
Abstract
Abstract
The systematic study of focused animal models has produced an explosion of information regarding the mechanisms governing lymphatic development and the diseases associated with lymphatic dysfunction. Nevertheless, the pathogenesis of systemic lymphangiomatosis has, thus far, eluded mechanistic comprehension. In this review, recent molecular advances in lymphatic vascular development are considered within the context of the animal models that have produced evolving insights. The emerging role of the zebrafish within lymphatic investigation is discussed. Specific models of the human disease pathology are considered in detail. While much has been learned about the molecular framework that surrounds normal lymphatic vascular development, the defect responsible for systemic lymphangiomatosis remains elusive. Development of more robust, recapitulative models will also be invaluable to investigate new and emerging therapeutics for the often devastating disease of systemic lymphangiomatosis.
Introduction
The systematic study of focused animal models has produced an explosion of information regarding the mechanisms governing lymphatic development and the diseases associated with lymphatic dysfunction. Animal studies have led to a new molecular model of embryonic lymphatic vascular development, and have, specifically, provided insight into the pathophysiology of lymphatic insufficiency in both heritable and acquired forms. 2 Thus, it is certainly relevant to address the potential for the mechanistic study of model systems to shed light upon the patheogenesis, natural history, and treatment options for systemic lymphangiomatosis, which, as a first approximation, can be conceptually considered to be the pathological result of a disordering of the normal mechanisms of lymphatic vascular development.
Genetic Models of Lymphatic Development
Comprehension of the molecular mechanisms that serve embryonic lymphatic development has been greatly facilitated by the generation of genetic animal models that target the specific genes proven to be integral to lymphatic structure and function. These models have shed light on the mechanisms of heritable lymphatic insufficiency and other pathologic manifestations of lymphatic dysfunction. Studies involving these animal models hold promise for the development of targeted therapies for the treatment of lymphatic disease.
A large and growing list of molecular signaling agents has now been identified as contributors to, and regulators of, the process lymphatic vascular development. VEGF-C and VEGF-D, originally cloned as ligands for VEGFR-3, were the first factors identified with the ability to induce growth of new lymphatic vessels.3,4 Both VEGF-C and VEGF-D promote, through activation of VEGFR-3, the proliferation of cultured human lymphatic endothelial cells (LECs). 5 VEGF-C and D, through VEGFR-3, are also able to induce lymphangiogenesis in vivo, demonstrable in the skin of transgenic mice that overexpress VEGF-C/D. 6
VEGF-C knockout mice demonstrate complete abrogation of early lymphatic vessel formation, 7 thus underscoring the vital role that VEGF-C plays during embryonic lymphangiogenesis. 8 Although PROX-1 expression is not inhibited in these VEGF-C- deficient mice, PROX-1-positive, lymphatically specified endothelial cells are unable to bud from the embryonic cardinal vein to form initial lymph sacs, suggesting that LEC specification and subsequent cell migration are controlled by distinct signaling pathways. 9 This sprouting defect is rescued by administration of VEGF-C or VEGF-D, but not VEGF-A, indicating specificity for VEGFR-3 and the necessity of VEGF-3-mediated signaling for initial lymphatic vessel formation. 7
In conjunction with the VEGF family, the angiopoietin signaling system appears to modulate both angiogenesis and lymphangiogenesis, 10 and offers another target for genetic manipulation in the study of lymphatic disease. Angiopoietin-2 (ANG-2) knockout mice, in addition to having defective postnatal angiogenesis, exhibit gross abnormalities of the lymphatic system. 11 ANG-2-deficient mice display a phenotype characterized by structurally irregular and leaky lymphatic vessels, chylous ascites, lymphedema, and early mortality. There is evidence that, in the context of lymphatic development, angiopoietin-1 (ANG-1) and ANG-2 cooperatively induce lymphangiogenesis as agonists of TIE-2 receptor.
In contrast, tissues obtained from mice that overexpress ANG-1 are characterized by lymphatic vessel endothelial proliferation, vessel enlargement, and new vessel sprouting, as observed in mice overexpressing VEGF-C. These effects are associated with upregulation of VEGFR-3 and inhibited by systemic blocking of VEGFR-3 signaling, reinforcing the concept that the angiopoietin and VEGF families collaborate during lymphatic vascular development.
With the exception of the VEGF family, perhaps no other molecular regulator plays as a pivotal role in lymphatic development as PROX-1, a homeodomain transcription factor that serves as a master regulator of cell fate decisions leading to specific cell lineages in numerous developing organs. 12 Studies of PROX-1 knockout mice have led to the current molecular model for early lymphatic vascular development in which PROX-1 induces lymphatic vascular development from the preexisting embryonic venous anlage. 13 Indeed, the evidence suggests that PROX-1 specifies LEC fate by directly reprogramming the transcriptome of embryonic venous endothelial cells.
PROX-1 plays a vital role in specifying the lymphatic fate of a subset of embryonic cardinal vein endothelial cells by inducing expression of lymphatic-specific genes in these precursor LECs. The mucin-type transmembrane glycoprotein podoplanin/T1α is one such key PROX-1-induced gene. 14 In the vascular system, podoplanin expression is first seen at E11.0 in budding PROX-1-positive lymphatic progenitor cells and later becomes specifically restricted to lymphatic endothelium.
Podoplanin-knockout mice exhibit cutaneous lymphedema associated with impaired lymphatic transport, dilated lymphatic vessels of the intestine and skin, and neonatal lethality due to respiratory failure. 14 Comparative analysis of wild-type, heterozygous, and knockout animals reveals graded hyperplasticity of lymphatic vessels and defective lymphatic capillary plexus formation. It has been proposed that this molecule is responsible for outlining the superficial capillary beds that anastomose with deeper lymphatic networks in subcutaneous tissue; in vitro studies of cultured endothelial cells indicate that podoplanin may play an important role in LEC migration, adhesion, and tube formation.
The specific expression of Prox1 in the cardinal vein and the Prox1-dependent differentiation of LECs are induced by the transcription factor sex-determining region Y box 18 (Sox18). 15 In addition to Sox18, the orphan nuclear receptor chicken ovalbumin upstream promoter transcription factor II (COUP-TFII), plays a vital role in the initiation and early maintenance of Prox1 expression. 16
Comparative transcriptional profiling of human lymphatic and blood vascular endothelial cells demonstrates remarkable differences in the expression of pro-inflammatory cytokines and their receptors, in addition to proteins involved in cytoskeletal and cell–cell interactions, such as the integrins. 17 As an example, integrin α9, is expressed primarily in LECs, while integrin α5 appears to be blood vascular endothelial–cell specific. 18
Importantly, the integrin α9 subunit appears to be important in the context of lymphatic development. Transgenic PROX-1 overexpression in human umbilical vein endothelial cells (HUVECs), along with the expected upregulation of lymphatic specific markers (including VEGFR-3 and podoplanin) also leads to increased integrin α9 expression, suggesting that integrin α9 upregulation play a role in the program of LEC differentiation. 19 Conversely, knockdown of PROX-1 expression in human LECs leads to reduced integrin α9 expression and a decreased chemotactic response to VEGF-C. These results suggest that integrin α9 may function as a key regulator of lymphangiogenesis acting downstream of PROX-1.
Integrin α9 subunit knockout mice provide the strongest evidence that integrin α9β1 plays a unique and non-redundant in vivo role in lymphatic development. 20 These animals appear normal at birth but later develop fatal respiratory failure from bilateral chylothorax, accompanied by edema and perilymphatic lymphocytic infiltration in the chest wall Although the mechanisms by which integrin α9β1 mediates lymphatic development require elucidation, interactions between integrin α9β1 and VEGF-C/D and/or VEGFR-3 may contribute to the phenotype.
Comparative transcriptional profiling has revealed that hepatocyte growth factor receptor (HGF-R), such as integrin α9, is more highly expressed by cultured LECs than by blood endothelial cells. HGF-R, a transmembrane tyrosine kinase whose expression is increased in sites of tumor growth and metastases, serves as a receptor for HGF, a heparin-binding glycoprotein expressed by cells of mesenchymal origin. HGF was originally identified as a mediator of liver generation and also appears to play a major role in tissue repair and promoting tumor invasiveness.
A recent study in transgenic hepatocyte growth factor (HGF)-overexpressing mice has demonstrated that HGF is also a potent lymphangiogenic factor in vivo. 21 These HGF-overexpressing mice manifest an increased number and enlargement of lymphatic vessels in various organ systems, including the skin and gastrointestinal tract. HGF-induced lymphangiogenesis does not require VEGFR-3 signaling. Systemic blockade of the HGF receptor inhibits cutaneous lymphatic vessel enlargement in a well-characterized mouse model of experimental skin inflammation. 21 Thus, the HGF receptor could serve as a potential target for inhibiting pathologic lymphangiogenesis, particularly since the receptor appears to be preferentially expressed by activated, proliferating lymphatic endothelium, but not by quiescent lymphatic vessels in normal skin. 21
Ephrin-B2 is another protein not originally known to have importance to the lymphatic system, but which has recently been identified to have an essential role in lymphatic development. A knock-in mouse model that expresses a hypomorphic ephrin-B2 gene but lacks its C-terminal PDZ interaction site is characterized by the postnatal development of chylothorax and gross defects of the lymphatic system, including hyperplasia and lack of luminal valve formation in the collecting lymphatic vessels. 22 In addition, while dermal lymphatics appear normal at birth, these mice fail to remodel their primary lymphatic capillary plexus into a hierarchical vessel network.
The Human Disease
Recent advances and accrued molecular insights are noteworthy and laudable, yet the existing model systems, chiefly devoted to altered expression of the known molecular mediators, have not successfully reproduced the pathological manifestations of systemic lymphangiomatosis.
Lymphangioma, as an isolated lesion, is a congenital lymphatic malformation that arises during embryologic development. Lymphangiomata may arise from segments of lymphatic vascular tissue that fail to anastomose properly, or they may represent portions of lymph sacs that become grouped together during development. 23 Lymphangiomatosis, therefore, represents the presence of multiple or widespread lymphatic vascular malformations of this type. The lesions are classified by size and depth of formation. Lymphangiomatosis describes the presence of multiple lymphangiomas. It is frequently associated with other lymphatic related abnormalities and usually involves multiple viscera. Histologically, these lesions are composed of an large numbers of dilated vascular channels, lined by lymphatic endothelium. 24 The cystic spaces are filled with proteinaceous lymph fluid that is free of erythrocytes. The connective tissue stroma consists of varying amounts of spindle-shaped smooth muscle cells, collagen bundles, fibroblasts, and lymphocytes and, often, benign lymphoid aggregates. 23 The cellular components are generally well-differentiated and lack cytologic atypia.
Lessons Learned from Zebrafish
In light of the putative mechanisms that may surround the formation of lymphangiomatous lesions, it would appear prudent to consider the normal processes that invest lumen formation of developing lymphatic structures. Studies of the process of lumen formation (luminogenesis) have been undertaken within in vitro models of tubulogenesis25,26 and much progress has been achieved though studies of the zebrafish vascular system.27,28 The zebrafish (Danio rerio) has only recently been recognized to have a functional lymphatic system and, thereby, has been identified as a model organism for the study of lymphatic vascular development and developmental pathology. 29 During zebrafish embryogenesis, independent cell clusters contribute to the development of a lymphatic trunk that is morphologically distinct from the main blood vessels, does not contain blood cells, and which appears capable of transporting interstitial fluid. This trunk subsequently sprouts to give rise to a lymphatic tree. Lymphatic development in the zebrafish does appear to depend on VEGF-C and VEGFR-3 signaling. 29 Further study of lymphatic development in the zebrafish may, therefore, represent a very rewarding model in which to gain future insights into the pathogenesis of lymphangiomatosis.
A Lymphangioma-Specific Mouse Model
There exists, to date, one murine model that seeks to recapitulate the histopathological described in the human lymphangioma lesion. 30 The model consists of the creation of lymphatic cell hyperplasia induced by intraperitoneal injection of incomplete Freund's adjuvant.30–32 White masses of lymphangiomata appear on the surface of the diaphragm within 1 month after the first injection (Fig. 1). Lymphatic vessels are identifiable with hematoxylin-eosin staining, and demonstrate characteristic LYVE-1 positivity. Within the histological specimens, various degrees of cell fusion can be observed, leading to a progressive increase of vessel-like lumen size (Fig. 1). 32
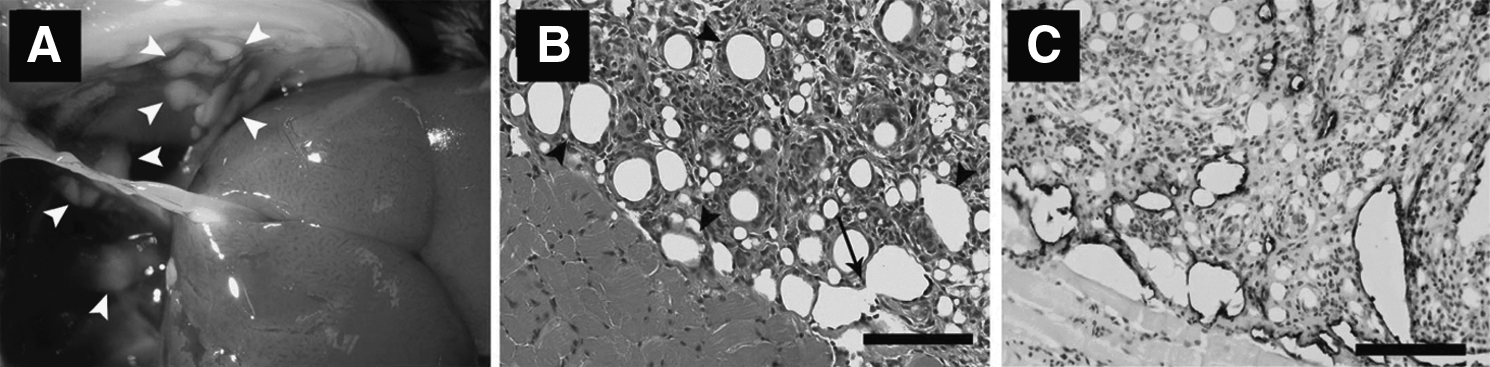
Mouse lymphangiomata induced by intraperitoneal injection of incomplete Freund's adjuvant.
Additional study of these lesions with transmission electron microscopy suggests that the establishment of intercellular spaces leads to the development of tubular structures (Fig. 2). The LECs were noted to align within cords that displayed a thin and irregular lumen. Matrix fragments were detectable in the interendothelial gaps and in the lumina of newly-formed vessels. 32
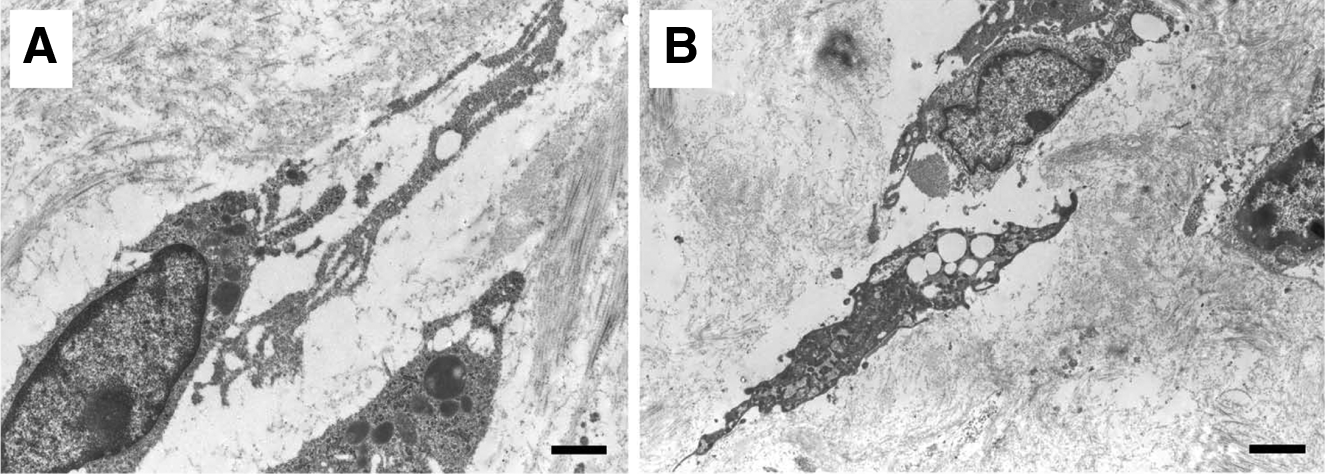
These observations are in general agreement with the phenomena that are observed within the zebrafish. The observations suggest that, in this pathological form of lymphatic vessel formation, there is obligatory alignment of LECs, thereby driving a tunnel through extracellular matrix. During lymphangiogenesis, cords of cells create an extracellular space by the degradation of collagen fibrils; this occurs both within and outside the cell. Sprouting LECs are characterized by the extension of long thin vacuolized processes which probe the extracellular environment and connect with adjacent cells, resulting in the formation of cord-like structures and pre-lymphatic vessels; an intense intracellular vacuolization associated with vesicle coalescence, leading to an intracellular luminal space; and remodeling of matrix, generating space between cells, promoting cell migration and contributing to lumen formation.
Conclusion
The recent observations derived from the experimental lymphangioma-specific murine model lesion are certainly elucidative and suggest that that there is more to be learned mechanistically from this approach. Nevertheless, one must also acknowledge that the model only weakly recapitulates the full systemic expression of the human disease, suggesting that continued, diligent search for new, applicable model systems is still necessary. While much has been learned about the molecular framework that surrounds normal lymphatic vascular development, the defect responsible for systemic lymphangiomatosis remains elusive. It is possible that continued study of the zebrafish development will uncover the mechanistic key. Development of more robust, recapitulative models will also be invaluable to investigate new and emerging therapeutics for this often devastating disease.