
Abstract
Abstract
Background:
Lymphedema is the progressive distention of tissue due to lymphatic dysfunction. The affected area enlarges over time because of fibroadipose deposition, causing morbidity. Because increasing tissue mass requires neovascularization, we hypothesized that angiogenesis or vasculogenesis might be upregulated in lymphedema.
Methods and Results:
Lymphedematous tissue was collected prospectively from nine patients after resection: upper extremity (n=1), lower extremity (n=3), penis/scrotum (n=5). Neovascularization was compared to normal tissue. Specimens were analyzed using immunohistochemistry for α-smooth muscle actin (pericyte marker), CD31 (microvascular density), CD31/Ki67 (proliferating endothelial cells), and CD34/CD133 (endothelial progenitor cells). Quantitative real-time PCR (qRT-PCR) was used to determine mRNA expression of progenitor cells (CD133) and factors that recruit them: vascular endothelial growth factor-A (VEGF-A), hypoxia-inducible factor 1α (HIF-1α), matrix metalloproteinase-9 (MMP-9), and stromal-cell derived factor 1α (SDF-1α). Angiopoetin-1,-2 (ANG-1,-2), matrix metalloproteinase-2 (MMP-2), and VEGF receptors (VEGFR1,2) were quantified using qRT-PCR. There was no difference in microvascular density, pericytic density, or endothelial proliferation between lymphedematous and normal tissue (p=0.1). Endothelial progenitor cells were not present in lymphedema or normal specimens (p<0.01). VEGF-A (1.3-fold), HIF-1α (0.8-fold), SDF-1α (2.1-fold), VEGFR2 (0.09-fold), and CD133 (0.02-fold) expression were not elevated compared to normal tissue (p=0.1). ANG-1 (5.6-fold), ANG-2 (2.5-fold), MMP-2 (3.9-fold), MMP-9 (33.4-fold), and VEGFR1 (12.8-fold) mRNA was increased in lymphedematous specimens compared to control (p<0.05).
Conclusions:
Lymphedematous tissue does not exhibit upregulation of angiogenesis or vasculogenesis. Neovascularization is unlikely to be involved in the pathogenesis of this disease.
Introduction
The mechanism for soft tissue expansion in lymphedema is unknown. We hypothesized that angiogenesis or vasculogenesis might contribute to the pathophysiology of this condition. Patients with this disease have increased skin temperature and blood flow in the affected limb.3–5 In addition, increasing tissue mass, including adipose tissue, requires neovascularization.6–11 The purpose of this study was to determine if lymphedematous tissue exhibits elevated angiogenesis or vasculogenesis.
Material and Methods
Patients
This study followed the ethical standards of the Committee on Clinical Investigation at Children's Hospital Boston, and in accordance with the Helsinki Declaration of 1975, as revised in 2008. Lymphedematous skin and subcutaneous tissue were collected prospectively from patients undergoing a clinically-indicated procedure. Neovascularization was compared to normal skin and subcutaneous tissue. The effects of age, gender, location, and lymphedema type (primary, secondary) were analyzed.
Immunohistochemistry
Specimens were fixed in 10% formalin, embedded in paraffin or OCT (Sakura Finetek, Torrance, CA), and 5 μm sections were cut. Paraffin slides were de-paraffinized with xylene and rehydrated with ethanol, followed by antigen retrieval in Target Retrieval Solution (DAKO, Carpinteria, CA) using a pressure cooker (Biocare Medical, Concord, CA). Sections were incubated with 3% hydrogen peroxide for 10 min to block endogenous peroxidase activity, followed by 5% normal goat serum (Vector Labs, Burlingame, CA) for 1 h at room temperature. Primary antibodies were applied at 4°C overnight and then incubated with biotinylated secondary antibodies (Vector Labs) for 1 h at room temperature. Avidin/streptavidin-based detection system (Vectastain elite ABC kit, Vector Labs) was used. The slides were developed using 3, 3′-diaminobenzidine (DAB) (Vector Labs) as substrate and counterstained with Mayer's hematoxylin. For double stained frozen sections, Alexa 488 or Alexa 594 labeled secondary antibodies (Invitrogen, Carlsbad, CA) were incubated for 45 min at room temperature.
To define angiogenic or vasculogenic factors in the specimens, the following primary antibodies were used: α-smooth muscle actin (α-SMA), present in pericytes, CD31, an endothelial marker; Ki67, a proliferation antigen; CD34, located on immature endothelial cells (all from DAKO); and CD133, a hematopoietic progenitor cell marker (Cell Signaling, Danvers, MA).
Quantification of immunohistochemistry was determined using image analysis. Pictures were obtained with a Nikon Eclipse E800 microscope (Nikon, Melville, NY) connected to a Spot 3.2 camera (Diagnostic Instruments, Sterling Heights, MI). The specimens were examined using Image-pro Analyzer 6.3 (Media Cybernetics, Silver Spring, MD) and staining was measured as percent of total area. An average of ten random high-powered fields was used to assess each specimen.
Quantitative Real-time Reverse Transcription-PCR (qRT-PCR)
Total RNA was extracted from each specimen using TRIzol (Invitrogen). RNA (0.5–1 μg) underwent reverse-transcription using an iScript cDNA synthesis kit (Bio-Rad, Hercules, CA). The resulting cDNA samples were diluted and analyzed in real-time PCR reactions using SYBR PCR reagents (Bio-Rad) and iQ5 real-time PCR system (Bio-Rad). mRNA levels were normalized by glyceraldehyde 3-phosphate dehydrogenase (GAPDH), and expressed as fold change in comparison to normal tissue. Sequences of primers were: (1) vascular endothelial growth factor-A (VEGF-A), 5′-TACCTCCACCATGCCAAG-3′(sense), 5′-GGTACTCCTGGAAGATGTC-3′(antisense); (2) VEGF receptor 1 (VEGFR1), 5′-GACCTGGAGTTACCCTGATGAAA-3′(sense), 5′-GGCATGGGAATTGCTTTGG-3′(antisense); (3) VEGF receptor 2 (VEGFR2), 5′-CACCACTCAAACGCTGACATGTA-3′(sense), 5′-GCTCGTTGGCGCACTCTT-3′ (antisense); (4) angiopoetin-1 (ANG-1), 5′-TTGATTTAGTACCTGGGTCTC-3′ (sense), 5′CTTGAGAATTACATTGTGGAAAAC-3′ (antisense); (5) angiopoetin-2 (ANG-2), 5′-GTAAGCCTCATTCCCTTCCC-3′ (sense), 5′-TGGGATTTGGTAACCCTTCA-3′ (antisense); (6) matrix metalloproteinase-2 (MMP-2), 5′-ACTTGCAGTACTCCCCATCG-3′ (sense), 5′-TTGACGGTAAGGACGGACTC-3′ (antisense); (7) matrix metalloproteinase-9 (MMP-9), 5′-TGGGATTTGGTAACCCTTCA-3′ (sense), 5′-TGCCCGGACCAAGGATAACAG-3′ (antisense); (8) CD-133, 5′-TCCAAGCCTTAGGAGCATCT-3′(sense), 5′-CCTCATGTTTGGAGGATCTTG-3′(antisense); (9) hypoxia-inducible factor −1α (HIF-1α), 5′-CGTTCCTTCGATCAGTTGTC-3′(sense), 5′-TCAGTGGTGGCAGTGGTAGT-3′ (antisense); (10) stromal cell-derived factor-1α (SDF-1α), 5′-ACTGGGTTTGTGATTGCCTCTGAAG-3′(sense), 5′GGAACCTGAACCCCTGCTGTG-3′(antisense); (11) GAPDH, 5′-GGTGAAGGTCGGAGTCAAC-3′(sense), 5′-CCATGGGTGGAATCATATTG-3′(antisense).
Statistical analysis
Immunohistochemical and qRT-PCR values between lymphedematous and normal tissue were compared by Student's t test or Wilcoxon rank sum test. Spearman's correlation coefficient or Student's t test was used to determine if patient age, gender, or lymphedema type affected the results. One-way ANOVA was utilized to ascertain if disease location influenced the data. Statistical analysis was performed using the SAS software package (version 9.2, SAS Institute Inc., Cary, NC). P values <0.05 were considered significant.
Results
Patients
Nine patients with lymphedema were included in the study (Table 1, Fig. 1). Lymphedematous specimens consisted of: upper extremity (n=1), lower extremity (n=3), penis/scrotum (n=5). Mean patient age at the time of resection was 28.3 years (range 2–51 years).
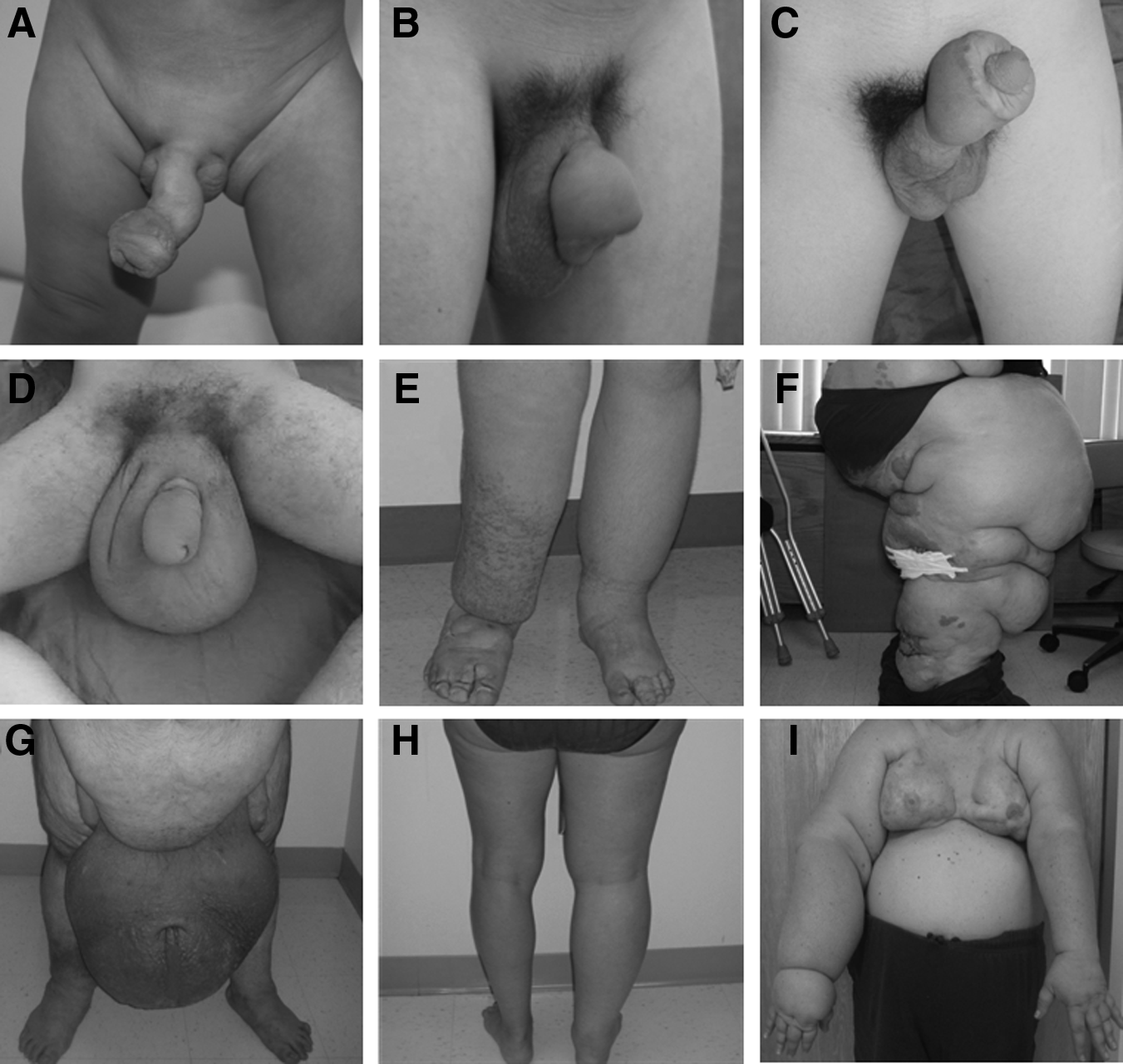
Study cohort.
Neovascularization
Microvascular density (1.3%) and pericytic density (2.7%) were not different compared to control (1.8% and 2.0%, respectively; p=0.1) (Figs. 2 and 3). Experimental and normal tissue did not exhibit endothelial proliferation (Fig. 4). Endothelial progenitor cells were not present in lymphedematous or control samples by CD34/CD133 staining or qRT-PCR for CD133 (0.02-fold; p<0.01) (Figs. 4 and 5). Age (p=0.8), gender (p=0.8), disease location (p=0.9), and lymphedema type (p=0.8) did not affect the results.
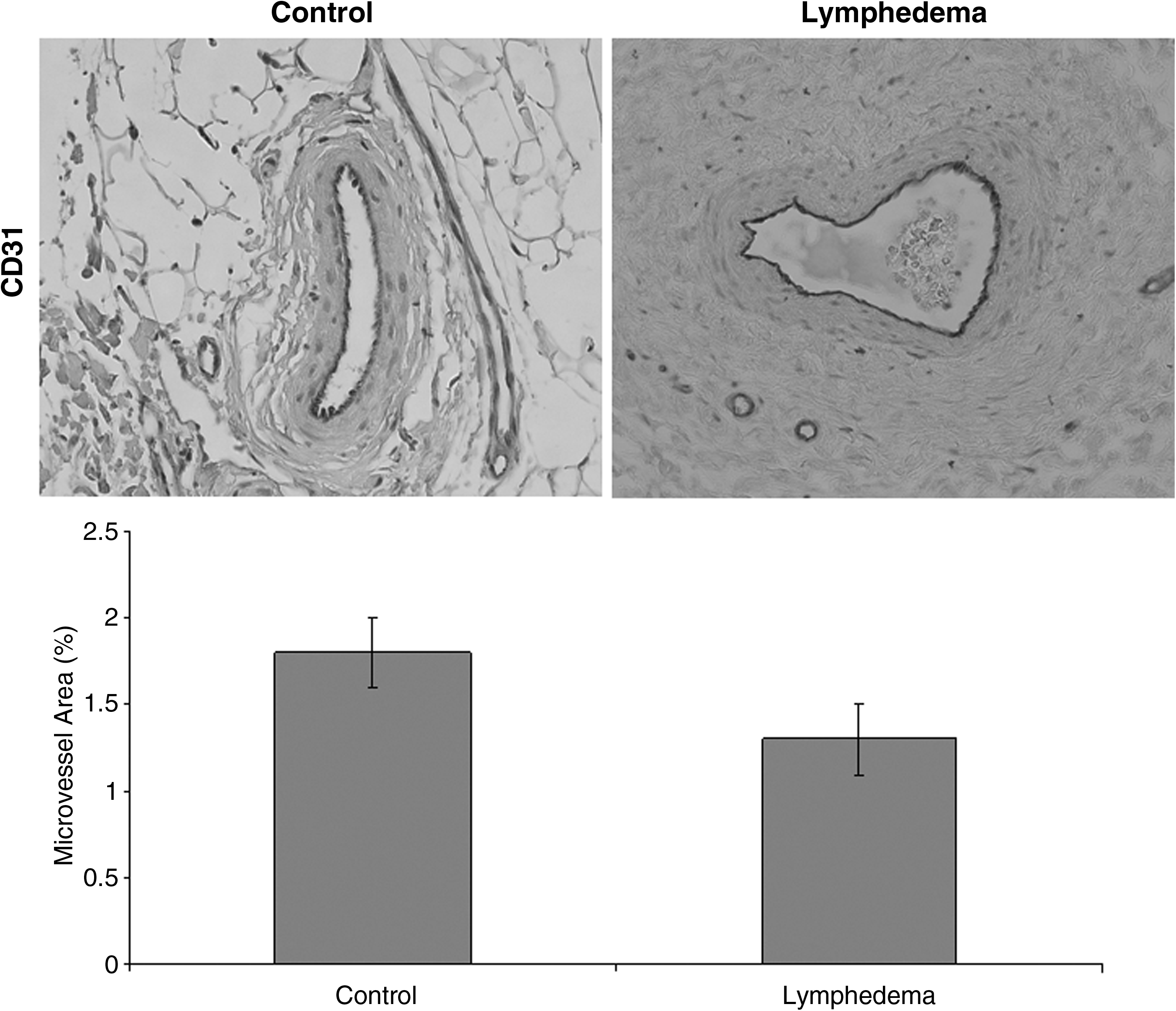
Microvascular density in lymphedema. CD31 immunohistochemical staining shows no difference in vasculature between lymphedema and normal tissue (p=0.1).
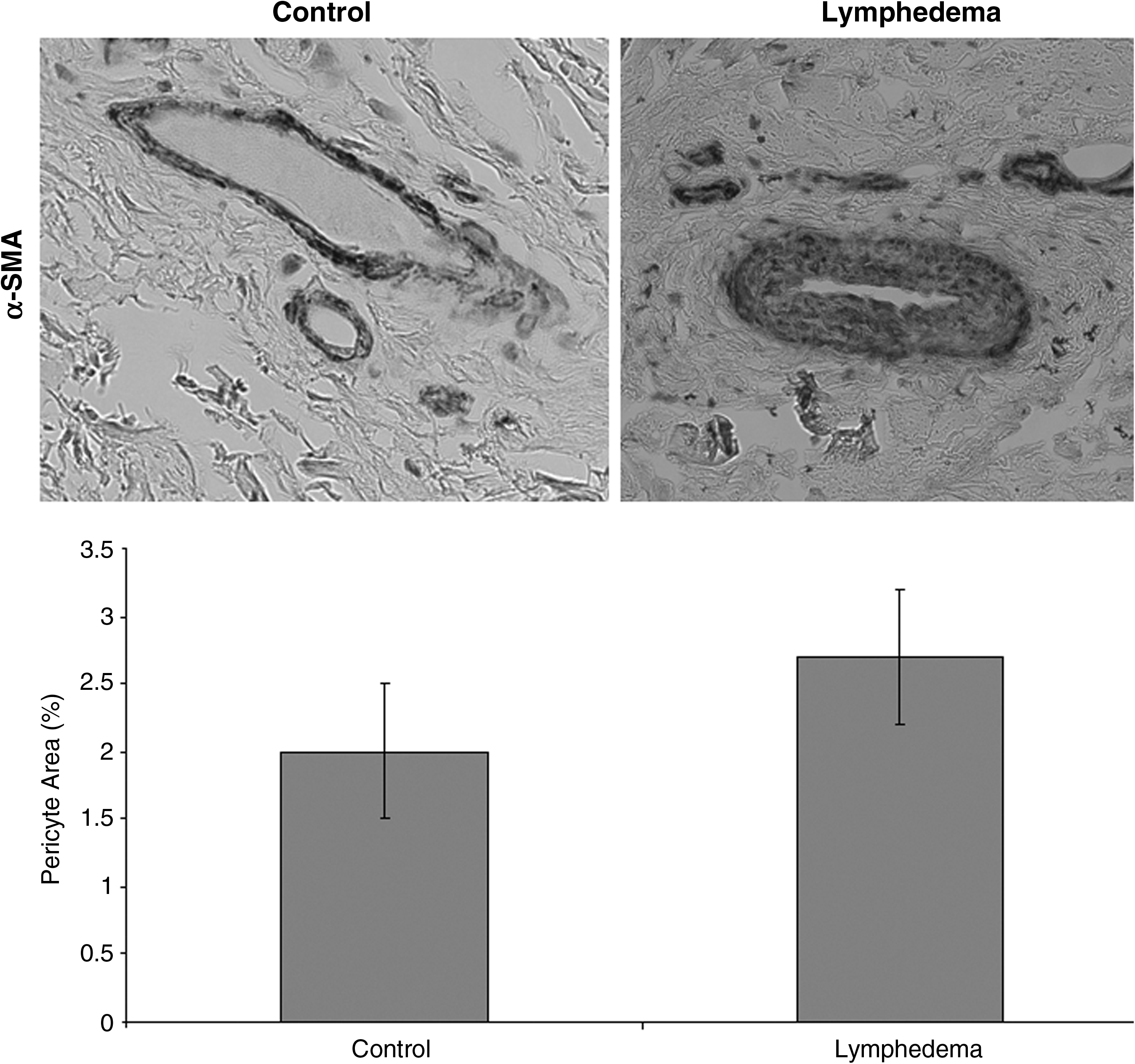
Pericytic density in lymphedema. α-SMA immunohistochemical staining shows similar pericytic density in lymphedematous tissue compared to control (p=0.1).
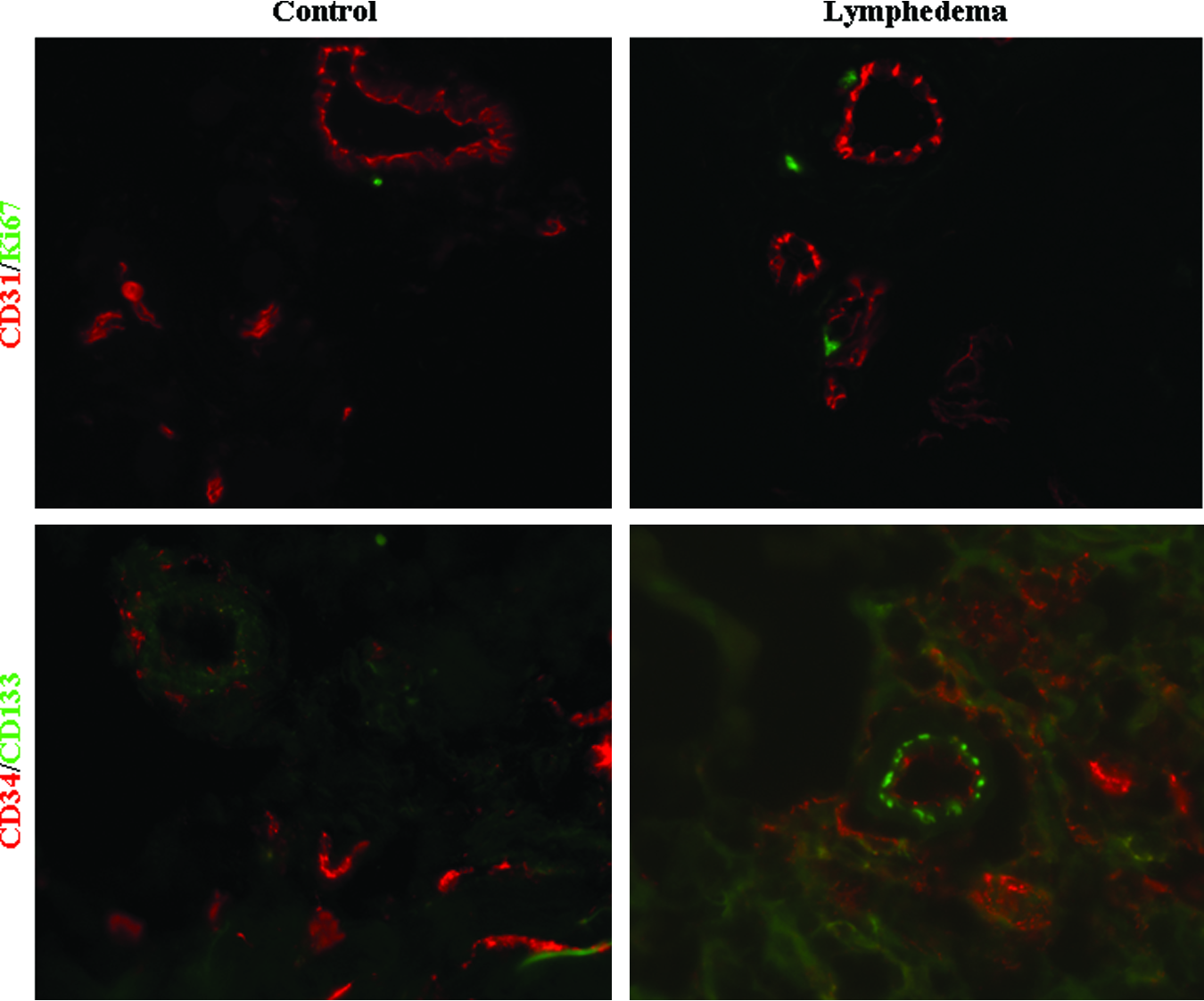
Endothelial proliferation and endothelial progenitor cells (EPCs) in lymphedema. Immunofluorescence with CD31 (red) and Ki-67 (green) antibodies demonstrates absence of proliferating endothelial cells in experimental and control specimens. Using CD34 (red) and CD133 (green) antibodies, no EPCs (yellow) are present. A color version of this figure is in the online article at www.liebertpub.com/lrb
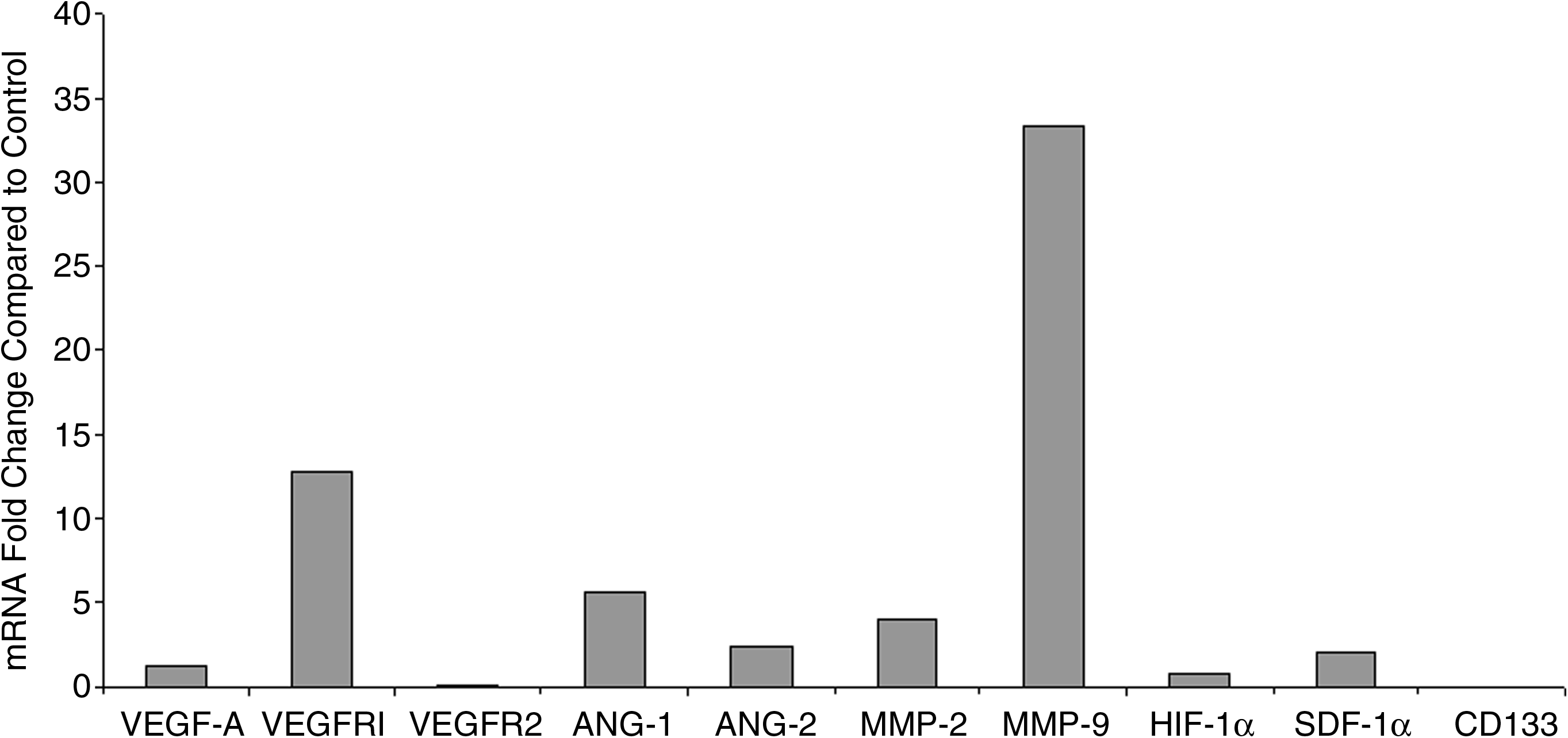
mRNA expression of angiogenic and vasculogenic factors in lymphedematous tissue.
Angiogenic/ vasculogenic factors
VEGF-A expression was not increased in lymphedematous samples (1.3-fold; p=0.2). VEGFR1, an inhibitory decoy receptor, was elevated in lymphedema specimens (12.8-fold; p=0.04); VEGFR2, the biologically active receptor, was not different (0.09-fold). HIF-1α (0.8-fold) and SDF-1α (2.1-fold) mRNA in lymphedematous specimens were similar to normal tissue (p=0.1). ANG-1 (5.6-fold) and ANG-2 (2.5-fold) levels were increased compared to control (p=0.03). MMP-2 and MMP-9 expression also were elevated (3.9-fold; p=0.05 and 33.4-fold; p<0.01, respectively). Expression of angiogenic or vasculogenic factors was not different by patient age (p=0.1), gender (p=0.7), disease location (p=0.9), or lymphedema type (p=0.9).
Discussion
Lymphedema causes morbidity because the subcutaneous layer enlarges over time from adipose deposition. 1 Increasing tissue mass requires neovascularization to support its expansion through angiogenesis (growth of new blood vessels from pre-existing vasculature),6–11 or vasculogenesis (de novo formation of new vasculature).12–16 Nevertheless, we found that angiogenic and vasculogenic factors were not upregulated in lymphedematous tissue; implying that neovascularization is likely not involved in the pathophysiology of this disease.
Lymphedematous tissue had increased expression of VEGFR1, an inhibitory decoy receptor, whereas the level of VEGFR2, the biologically active receptor, was not different than in control tissue. 17 This finding may explain why neovascularization was not upregulated in lymphedematous samples, since increased binding of VEGFR1 to VEGF-A prevents VEGF-A signaling through VEGFR2.
Vasculogenesis is also likely not involved in the pathogenesis of lymphedema. Endothelial progenitor cells were not present in lymphedematous tissue, and factors that recruit them, HIF-1α,18–20 SDF-1 α,21,22 and VEGF-A,23–25 were not elevated.
Normal pericytic density and elevated ANG-1 expression compared to ANG-2 were consistent with a stable vasculature in lymphedema. Pericytes are involved with vessel maturation; their loss is associated with neovascularization.26,27 ANG-1 stabilizes endothelial cells by preventing VEGF expression and the recruitment of pericytes. 28 In contrast, ANG-2 promotes endothelial sprouting by destabilizing endothelial–pericyte contact. 29 ANG-1 may be upregulated in lymphedema to reduce inflammation caused by increased interstitial fluid. 30 ANG-1 protects against inflammation, while ANG-2 induces inflammation and vascular permeability. 31 Because pericytic density in lymphedematous tissue was no different than in control tissue, ANG-1 and ANG-2 expression may be a response to chronic inflammation, rather than promoting neovascularization.
MMP-2 and MMP-9 were elevated in lymphedematous tissue. These proteases are involved in angiogenic, vasculogenic, and inflammatory processes.32,33 MMP-2 and MMP-9 promote angiogenesis by degrading extracellular matrix, releasing pro-angiogenic factors, and facilitating endothelial proliferation. 34 MMP-9 also is pro-vasculogenic; it facilitates endothelial progenitor cell migration from the bone marrow into the circulation.35,36 Absence of neovascularization between lymphedematous and normal tissue suggests that inflammation, instead of neovascularization, is likely responsible for the elevated MMP-2,9 expression. Fibrosis in lymphedema is the result of extracellular remodeling by local inflammation, which is mediated by MMPs.32,33
One limitation of this study is the heterogeneity of the cohort. Patients with primary and secondary lymphedema, affecting different anatomical locations, were included. Control specimens from the contralateral/unaffected area of the same patient were unavailable for comparison. In addition, longitudinal comparison of lymphedematous tissue in the same patient was not possible. Neovascularization might be elevated if assessed in a sample in the same patient prior to progression. These limitations could be minimized if an animal model were available that recapitulated the progressive fibroadipose deposition that occurs in human lymphedema.
Conclusions
Lymphedematous specimens do not exhibit upregulation of angiogenesis or vasculogenesis compared to normal tissue. Neovascularization is likely not involved in the pathogenesis of this condition.
Footnotes
Acknowledgment
We thank David Zurakowski Ph.D. for his statistical expertise.
Author Disclosure Statement
No competing financial interest exists.