an ideal ‘control’ for limb and genital lymphedema tissue, however, there are few procedures carried out on the legs that would permit the excision of skin for research purposes. Samples of skin were obtained with ethical approval and informed consent (St. Thomas's Hospital ethics approval number EC 02/064) from a total of 23 patients.
Isolation and cell culture of LECs
Skin samples were obtained and incubated overnight with dispase (50 U/ml; Gibco) for 30 min at 37°C, with subsequent removal of the epidermis as previously described. 17 Endothelial cells were released from the skin by squeezing and scraping. The endothelial cells were then pelleted, and re-suspended in endothelial cell growth medium (EBM-2, Cambrex Bio Science, UK). Cells were then cultured on gelatin-coated flasks to obtain a colony of cobblestone cells.
Purification of LECs
Cells were detached from the culture flasks by incubation for 10 min at 37°C with accutase (PAA laboratories, Somerset, UK). 10 ml of MACS buffer (2 mM EDTA, 0.5% bovine serum albumin in PBS; Miltenyi Biotech, Surrey, UK) was then added to the cell solution to neutralize the action of the accutase. Cells were incubated with 340 μg/ml of the lymphatic specific antibody rabbit anti-LYVE1 polyclonal antibody and rabbit anti-podoplanin antibody (Research Diagnostics, Inc.) at 4°C for 30 min. The cells were centrifuged at 210 g force and then re-suspended in 2 ml of MACS buffer. The antibody-coated LECs were purified from this suspension by incubation with goat anti-rabbit IgG magnetic micro beads (MACS micro beads; Miltenyi Biotech) at 4°C for 30 min. The cell suspension was then passed through a magnetic column (Miltenyi Biotech) and re-suspended in 5 ml of EBM-2. This was then cultured on gelatin-coated flasks for 2–5 days until a confluent colony of cells was obtained.
Immunocytochemical staining of isolated cells
Antibodies to the endothelial cell markers CD31 and CD44 and lymphatic specific markers (anti-LYVE 1 and anti-podoplanin) were used. Although CD44 is a common protein expressed on most blood vascular endothelial cells, blood cells, epithelial cells and stromal cells, its absence from LECs makes it a useful negative marker.13,17 Cells purified and cultured as described above, were detached from culture flasks by incubation for 10 min at 37°C with accutase. 0.5 ml of cell suspension was seeded onto separate chambers on an 8-chamber slide (Sigma, UK), and cultured for 48 h to reach confluence. Endothelial cells were then washed with phosphate-buffered saline (PBS; Sigma, UK), and incubated for 30 min at room temperature with rabbit anti-LYVE1 antibody (2 μg/ml) and one of either mouse anti-podoplanin antibody (2 μg/ml, Research Diagnostics, Inc), mouse anti-CD31 antibody (8 μg/ml in PBS, Dako, Cambridge, UK) or mouse anti-CD44 antibody (8 μg/ml. in PBS, Dako). Cells were then washed with PBS, and incubated with Alexa fluor 568 conjugated goat anti-rabbit IgG and Alexa fluor 488 conjugated goat anti-mouse (2 μg/ml in PBS, Molecular Probes, UK) for 30 min at room temperature. The slides were then washed with PBS, and mounted with vectashield containing the blue fluorescent nuclear stain DAPI (Vector Laboratories, Peterborough, UK). Images were captured with a Leica (Leitz DMRB) fluorescent microscope at X 200 eyepiece magnification.
Tubule formation assays
Lymphatic endothelial cells from all three sources (normal skin, limb, and genital lymphedema skin) were cultured on a three-dimensional medium (Matrigel). This was to assess whether the abnormality in this group of patients with primary lymphedema was due to an inability of the LECs to adhere and interact with each other in a three-dimensional matrix. A positive finding would be the inability of LECs isolated from limb or genital lymphedema skin to form capillary-like networks when cultured on Matrigel.
To assess in vitro tubule formation, confluent LECs (2.5 × 105) were seeded onto Matrigel-coated 24-well culture plates (BD Biosciences, Oxford, UK). 24 Culture plates were inspected under phase contrast light microscopy at 24 h to assess for the formation of tubules.
Proliferation assays with XTT
Lymphatic endothelial cells from different sources were compared in their ability to respond to the growth factors VEGF-A, VEGF-C, and VEGF-D. The controls for each of these experiments were the same set of LECs, untreated with any growth factor. This was to determine if defective VEGF-C, -D/VEGFR-3 signaling was linked to the etiology of primary lymphedema in this subgroup of patients. The authors expected to find significant differences (normal response) in the proliferation curves of LECs from all sources skin treated with VEGF-A (comparing treated and untreated LECs). In addition, the authors expected to find a similar response in the proliferation curve of LECs from normal skin treated with VEGF-C and VEGF-D. However, in the case of LECs from limb and genital lymphedema skin treated with VEGF-C and VEGF-D, the authors expected to find an impaired response (assuming the defect in this subgroup of patients was linked to defective VEGF-C, -D/VEGFR-3 signaling).
LECs (5 × 104) were seeded onto fibronectin-coated 96-well plates. Triplicate dishes were treated without or with 100 ng/ml of VEGF-A, VEGF-C, and VEGF-D (R & D Systems, Abingdon, UK) in endothelial growth medium. Culture plates were then incubated at 37°C, and the rate of cellular proliferation measured by the addition of XTT and estimation of the color change on a colorimeter. 25
The cell solution (90 μl), containing 2 × 104 LECs /ml was pipetted into 96-well plates. Endothelial growth medium (10 μl) containing 100 ng of growth factor (one of either VEGF A, VEGF C, or VEGF D) or 10 μl of medium alone (controls) was added into each well. Four identical plates were set up for each LEC isolate, and the number of cells in each well measured on days 0, 2, 4, and 7 using the XTT ({2, 3-bis (2-methoxy-4-nitro-5-sulfophenyl)-5-[.(phenylamino) carbonyl ]-2H-tetrazolium hydroxide}) method 2. 25
Proliferation curves and statistical analysis
To estimate the rate of proliferation of each LEC type, the XTT absorbance ratio was plotted against time, using the graph pad software package PRISM. The XTT absorbance ratio was obtained by dividing the values for the absorbance of XTT on days 0, 2, 4, and 7, by the value obtained on day 0. The mean absorbance ratio (and standard error of the mean) for each LEC type (normal LECs and LECs from limb or genital lymphedema skin) was then calculated at the different time points (days 2, 4, and 7).
This produced different proliferation curves for each LEC type treated with the growth factors VEGF-A, VEGF-C, VEGF-D, and for untreated LECs. The treatment of each group of the three LEC types with growth factor VEGF-A, -C, or -D was then compared with untreated LECs by a two-way analysis of variance (ANOVA), with the graph pad software package PRISM.
RNA extraction
Isolated LECs from five lymphedema tissue samples and five normal skin samples were pooled into two separate samples, respectively. This was due to the fact that it was difficult to obtain adequate RNA for microarray analysis from a single sample from one individual. Total RNA was isolated using the manufacturer's protocol (RNeasy Mini kit, Qiagen, West Sussex, UK). The cell pellets were centrifuged for 5 min at 300 g and the supernatant carefully removed by aspiration. The cells were then lysed using buffer RLT (Qiagen). The lysate was homogenized by passing it through a blunt 20-gauge needle connected to an RNAse-free syringe, repeating the process five times. 70% ethanol was then added to the lysate and the sample transferred to an RNeasy spin column which was then centrifuged and the flow-through discarded. Buffer RW1 (Qiagen) was then added to the spin column and then centrifuged at 8000 g for 15 sec. DNAse I incubation mix was then added directly to the spin column and left to stand for 15 min at room temperature. This optional step was added in order to decrease possible contamination with genomic DNA, which could affect the specificity and sensitivity of the downstream microarray experiment. The spin column was then washed by centrifugation with buffer RPE (Qiagen). Total RNA was then eluted by adding 50 μl of RNAse-free water to the spin column and stored at −80°C. The purity of the RNA was then analyzed using the Bioanalyser 2100 (Agilent, Santa Clara, CA) and the Nanodrop (ND-100).
Microarray analysis
The microarray analysis was performed to look for a differential fold change between LECs from lymphedema skin and normal skin, respectively. The aim of the microarray experiment was to look for putative genes involved in the pathogenesis of primary lymphedema. Biotin-labeled cDNA was prepared from the total RNA using reagents and protocols provided by Affymetrix (Affymetrix, Santa Clara, CA). The labeled cDNA samples were then hybridized to the Human Genome U133 Plus 2.0 microarray (Affymetrix). This genechip is an oligonucleotide microarray that allows detection of up to 54,675 transcripts encompassing the whole human genome. The genechip is then stained with streptavidin–phycoerythrin which binds to biotin and fluoresces under the laser in the scanner. The intensity of the fluorescence reflects the level of expression of the gene. The quality of the raw data from the genechips were analyzed with the GCOS system (Affymetrix).
The microarray data from the two samples were imported into the Genespring software (Agilent Technologies). Differential gene expression between the two samples was analyzed. The differentially expressed genes were then classified according to the following molecular functions: apoptosis, proteolysis, inflammation, and angiogenesis. Gene ontology analysis was carried out by uploading the differentially expressed data into GeneCoDis2 (www.genecodis.dacya.ucm.es), a web-based microarray analysis tool. 26 The GeneCoDis2 program determines annotations (cellular component, biological process, and molecular function) that are over-represented in the differentially expressed group of genes with respect to a reference set of genes. Statistical analysis is carried out using hypergeometric distribution 27 and significant annotations with a p value <0.05 are computed. Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway analysis was performed for the differentially expressed genes and scored for significance.
Results
Isolation from primary lymphedema skin
Fifteen patients with primary lymphedema consented to the use of skin samples from tissue excised at surgery. LECs were successfully isolated from the skin of 13 out of these 15 patients (Table 1). Eight had limb lymphedema and five had genital lymphedema. In addition, LECs were successfully isolated from 10 normal skin samples. The initial process of isolation of LECs from primary lymphedema skin yielded a confluent layer of cobblestone cells in 2–5 days (Fig. 1). Immunofluorescence staining of this unselected cell population, confirmed co-localization of the lymphatic marker LYVE-1 and the pan-endothelial cell marker CD31 to a subset of cells (Figs. 2A–2C). Purification of this unselected cell population yielded a colony of confluent colony of cells that stained positive for both the lymphatic markers LYVE-1 and podoplanin (Figs. 3A–3C).

Primary culture of unselected endothelial cells from primary lymphedema skin. Bar = 100 μm.

Immunofluorescent staining of a slide of endothelial cells isolated from primary lymphedema skin. The lymphatic marker LYVE-1
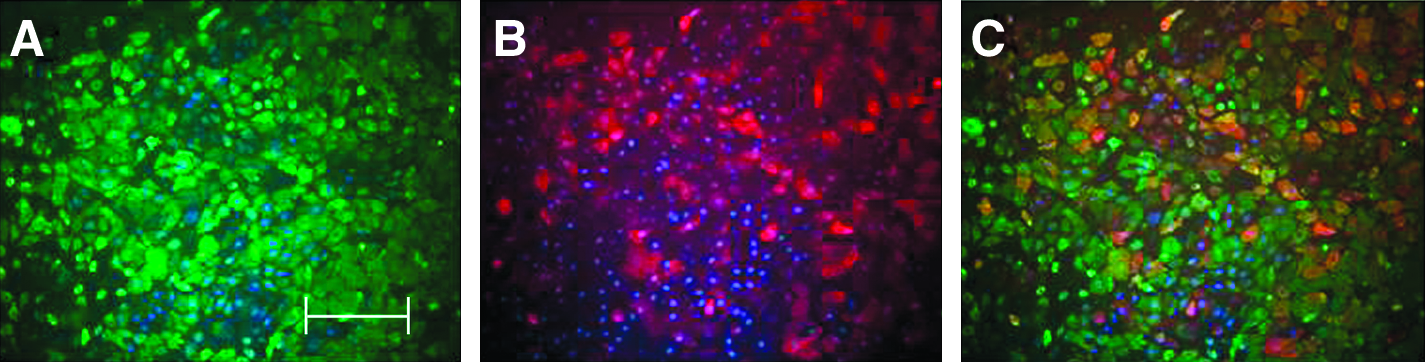
A slide of purified LECs from primary lymphedema skin, stained with the lymphatic markers LYVE-1 (
Primary lymphedema LECs form vascular tubes on Matrigel
There was no difference in the ability of LECs from all sources to form networks on three-dimensional matrix (Matrigel) (Figs. 4A–4C).
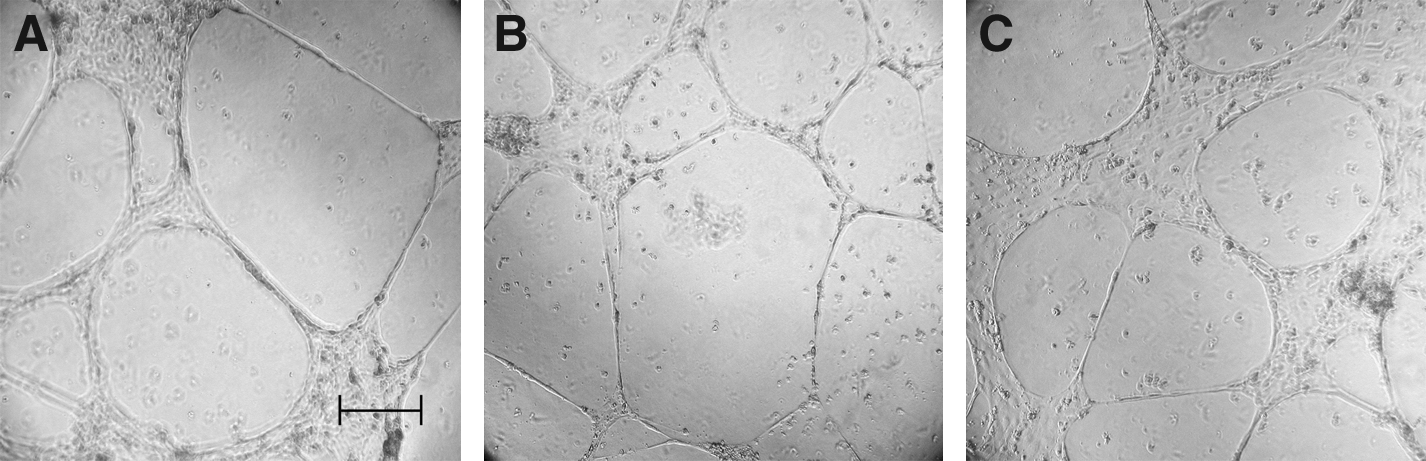
Lymphatic endothelial cells from normal skin
Proliferation of LECs in response to VEGF treatment
VEGF-A
VEGF-A treatment significantly increased the rate of proliferation of all three LEC types (P < 0.01, Figs. 5A–5C).
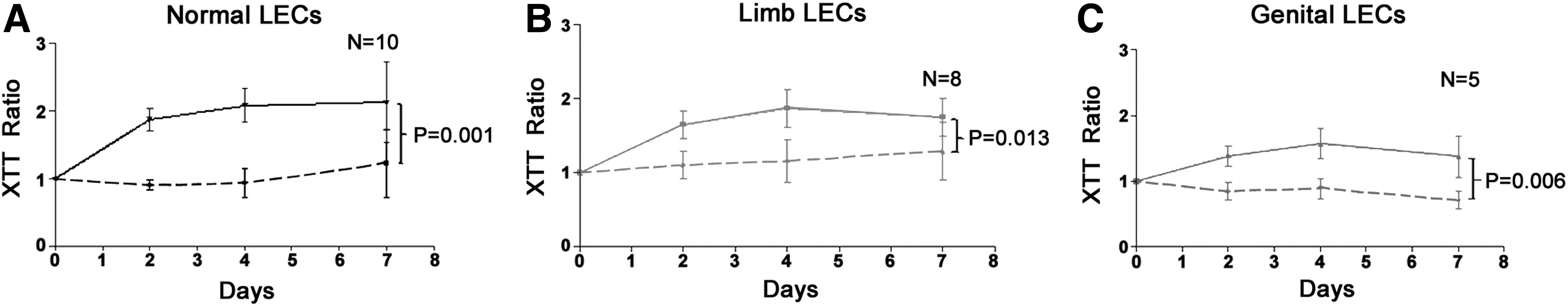
The effect of VEGF-A on LECs isolated from
VEGF-C and VEGF-D
VEGF-C and VEGF-D treatment on all LECs isolated from lymphedema and normal skin had very little effect on cellular proliferation (Figs 6A–6C and Figs. 7A–7C).
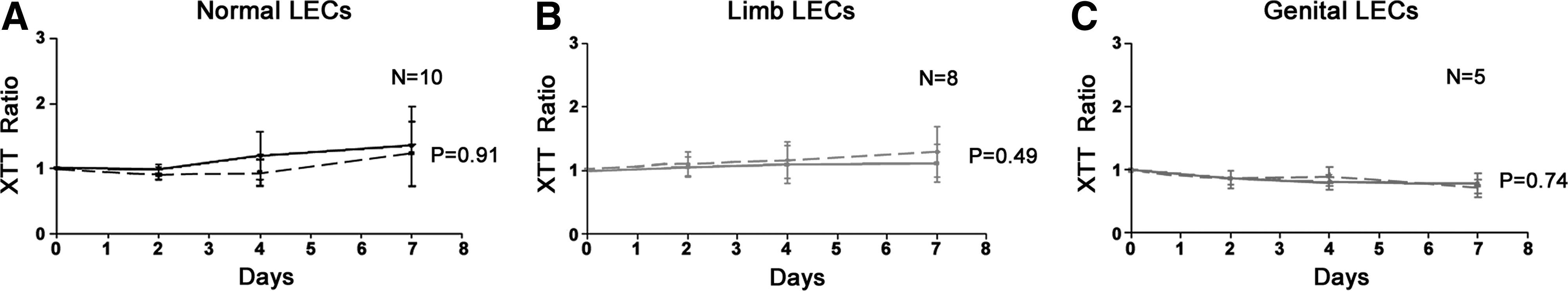
The effect of VEGF-C on LECs isolated from
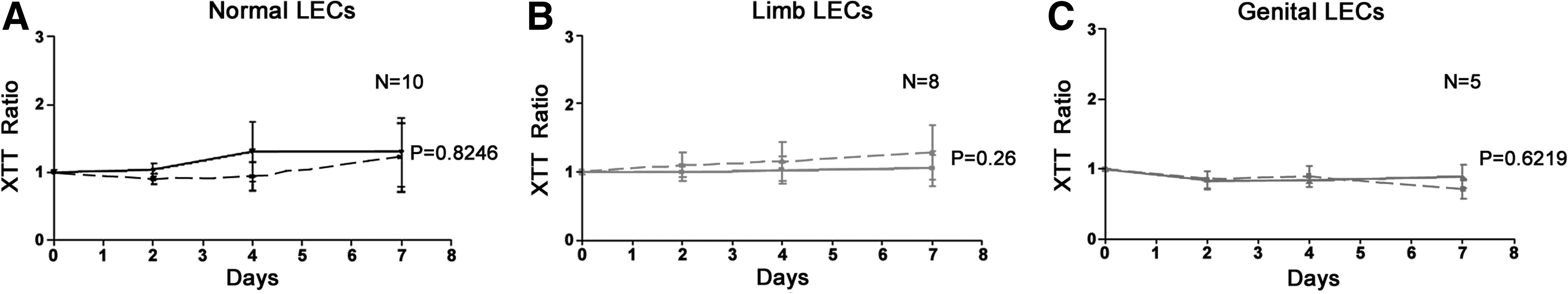
The effect of VEGF-D on LECs isolated from
RNA extraction
The RNA extracted for both samples were of high quality. Both samples had an RNA integrity number (RIN) of 9.4 (RIN > 5.5, pure RNA). The quality of the RNA as measured by the A260/280 ratio on the nanodrop was 1.9 in each case (1.9–2.1, pure RNA). The nanodrop results also confirm minimal contamination with genomic DNA, protein, and phenol.
Microarray analysis
cDNA were successfully prepared from the RNA samples. Comparative analysis between lymphedema tissue and normal tissue (fold change > 2) showed differential expression of 2793 genes (5% of all transcripts), 2184 genes were upregulated, and 609 downregulated. Differentially expressed genes (3-fold or more) comprised genes involved in the processes of inflammation, cellular adhesion and motility, apoptosis, extracellular matrix turnover, and collagen deposition (Table 2).
KEGG pathway analysis was also used to group the differentially expressed genes into known biochemical or functional pathways (Fig. 8 and Fig. 9). Genes that were upregulated in LECs from primary lymphedema tissue were significantly involved in the following pathways: cancer, cytokine–cytokine, chemokine, insulin signaling, and calcium signaling. Genes involved in the VEGF signaling pathway, known to play an important role in lymphangiogenesis, 28 were also upregulated in LECs from lymphedema skin, as opposed to LECs from normal skin (Table 2).
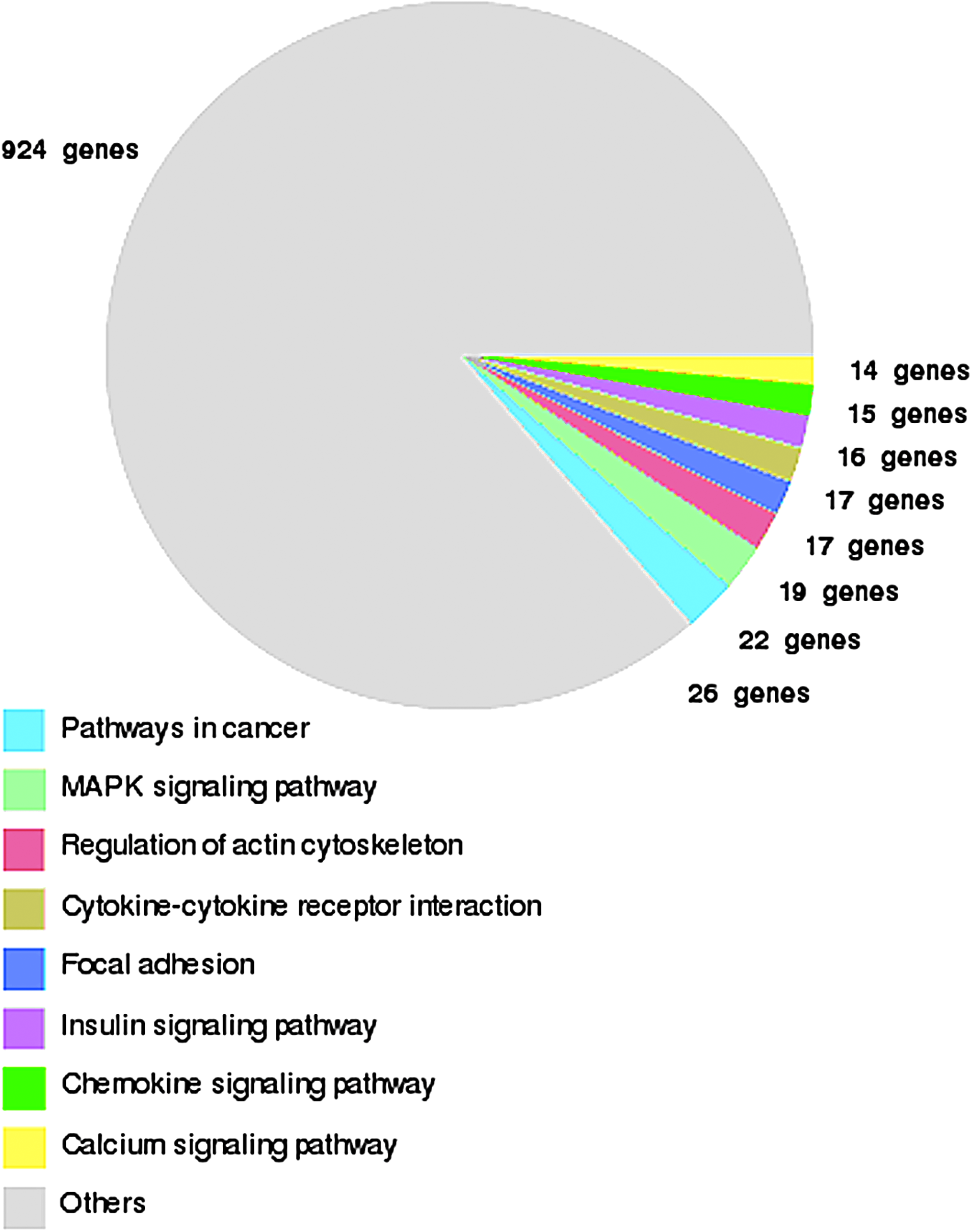
KEGG pathway summary for upregulated genes in LECs isolated from primary lymphedema skin. Genes are grouped into known biochemical or functional pathways. This figure is available in color in the online version of this article at www.liebertpub.com.
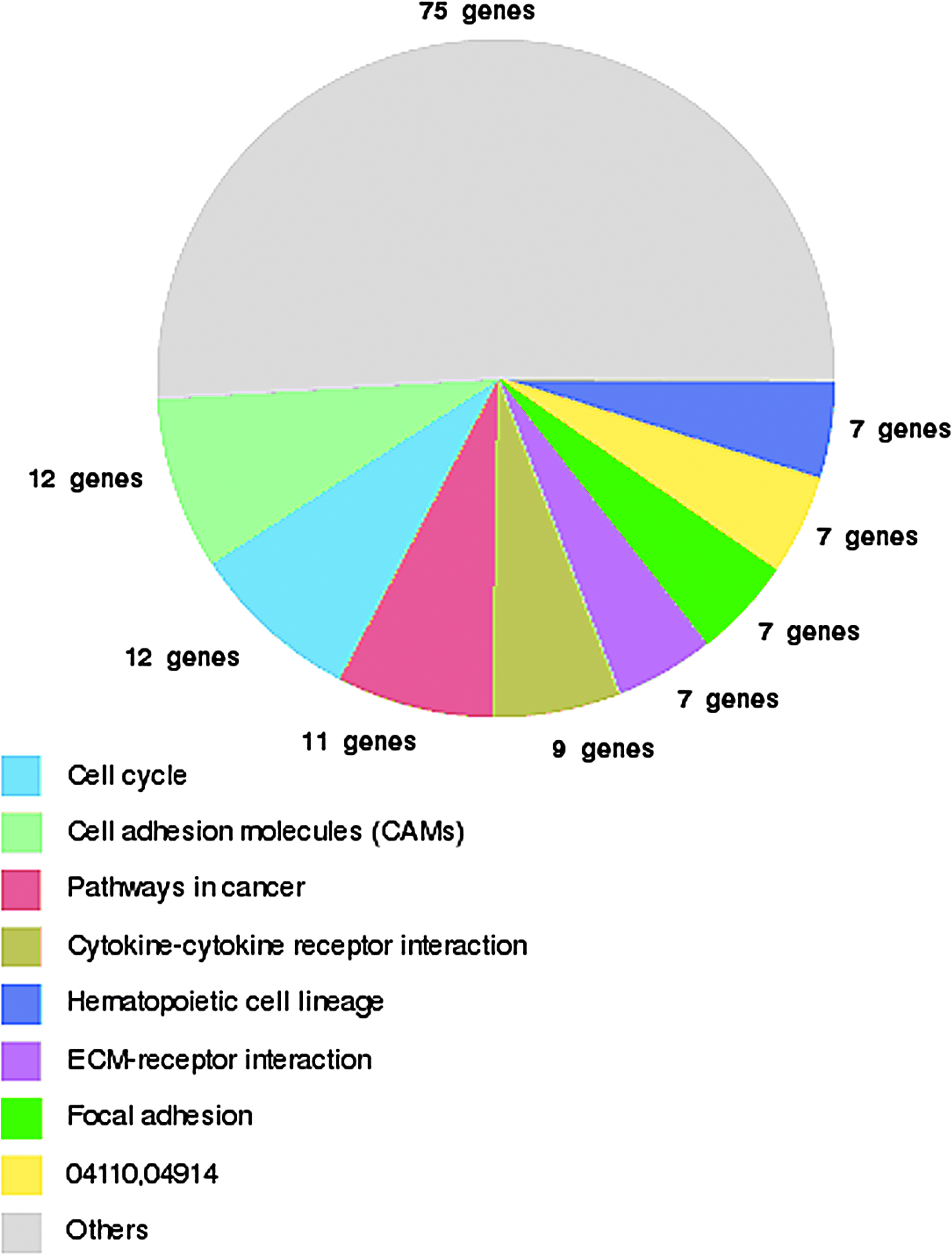
KEGG pathway summary for downregulated genes in LECs isolated from primary lymphedema skin. Genes are grouped into known biochemical or functional pathways. This figure is available in color in the online version of this article at www.liebertpub.com.
Discussion
Techniques for isolating LECs using the lymphatic specific antibodies anti-LYVE1 with anti-podoplanin have allowed us to separate and culture LECs from primary lymphedema skin and compare it to LECs from normal skin.
LECs isolated from both normal and primary lymphedema skin have the ability to form capillary-like tube structures when cultured for 24 h on growth factor-depleted matrigel. This is independent of the effect of any of the vascular endothelial growth factors. These findings are consistent with those obtained by Podgrabinska et al, 20 who studied tube formation in LECs and BECs (blood endothelial cells) from neonatal foreskin, cultured on a collagen gel ‘sandwich’ assay for 48 h. It appears therefore that the lymphedema in these patients may not be the result of an inherent inability of their LECs to form tubules.
Normal, genital, and limb lymphedema LECs, cultured in endothelial growth medium, all respond in a similar fashion to VEGF A. Veikkola et al. 21 reported similar findings with normal LECs isolated from human dermal microvascular endothelial cells treated with VEGF-A. They showed that the proliferation of these LECs, in response to VEGF-A, was two-fold higher than untreated cells.
VEGF-C and VEGF-D appeared to have little to no effect in isolation on LECs isolated from any of the three sources, when cultured in vitro. Veikkola et al. 21 reported small differences (1–2 fold increase) in cell proliferation on stimulation of LECs with VEGF-C in vitro. This variation in their results compared to those from this study may be due to differences in the constituent composition of the endothelial culture mediums.
It would therefore appear that LECs from primary lymphedema skin respond in a similar fashion to the growth factors VEGF-A, -C, and -D, as LECs from normal skin. It may be that defective VEGFR-2 and VEGFR-3 signaling may not play a role in the etiology of this subgroup of primary lymphedema. The presentation and management of this subgroup of patients with primary lymphedema is completely different from the much rarer Milroy's disease phenotype. 16 It is therefore reasonable to guess that the etiology of lymphedema in these patients may be unrelated to VEGFR-3 signaling.
In arriving at the results of the proliferation studies, we have made two assumptions. First, that LECs isolated from different patients, with the same clinical subtype of primary lymphedema, will respond in a similar fashion to the growth factors VEGF-A, -C, and -D. This may apparently not be the case as there may be variability in the individual responses to stimulation. In addition, we have also assumed that normal LECs from different sites in the body (breast and abdominal skin) will respond to growth factors in a similar fashion. Unfortunately, there are very few procedures carried out on legs that would permit excision of normal skin for research purposes.
The results from the microarray analysis point to differential expression of genes involved in the extracellular matrix (ECM), inflammation, and the processes of cellular turnover and adhesion, in LECs from lymphedema skin compared to normal skin.
Nelson et al, 29 in studies on the differential gene expression of primary lymphatic endothelial cells (compared to blood endothelial cells), from normal skin, reported the upregulation of genes linked to calcium signaling (15 genes) and cell adhesion (5 genes). Our microarray results from primary lymphedema skin point to a more prominent role for all these processes, as evidenced by the larger number of KEGG pathway-associated genes involved in calcium signaling and cell adhesion that were upregulated or downregulated. Hypoxia has been shown to alter the adhesive properties of LECs cultured in vitro, 30 by increasing attachment of LECs to the extracellular matrix. Genes involved in the processes of cell adhesion, apoptosis, and chemotaxis have been shown to be differentially upregulated or downregulated by LECs under hypoxic conditions. 30 It may be that hypoxia plays a major role in the clinical manifestations seen in primary lymphedema skin. Differential expression of genes involved in cell turnover, basic metabolism, and the cytoskeleton has been shown to be linked to the propagation of LECs cultured in vitro. 31 This was associated with the downregulation of genes encoding extracellular matrix components. 31 Our results from LECs from primary lymphedema skin favor reduced turnover in the components of the extracellular matrix. TIMP metallopeptidase inhibitor 3, an inhibitor of the matrix metalloproteinases, was upregulated. The matrix metalloproteinases are involved in the degradation of the extracellular matrix. 32 MMP-16 was upregulated 15-fold in LECs from lymphedema skin. Also upregulated was the gene encoding for the major structural protein of the basement membrane, collagen type IV alpha 3 33 and collagen type IV alpha 5. It is tempting to speculate that the combination of reduced breakdown of the extracellular matrix, and increased deposition of certain types of collagen, may in part account for the thick, warty appearance of severe primary lymphedema tissue.
The genes encoding for the interleukins-6, -8, and -32 (IL-6, IL-8, and IL-32) were all downregulated in LECs from primary lymphedema skin. IL-8 is the major interleukin of the inflammatory process 34 and has been implicated in angiogenesis, chemotaxis, and cancer metastasis. 35 In addition, the gene encoding for the interleukin-7 receptor, known to play an important role in the activation of T-cells, B cells, and macrophages in inflammatory diseases like rheumatoid arthritis and psoriasis,36,37 was also downregulated. Interestingly, the cell adhesion molecule, E-selectin, known to play a crucial role in leukocyte recruitment during acute inflammation 38 was also downregulated. E-selectin had been shown to be expressed in a dose dependent manner by cultured human LECs stimulated with lipopolysaccharides. 39 This pattern points towards a general suppression of the genes involved in the acute inflammatory process.
Differential gene expression of primary cultured LECs and blood vascular endothelial cells (BECs) have previously shown the upregulation of pro-inflammatory genes like selectin and chemokine (C-X-C motif) ligand 11. 29 In our study, E-selectin was downregulated 21-fold. A possible explanation for this may be the chronic nature of the inflammatory process occurring in this subset of patients with chronic lower limb and genital swelling. E-selectin is one of the cell adhesion molecules known to be expressed in response to the local release of cytokines.38,39 It may be that the pathological process occurring in this subset of patients with primary lymphedema may be more of a chronic inflammatory process, with the suppression of pro-inflammatory genes. Another possible explanation may be differential patterns of expression of E-selectin in vitro. The expression of E-selectin by lymphatic endothelial cells in vitro has been shown to depend on how the LECs are cultured. 40 Tumor necrosis factor, member 15 (vascular endothelial growth inhibitor), upregulated 17-fold in lymphedema LEC isolates, has been implicated to have an inhibitory effect on cellular motility and adhesion in bladder cancer cells, and is thought to function as a negative regulator of aggressiveness during the development and progression of bladder cancer.41,42 It has also been shown to induce apoptosis in endothelial cells and inhibit tumor neovascularization and tumor progression in cellular and animal models.43,44 Zinc finger protein has been linked to the induction of apoptosis and enhancement of the effect of drugs on hepatocellular carcinoma cells. 45 Its expression has been linked to the suppression of E-cadherin and loss of cell–cell adhesion in human carcinoma cell lines, thereby favoring tumor invasion. 46 Taken together, this pattern of gene expression points towards increased cell turnover, consistent with transcriptomal studies of human dermal LECs propagated in vitro. 31
In summary, this study has isolated and characterized for the first time, LECs from primary lymphedema skin (patients with massive limb and genital swelling). It is also the first time a whole transcriptome study has been performed on human primary lymphedema skin. LECs isolated from primary lymphedema skin respond to the growth factors VEGF-A, VEGF-C, and VEGF-D in a similar fashion to LECs from normal skin. These LECs are also capable of forming microtubules in culture. Transcriptional profiling of these LECs has shown a large number differentially expressed genes. Genes involved in cellular and extracellular matrix turnover and type IV collagen deposition were upregulated. Various pro-inflammatory genes were downregulated. Abnormalities in the basement membrane component, chronic inflammation, and intrinsic defects in extracellular matrix turnover may play a major role in the pathogenesis of primary lymphedema.
Future research work should entail in vivo studies with LECs cultured from primary lymphedema tissue, to see if the results above are replicated under physiological conditions.
Footnotes
Author Disclosure Statement
No competing financial interests exist.