
Abstract
Abstract
Background:
Analysis of the innervation pattern of the thoracic duct in young and elderly human subjects has been performed. The subdivision of the vessels in cervical and lumbar region were taken in consideration.
Methods and Results:
Immunostaining for general nerve fibers with a PGP 9.5 marker disclosed a diffuse innervation of the thoracic duct in young subjects, which was strongly reduced in elderly subjects. In young subjects, tyrosine hydroxylase (TH) and neuropeptide Y (NPY) immunoreactive fibers, markers of noradrenergic postganglionic sympathetic fibers, were frequent; choline acetyltransferase (ChAT) immunoreactive fibers, marker of cholinergic parasympathetic nerve fibers, were also well represented. Therefore, the influence of sympathetic and parasympathetic nerve systems on the thoracic duct can be confirmed. The immunoreactivity of vasoactive intestinal peptide (VIP), a neuropeptide frequently present in cholinergic parasympathetic nerve fibers, was scarcely present. Dopamine-positive fibers were observed in few short nerve fibers. Substance P (SP)-positive fibers were widely distributed in the medial and intimal smooth muscle layers, suggesting their involvement as contractile modulating fibers and sensitive fibers. In elderly subjects, an evident reduction of all specific nerve fibers analyzed was detected, the ChAT-positive fibers being the most affected.
Conclusions:
The lymphatic vessel thoracic duct is able to regulate hydrodynamic lymph flow by intrinsic contraction of its smooth muscle layer. Therefore, analysis of the thoracic duct innervation pattern may be important in assessing the regulation of vessel contraction. These findings called attention to the reduction of lymphatic drainage functionality affecting fluid balance in the elderly.
Introduction
We now understand that this phenomenon is due to an intrinsic force consisting of phasic contractions initiated by action potentials originating in pacemaker cells in the muscle layer. 5 Therefore, the smooth muscle layer not only plays a role in the elastic behavior of the wall, significantly affecting passive driving forces, 6 but it also plays an active part in lymph propulsion.7,8
Considerable evidence suggests that lymphatic vessels receive a nerve supply.9–11 Nerve stimulation in vivo was also demonstrated to change lymph flow rate and lymphatic pressure.7,12 Furthermore, several neuropeptides, such as neuropeptide Y (NPY), vasoactive intestinal peptide (VIP), and substance P (SP), were found associated with nerve fibers innervating mesenteric or femoral lymphatic vessel collectors.10,11,13
However, much understanding about lymphatic innervation patterns comes from studies using experimental animal models, 14 and only a few works deal with human specimens. Moreover, little is known about the impact of the aging process on lymphatic innervation.
The purpose of the present study was to investigate the innervation pattern of human thoracic duct. Given the emerging evidence of microanatomic differences between the upper and lower parts of human thoracic duct, 15 we also analyzed the cervical and lumbar regions separately. Moreover, the study included morphological and morphometric comparison of thoracic duct from young and elderly humans.
Materials and Methods
Ethics
All the experiments performed in this work were previously approved by the local Ethical Committee, and were conducted in accordance with the Declaration of Helsinki in its revised edition, the Guidelines of Good Clinical Practice (CPMP/ICH/135/95), and international and local regulatory requirements. To each sample was assigned a progressive number followed by a letter indicative of the participating unit. For each case, a report was prepared, indicating the age and sex of patients as well as general information. The anatomical study units (Camerino, Sapienza, and Novara) knew only the number and letter of each sample. Therefore, a “double blind” procedure was used in all experiments.
Samples
Specimens of thoracic duct were obtained from autopsies on 16 dead subjects: 8 male subjects (age range 28–35 years) were considered for the young group; 8 male subjects (age range 68–75 years) were considered for the elderly group. Specimens of thoracic duct from the cervical region (3 cm from the blood venous stream confluence) and from the lumbar region (3 cm above the cisterna chyli structure) were obtained from each subject. Each sample (1.5 cm in length) was rapidly washed in cold phosphate-buffered saline (PBS), divided into halves, one oriented along its sagittal plane, the other oriented along the coronal plane, embedded in a cryoprotectant medium, and frozen in isopentane cooled with liquid nitrogen. Serial sections, 40 μm thick, were cut on a cryostat at −20°C.
Thick sections were used for better visualization of the total branching of nerve fibers along the vessel walls. In fact, following previous observations performed in our laboratories on lymphatic vessels, we have observed a loss of visualization of the finest fiber branching in sections below ∼45 μm thick. Otherwise, sections thicker than 40–45 μm showed a high background fluorescence signal.
Owing to the considerable thickness of the slides, immunohistochemical experiments on cryostatic slides were performed by the free-floating method.
The first section of each sagittal and coronal specimen was stained for the immunohistochemical detection of nerve fiber profiles using a general marker for nervous tissue, Protein Gene Product 9.5 (anti-PGP 9.5, rabbit polyclonal, diluted 1:600 in PBS; Cambridge Research Biochemicals, UK).
The second section was stained for the immunohistochemical detection of tyrosine hydroxylase (TH) to visualize catecholaminergic fibers (anti-TH, mouse monoclonal, dilute 1:10 in PBS, Novocastra Laboratory, UK).
The third section of each specimen was stained for the immunohistochemical detection of choline acetyltransferase (ChAT) to visualize cholinergic fibers (anti-ChAT, goat polyclonal, diluted 1:30 in PBS, Chemicon, CA).
The fourth section of each specimen were processed for immunohistochemical detection of neuropeptide VIP (rabbit polyclonal, diluted 1:1000 in PBS; Peninsula Laboratories, Chicago, IL).
The fifth section of each specimen were processed for immunohistochemical detection of neuropeptide NPY (rabbit polyclonal, diluted 1:600 in PBS; Cambridge Research Biochemicals).
The sixth section of each specimen was stained for the immunohistochemical detection of neuropeptide SP (rabbit polyclonal, diluted 1:400 in PBS; Cambridge Research Biochemicals).
The seventh section of each specimen was stained for the immunohistochemical detection of dopamine-positive fibers (mouse monoclonal antibody, diluted 1:50; Abcam, code ab8892; Cambridge UK).
To visualize the immunoreaction for VIP, NPY, and NT, a secondary antibody fluorescein isothyocyanate-conjugated antiserum (Vector Laboratories, Burlingam CA) was used. Control sections were processed as above, but using a non-immune rabbit IgG instead of the primary antibody. No positive reaction was observed under these conditions.
The eighth section of coronal specimens only was processed for hematoxylin and eosin staining to assess general microanatomy and to detect morphometric parameters
Vessel and nerve fiber morphometry
Morphometric parameters of cervical and lumbar region of thoracic duct were detected on hematoxylin and eosin staining slides by light microscope connected to a digital camera, using image analysis computer software (Qwin, Leica). Total area and lumen area were delineated on vessel visualization; wall thickness and wall area were calculated from above parameters. Smooth muscle cell number were counted, by the evidence of their nuclei, along 5 randomly chosen wall thickness line, and referred as mean for each slides. From these data, the mean±SEM for each experimental group were calculated. Sagittal sections only were considered for nerve fiber morphometric analysis.
The nerve fiber profiles along the thoracic duct were detected by fluorescence microscope connected to a digital camera, using image analysis computer software (Qwin, Leica).
Analysis protocol was performed at high magnification (65X) to have better visualization of running fibers, and to reduce the dark/light effect of sinusoidal fibers running along the z-axis of slides. Each slide showed a total histological section of ∼170×104 μm2. Five randomly single fields of analysis (each field=150×103 μm2, corresponding to computer visualization) were chosen, for a final total field of analysis of 750×103 μm2. The experimental data (immunopositive nerve profile) were obtained by the sum of the data obtained in the five single fields.
The stereological method for counting fibers was adopted in accordance with the evaluation of length fiber density (Lv) parameter. 16 In each field under analysis, Lv represents the density of nerve profile per unit volume. The volume of analysis consisted in total area of analysis field×slides thickness (300×105 μm3).
A nerve profile was defined as an immunopositive segment seen regardless of its size and length (isotropic distribution). The shift of focus plane was used to better observe the several nerve profiles included in the volume of analysis. Experimental data were obtained using following formula: Lv=Q/ V, where Q represents the number of immunopositive nerve profiles, and V represents the volume of analysis.
No Lv data normalization related to vessels thickness was adopted, because the volume of analysis was constant during data detecting, and the variability of wall thickness was avoid (as detailed above).
Data analysis
Data are presented as mean±S.E.M. calculated from values detected in the individual samples of each experimental group. The normal distribution of nerve fiber data and cell counting data were assessed by means of the Kolgomorov-Smirnov test.
Statistical differences among experimental groups were then assessed by the ANOVA test, followed by the Newman-Keuls post hoc test. Statistical procedures were performed with the Prism 2.01 statistical software (GraphPad Software Inc., CA). The level of significance was p<0.05.
Results
The microanatomical structure of thoracic duct shows a vessel wall composed of an external adventitial layer, a muscular medial layer, and an internal endothelial layer. Cervical region sections displayed a thicker medial layer and smaller lumen diameter in comparison to lumbar section ones (Figs. 1A and 1B).
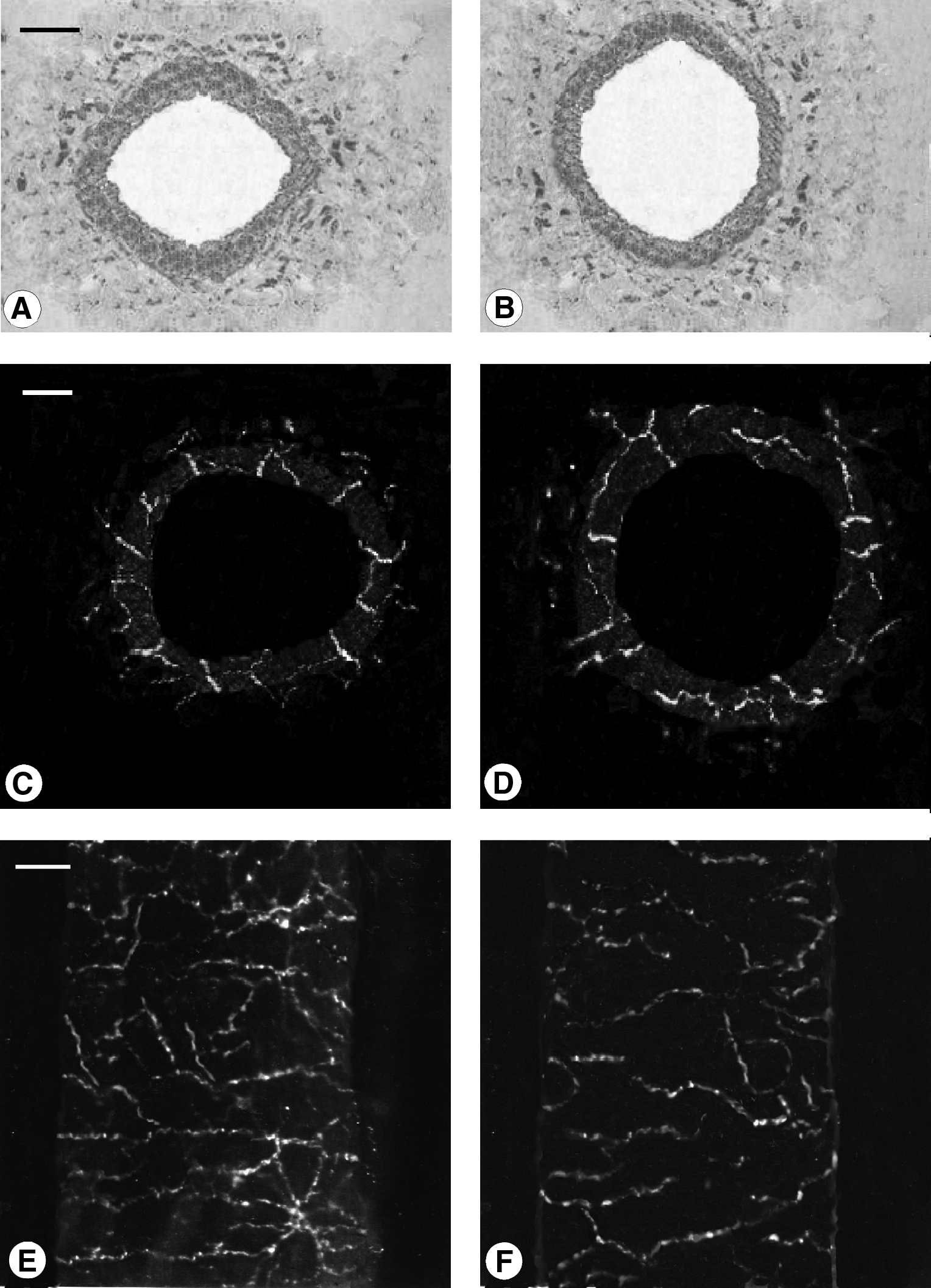
Microphotograph panel showing microanatomical appearance of thoracic duct
Significant differences in morphometric parmeters of the cervical and lumbar regions of the thoracic duct were seen both in young and elderly subjects (Table 1). Furthermore, a significant reduction of wall thickness and wall area in thoracic duct from elderly subjects was detected in comparison to young ones, for both cervical and lumbar regions. The cervical region appeared to be more affected (−23 %) than the lumbar region (−18 %) (Table 1). The number of smooth muscle cells reflect the different thicknesses observed between experimental groups; a lesser number of cells in the elderly thoracic duct has been detected than in young one, both in cervical and in lumbar regions (Table 1).
Data reports as mean±S.E.M. Smooth muscle cells number is referred to vessels thickness (see Materials and Methods section for details). *p<0.05 vs. young; †p<0.05 vs. cervical region.
Immunopositive nerve fibers for neurotransmitter and neuropeptides taken under study were found in all samples from young and elderly subjects. The several immunohistochemical-positive fibers are shown in Figures 1, 2, and 3, while quantitative analysis is seen in Figure 4.
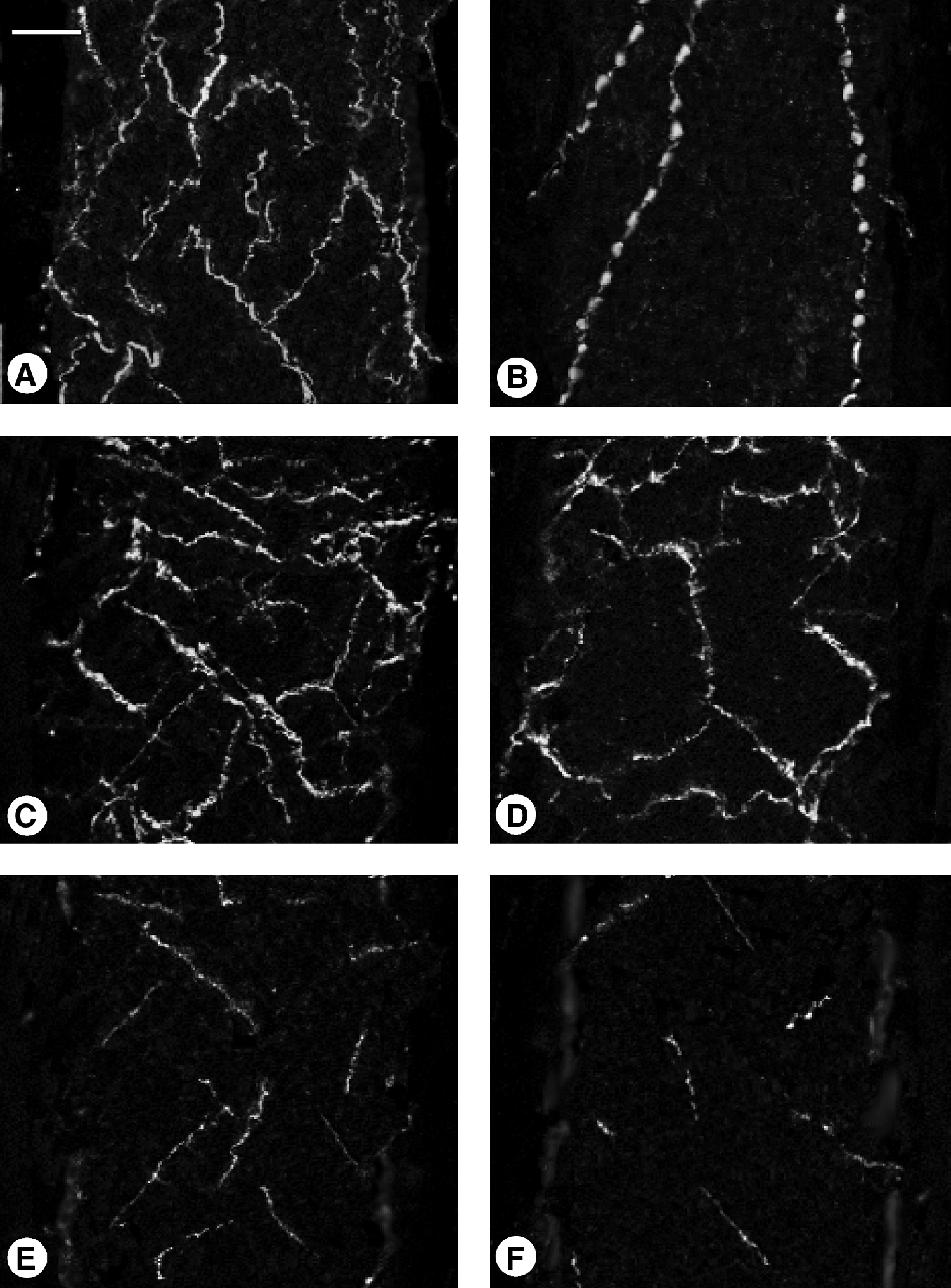
Microphotograph panel showing several immunopositive nerve fibers observed in the thoracic duct of young
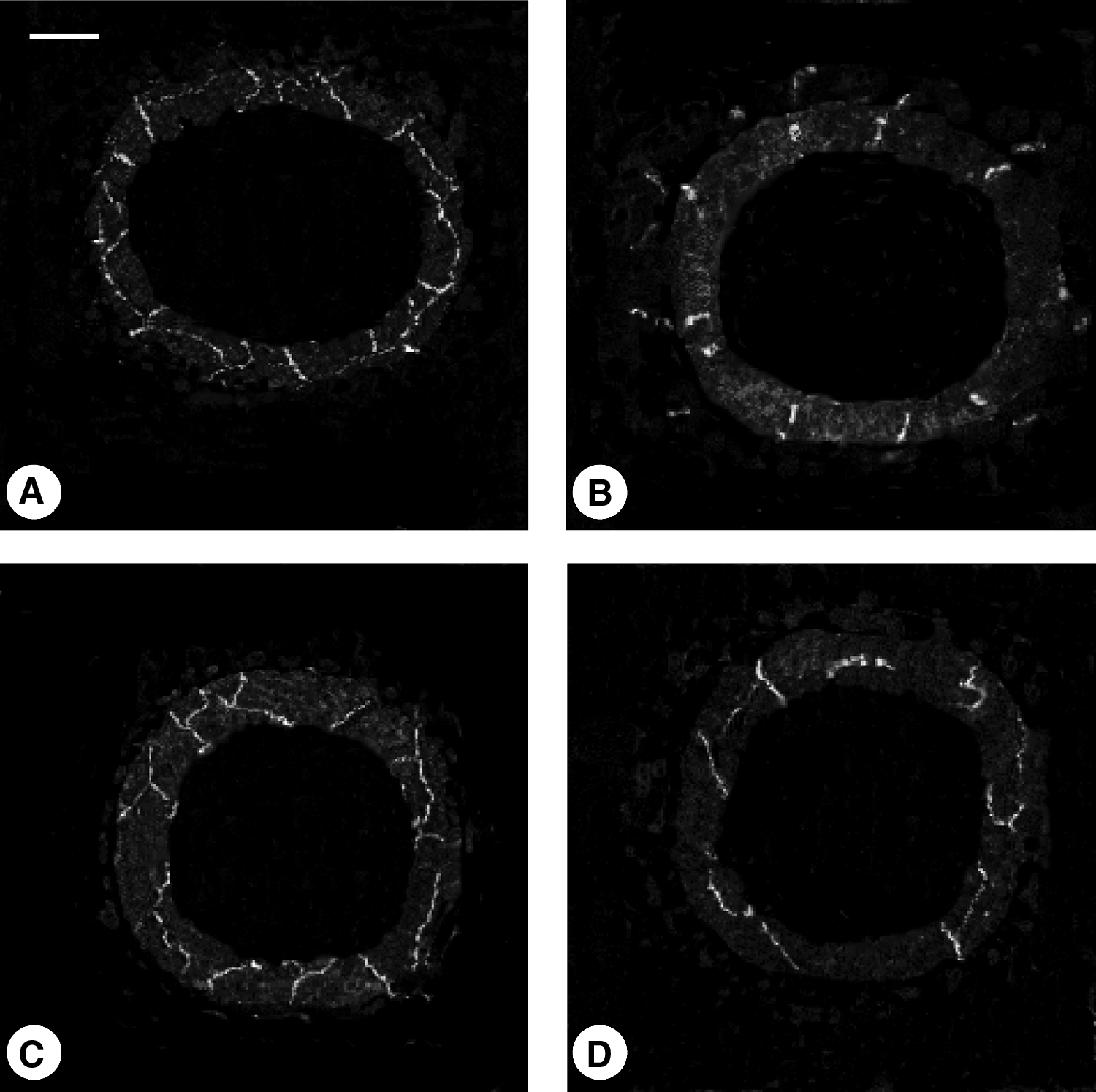
Microphotograph panel showing immunopositive nerve fibers in the coronal sections of thoracic duct in young
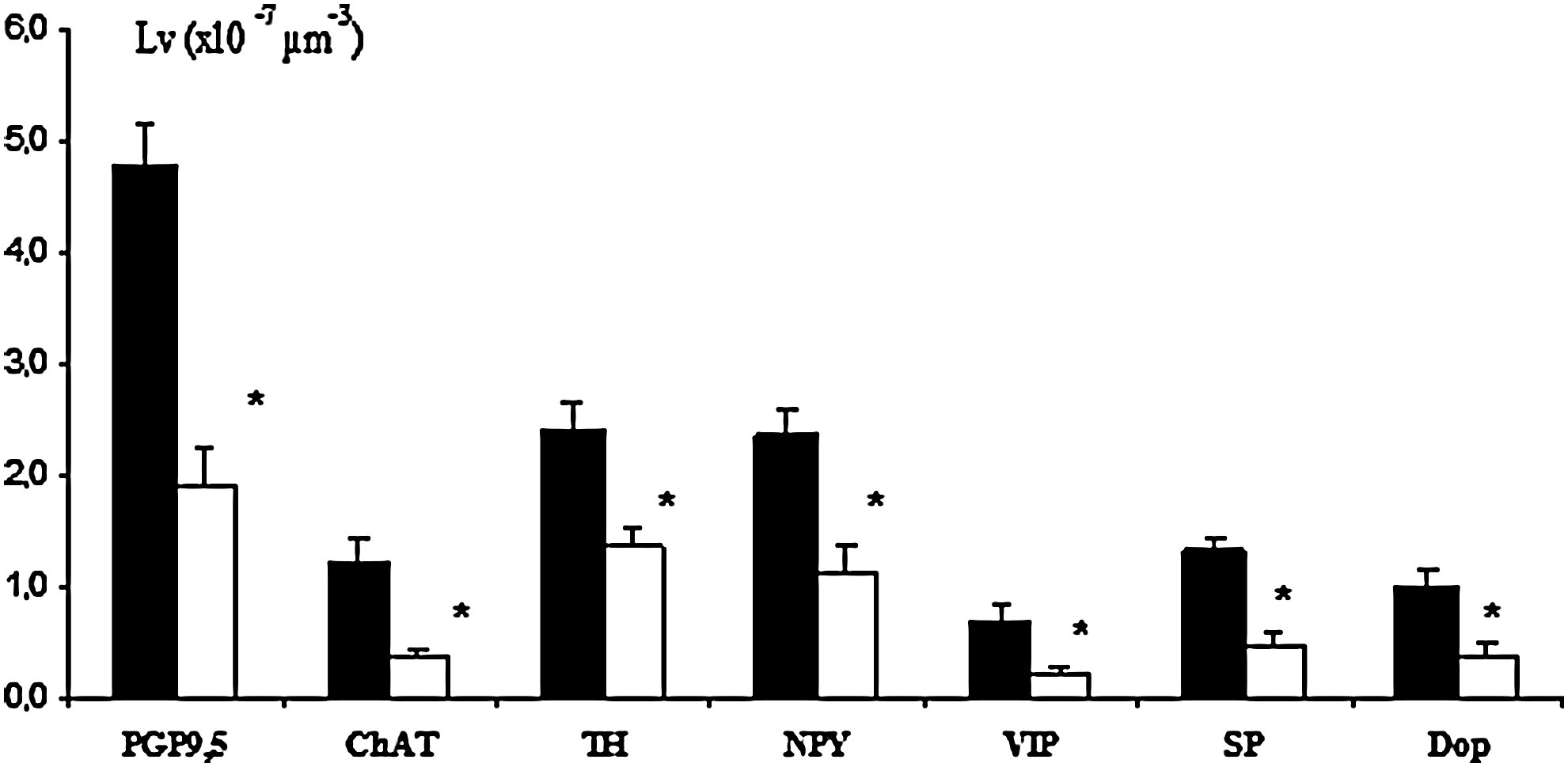
Histogram showing morphometric evaluation of several immunopositive nerve fibers detected in thoracic duct of young (▪) and elderly (□) subjects; *p<0.05 vs. young.
General innervation
Immunostaining for the general neural markers PGP 9.5 showed a rich innervation in coronal and sagittal sections in specimens from young human subjects (Figs. 1C, 1D, and 1E). In coronal sections, PGP 9.5 immunohistochemistry showed immunopositive fibers distributed in the several layers of thoracic duct, giving the impression of nerve fibers running from adventitia and penetrating into the media to reach the endothelial layer. In particular in the medial layer, thick nerve fibers in perpendicular orientation to the longitudinal axis with multiple interconnecting anastomoses of thin nerve fibers were observed (Figs. 1C and 1D). No differences were detected between cervical (Fig. 1C) and lumbar regions (Fig. 1D).
In thoracic duct sections from elderly subjects, an evident and significant reduction of PGP 9.5 immunopositive-nerve fibers was detected (Figs. 1F and 4); no differences were detected among cervical and lumbar part of thoracic duct from elderly subjects.
Specific innervation
In sagittal sections from young subjects following immunohistochemistry for ChAT, NPY, and TH, prevalent longitudinal fibers linking to several transversal anastomosis were displayed (Figs. 2A and 2C), whereas VIP immunostaining was visible with a dotted appearance in a few longitudinal fibers (Fig. 2B), and dopamine-positive fibers displayed a prevalence of longitudinal fibers only rarely linked by anastomoses (Fig. 2E). In specimens from elderly subjects, fibers displayed an arrangement similar to that observed in young samples, but with a lower number of immunopositive fibers (Fig. 2D); VIP was detectable in a lesser number of fibers, while dopamine was detectable in a few short fibers only (Fig. 2F).
In coronal sections from both young and elderly subjects, adventitial immunopositive fibers were observed following TH, ChAT, and VIP immunohistochemistry only (Fig. 3B). In the medial layer, longitudinally and obliquely connecting fibers were noted following TH, ChAT and NPY immunohistochemistry (Fig. 3A), while immunoreactive VIP was observed in perpendicular fibers or dot spot fibers (Fig. 3B). Occasional dot spot immunopositive formation were observed in the endothelial layer following SP immunohistochemistry, suggesting a particular distribution of these fibers (Figs. 3C and 3D). Dopamine immunohistochemistry displayed rare dot spot or very short fibers that did not reach vessel endothelial margins (data not shown).
Morphometrical analysis showed TH- and NPY-immunopositive fibers to be the most represented fibers, while immunopositive-VIP fibers are the least (Fig. 4). In thoracic ducts from elderly subjects, a wide reduction of all immunopositive fibers were detected; TH- and NPY-immunopositive fibers were equally affected in the elderly, and the ChAT-immunopositive fibers appeared the most affected (Fig. 4).
No morphometrical differences in specific innervation patterns were detected among cervical and lumbar region of thoracic duct from young or elderly subjects (data not shown).
Conclusions
Morphometric analysis of the cervical and lumbar regions of the thoracic duct showed morphological differences that may be correlated with different sizes and disposition of smooth muscle cells forming the wall thickness. Indeed, El-Zawahry et al. 15 indicated a different organization of smooth muscle in the abdominal region, where the cells are large and organized in various orientations, while those of the cervical regions are smaller and arranged in a circular fashion. These authors suggest a different action of two thoracic duct regions, where the lumbar ones may be more active in peristalsis, perhaps following a larger size and contractile strength of smooth muscle cells. The morphological variations in the smooth muscle layer may account for the different elastic behavior of the wall, owing to the visco-elastic intrinsic properties of smooth muscle cells. Nevertheless, we did not detect any differences in innervation patterns along the thoracic duct. With this point of view, if differences exist in functional and active regulation of the thoracic duct lymph pump, morphological innervation patterns do not reflect these differences.
The results of this study provide clear immunohistochemical evidence of well-characterized populations of nerve fibers around the thoracic duct.
The influence of the sympathetic and parasympathetic nervous systems on the thoracic duct can be confirmed respectively by clearly evident TH and ChAT immunopositive fibers. In thoracic duct, the sympathetic system is more represented than the parasympathetic one; the parasympathetic system seems to be more affected by aging than the sympathetic system. These findings may indicate that the limited vasodilatory function of ChAT fibers, compared to the effect of vasoconstrictor TH fibers, causes an imbalance between the two systems with consequent impairment of thoracic duct response to lymph flow variation in its passive and active performance.
In visceral peripheral innervation, dopaminergic fibers are widespread and often observed co-localized with NPY-positive fibers. In the thoracic duct, we observed very few dopamine fibers, limited to a restricted portion of the medial layer. The peculiar disposition of dopamine-positive fibers suggests a specialized function, perhaps focused on NPY modulation.
VIP secretion is frequently associated with parasympathetic ChAT fibers, and in the lymphatic system, VIP was observed to act as acetylcholine potentiator and a smooth muscle relaxant. 17 In our study, we found few VIP-positive fibers in the thoracic duct, and comparison between the number of ChAT fibers and VIP fibers suggests that only part of the ChAT fibers might express VIP. These findings lead us to hypothesize that in thoracic duct, VIP may be a minor modulation of the parasympathetic component.
Substance P-immunoreactive fibers were found well represented in the medial layer of thoracic duct running along longitudinal fibers. In fact, since SP has been observed to improve pump efficiency in rat lymphatic muscle, 18 we may hypothesize a similar effect in humans as well. Interestingly, we observed SP-positive fibers frequently reaching the intimal layer of thoracic duct. These findings are well in accordance with previous work which reported, using immunohistochemical and electron microscopy, SP-positive fibers in the subendothelial layer. 19
We hypothesize a dual role for SP-positive fibers in which SP-positive fibers branching in the medial layer may exert modulatory control on smooth muscle activity, while subendothelial SP-positive fibers may act as sensitive fibers. With this point of view, the co-localization of SP in motor parasympathetic ChAT fibers may be an interesting hypothesis.
PGP 9.5 is a soluble protein present in nervous and neuroendocrine cells. It has been identified as ubiquitin carboxyl-terminal hydrolase and has been observed involved in axonal transport physiology. 20 In our work, the widespread reduction of PGP 9.5 immunoreactivity observed in thoracic duct from elderly subjects, leads us to hypothesize a functional default of nerve fibers that is not necessarily followed by an anatomical disappeared of fibers themselves. Moreover, the reduction of specific nerve fibers should be interpreted as a consequence of general nerve fibers disappearing rather than a specific involvement of neuro-signaling factors analyzed.
In conclusion, we have observed in human thoracic duct a widespread presence of different types of nerve fibers, indicating a high modulatory level of thoracic duct hydrodynamics. With aging, the broad reduction of nerve fibers in the elderly may imply the loss of functionality in lymphatic drainage with a consequent effect on fluid balance, and also a limitation in circulation of immune cells throughout the body.
Footnotes
Author Disclosure Statement
No competing financial interests exist for any author.