
Abstract
Abstract
Background:
We have previously shown that aging is associated with weakened rat mesenteric lymphatic vessel (MLV) contractility. However, the specific mechanisms contributing to this aging-associated contractile degeneration remain unknown. Aging is often associated with elevations in oxidative stress, and reactive oxygen species (ROS) have been shown to reduce the contractility of MLV. Thus in the present study, we sought to assess whether aging is associated with increased levels of oxidative stress and oxidative damage in MLV.
Methods and Results:
MLV were isolated from 9-mo- and 24-mo-old Fischer-344 rats and subjected to the following experimental techniques: measurement of total superoxide dismutase (SOD) activity; estimation of lipid peroxidation levels via measurement of thiobarbituric acid reactive substances (TBARS); detection of superoxide and mitochondrial ROS in live MLV; Western blot analysis, and immunohistochemical labeling of the SOD isoforms and nitro-tyrosine proteins. We found that aging is associated with increased levels of cellular superoxide and mitochondrial ROS concomitant with a reduction in Cu/Zn-SOD protein expression and total SOD enzymatic activity in MLV. This increase in oxidative stress and decrease in antioxidant activity was associated with evidence of increased lipid (as indicated by TBARS) and protein (as indicated by nitro-tyrosine labeling) oxidative damage.
Conclusions:
Thus for the first time, we demonstrate that aging-associated increases in oxidative stress and oxidative damage is indeed present in the walls of MLV and may contribute to the aging-associated lymphatic pump dysfunction we previously reported.
Introduction
The free radical theory of aging suggests that there is a progressive reduction in NO production and a simultaneous increase in free radical production leading to endothelial dysfunction. Beckman and Ames suggested that oxidative stress is an important factor contributing to vascular dysfunction with aging. 20 Vascular aging is associated with both structural and functional changes that can take place at the level of the endothelium, smooth muscle cells, and the extracellular matrix of vessels. Aging is also correlated with increased oxidative stress and oxidative damage, and the endothelium appears to be an important source of superoxide anion (O2-) in the vascular wall. Marin and Rodriguez-Martinez reported that endothelial cells are vulnerable to oxidative stress due to their low antioxidant capacity. 21 Aging-related endothelial dysfunction may involve mechanisms such as alterations in the antioxidant defense systems, increased oxidative injury, or both. Studies suggest that inactivation of nitric oxide by superoxide contributes to impaired vascular function.22–25 NO reacts with superoxide radical (O2-) to form peroxynitrite (ONOO-), which can further induce protein modification and DNA damage in the microvascular system. 26 Thus, decrease in NO bioavailability as a result of excess O2- formation is a major cause of endothelial dysfunction in aging. Aging-associated elevations in oxidative stress may be related to alterations in antioxidant defense enzymes such as the superoxide dismutase (SOD) isoforms Cu/Zn-SOD (located in the cytoplasm), Mn-SOD (located in the mitochondria), and extracellular SOD (EC-SOD). Additionally, the oxidative stress may come from increased production of reactive oxygen species (ROS) via mitochondrial dysfunction, activation of NADPH oxidase, or uncoupling of nitric oxide synthase (NOS).27–29 The balance between the levels of free radical production and cellular antioxidant activity determine the oxidative stress on the tissue and subsequent degree of oxidative damage. Importantly, Zawieja et al. 10 reported that oxygen radicals significantly inhibited contractile activity of rat mesenteric lymphatic vessels, thus we propose that aging-associated increases in oxidative stress could contribute to the decline in contractile activity seen in aged lymphatic vessels. Currently, there are no investigations of aging-associated oxidative stress or oxidative damage in lymphatic vessels. Thus, the goal of the present study is to examine the aging-induced changes in expression and activity of the major cellular antioxidant enzyme, superoxide dismutase (Cu/Zn-SOD, EC-SOD, and Mn-SOD isoforms) while also evaluating peroxynitrite-mediated cellular damage and mitochondria-related superoxide radical production in aged mesenteric lymphatic vessels from Fischer-344 rats in comparison with their adult counterparts.
Materials and Methods
Animals and surgery
For the current studies, we used 9- and 24-month-old male Fischer-344 (F-344) rats obtained from the aged rat colony maintained by the NIH National Institute of Aging. This is a commonly used rat strain in aging-related research,30,31 and the age groups selected represent adulthood and senility, respectively. All animal procedures for current studies were reviewed and approved by our Institutional Animal Care and Use Committee.
Isolation of the mesenteric lymphatic vessels
To isolate mesenteric lymphatic vessels (MLV), rats were anesthetized with a combination solution of 0.4 mg/ml fentanyl and 20 mg/ml droperidol (0.3 ml/kg IM) and diazepam (2.5 mg/kg IM). A 4-cm long midline abdominal incision was made through the skin, underlying fascia, and muscle layers. A small loop of intestine 6–7 cm in length was exteriorized through the incision. A section of mesentery containing lymphatic vessels was positioned in a custom dissection chamber within the field of view of a stereomicroscope and continuously suffused with standard physiological salt solution (PSS) (in mM: 145.0 NaCl, 4.7 KCl, 2.0 CaCl2, 1.2 MgSO4, 1.2 NaH2PO4, 5.0 dextrose, 2.0 sodium pyruvate, 0.02 EDTA, 3.0 MOPS) with pH adjusted to 7.36 at 38°C. Suitable mesenteric lymphatic vessels were identified and cleared of all surrounding tissue. Sections of mesenteric lymphatic vessels 1–3 cm in length were carefully dissected and used for subsequent analyzes. After isolating mesenteric lymphatic vessels, the rat was euthanized with pentobarbital (120 mg/kg body weight IC).
Measurement of total SOD activity
The total activity of SOD in 9-mo and 24-mo-old MLV was measured using the Superoxide Dismutase Assay Kit (Trevigen, Inc., Gaithersburg, MD, Cat. # 7500-100-K) according to manufacturer's instructions. Each sample contained approximately 7–10 isolated MLV pooled in phosphate buffered saline (PBS) (EMD Chemicals, Gibbstown, NJ, Cat. # 6505), and stored in liquid nitrogen at −80°C. MLV samples were homogenized with PBS, and 20 μl of supernatant per sample was used for the SOD activity assay. SOD activity in homogenates was determined by the inhibition of formazan dye formation due to superoxide generated by xanthine and xanthine oxidase. The intensity of the chromogen was detected at 550 nm via spectrophotometry (Spectrophotometer DU®-65, Beckman Instruments, Inc., Fullerton, CA). The SOD activity was expressed as unit/ml as determined by comparison of sample values to those obtained from a standard curve prepared using pure SOD (provided in kit) in place of sample homogenate.
Estimation of thiobarbituric acid reactive substances
Thiobarbituric acid reactive substances (TBARS) have been widely used as a general index of lipoperoxidation. 32 We quantitated TBARS as described by Fraga et al. 33 Each sample contained approximately 7–10 isolated MLV pooled in 0.1 M Tris-HCl buffer (pH 7.4) and stored in liquid nitrogen at −80°C. Briefly, 9-mo- and 24-mo-old MLV were homogenized in 0.1 M Tris-HCl buffer (pH 7.4), treated with 0.38% thiobarbituric acid (TBA), 1 N HCl, 15% trichloroacetic acid (TCA) solution, and incubated for 30 min at 90°C in a water bath. After cooling to room temperature, the samples were centrifuged at 3000 g for 10 min, and the absorbance of the supernatant was measured at 535 nm via spectrophotometry (Spectrophotometer DU®-65, Beckman Instruments, Inc.). The sample values were compared to a standard curve generated using 1,1,3,3-tetramethoxypropane in place of sample homogenate and the formed lipoperoxide in each sample was expressed as nM/mg wet tissue.
Detection of superoxide in live mesenteric lymphatic vessels
Dihydroethidium (DHE) (Invitrogen Corp., Carlsbad, CA, Cat. # D-1168), a superoxide fluorescent indicator dye, was used to localize O2- in 9-mo and 24-mo in live MLV. DHE can enter the cell and be oxidized by O2- to yield ethidium, which binds to DNA to produce red fluorescence. The increase in ethidium fluorescence is suggestive of O2- production within the cells. Isolated 9-mo and 24-mo MLV were cannulated on glass pipettes in custom-made single vessel chambers. Their lumen was flushed once with PSS and the samples were then treated from the outside of vessel with DHE (15 μM/L; 1 ml volume per vessel) in PBS buffer. All vessels were incubated at 37°C in a light-protected humidified environment. After 30 min of incubation, vessels were rinsed with PBS to remove unoxidized DHE. After rinsing, vessels were imaged immediately using an Olympus DP72 fluorescent camera and Olympus CKX41 inverted fluorescent microscope equipped with filters suitable for ethidium imaging. The same parameters of image capturing were utilized for all vessels. The fluorescence intensity was analyzed via densitometry using the NIH ImageJ program. Multiple, equal-sized regions of interest (ROI) were selected within each image. ROI intensities were first averaged across the vessel within each image, then averaged across each image acquired across the entire vessel length, and finally averaged between two vessels acquired from each animal.
Detection of mitochondrial ROS (mtROS) in live mesenteric lymphatic vessels
Mitochondria produce a variety of ROS such as ONOO-, and O2.-. MitoTracker Red CM-H2XRos (Invitrogen Corp., Cat. # M-7513) was used to detect the levels of mtROS in live MLV. Isolated 9-mo and 24-mo-old mesenteric lymphatic vessels were cannulated on glass pipettes in a custom-made single vessel chamber. Their lumen was flushed once with PSS, and the vessels were then abluminally treated with MitoTracker Red CM-H2XRos (25 μM/L; 1 ml volume per vessel) in 1X PBS buffer. All vessels were incubated at 37°C in a light-protected humidified chamber. After 30 min incubation, vessels were rinsed once with PBS to remove unoxidized MitoTracker Red CM-H2XRos. After rinsing, vessels were imaged immediately using an Olympus DP72 fluorescent camera and Olympus CKX41 fluorescent microscope equipped with filters suitable for MitoTracker Red imaging. Image capturing and analysis was performed in same manner indicated for DHE imaging above.
Western blot analysis of SOD isoforms and nitro-tyrosine
Seven to ten isolated MLV were homogenized in RIPA buffer (0.5 M Tris pH 7.4, 150 mM NaCl, 1% NP-40, 1% sodium deoxycholate, 0.1% SDS, and 1X preotease inhibitor). Total protein concentration was determined by Micro BCA Protein Assay Kit (Thermo Fischer Scientific Inc., Rockford, IL, Cat. # 23235). Sample proteins were separated under reducing conditions via SDS-PAGE on a Criterion XT 4-12% Bis-Tris gel (Bio-Rad Laboratories, Hercules, CA, Cat. # 345-0125), and then electrotransferred to nitrocellulose membrane. After transfer, the blots were blocked for 2 h with 5% nonfat dry milk in 0.05% Tween 20 in 1X Tris buffered saline (TTBS) buffer, followed by incubation for 2 h in blocking solution containing one of the following primary antibodies against: Cu/Zn-SOD (Millipore, Billerica, MA, Cat. # 07-403, dilution 1:500); Mn-SOD (Millipore, # 06-984, dilution 1:1000); EC-SOD (Millipore, Cat. # 07-704, dilution 1:1000); nitro-tyrosine (Cell Signaling Technology, Inc., Danvers, MA, Cat. # 9691, dilution 1:1000), and β-actin (Sigma-Aldrich Corp., St. Louis, MO, Cat. # A1978, 5 μg/ml). The blots were washed with TTBS and incubated with goat anti-mouse IgG or anti-rabbit IgG HRP conjugate (Jackson ImmunoResearch Laboratories, Inc., West Grove, PA, Cat. # 715-035-151 and 805-035-180) for 1 h. After further washing with TTBS, the bands were detected via chemiluminescence (SuperSignal West Dura Extended Duration Substrate, Thermo Fischer Scientific Inc., Rockford, IL, Cat. # 34076), and imaged using a LAS-4000 imager (FujiFilm, Tokyo, Japan). SOD isoforms and nitro-tyrosine expression was analyzed via densitometry using MultiGage software (FujiFilm) and normalized to β-actin expression within the same sample.
Immunohistochemical analysis of SOD isoforms and Nitro-tyrosine
Isolated 9-mo and 24-mo-old Fischer-344 rat MLV were cannulated on glass pipettes in a custom-made single vessel chamber, fixed for 30 min at 0°C in 100% methanol, and washed with 0.05% Tween-20 in PBS (PBST). The vessels were blocked overnight at 4°C in 5% normal goat serum (Jackson ImmunoResearch Laboratories, Inc., Cat. # 005-000-121) in PBST, washed three times for 15 min each in PBST, and then incubated 1 h in PBST containing primary antibodies against: Cu/Zn-SOD (Millipore, Cat. # 07-403, dilution 1:500); Mn-SOD (Millipore, Cat. # 06-984, 10 μg/ml); EC-SOD (Millipore, Cat. # 07-704, 2 μg/ml) and nitro-tyrosine (Abcam, Cambridge, MA, Cat. # ab61392, 8 μg/ml). After washing three times for 15 min each with PBST, the vessels were incubated 1 h in PBST containing goat anti-mouse IgG2a (Cat. # A21241) or anti-rabbit IgG (Cat. # A21245) Alexa Fluor 647 conjugate (all: Invitrogen Corp.,). After further washing in PBST, the vessels were mounted on glass slides under coverslips using ProLong Gold antifade reagent (Invitrogen Corp., Cat. # P36934) and imaged using an Olympus DP72 fluorescent camera and Olympus CKX41 fluorescent microscope equipped with filters suitable for AlexaFluor 647 imaging. Image capturing and analysis was performed in same manner indicated for DHE imaging above.
Statistical differences were determined Student's t-test (JMP software version 9.0.0. for Windows) and considered significant at p<0.05. Data presented as mean±SEM. All data acquired from vessels obtained from a single animal was averaged and considered one sample.
Results
Total SOD enzyme activity and lipid peroxidation in mesenteric lymphatic vessels
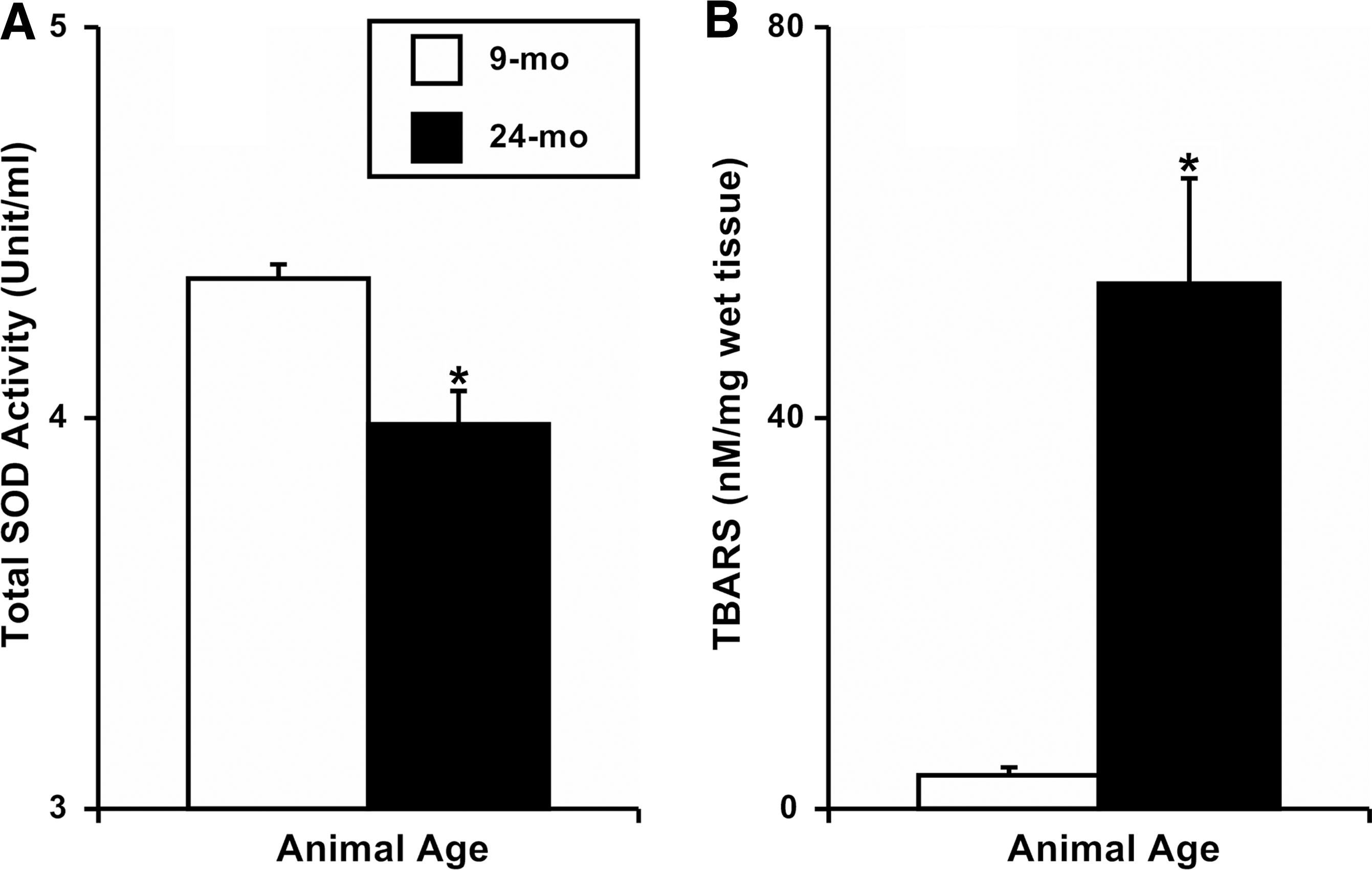
We measured total activity of SOD in adult and aged MLV and found a significant decrease in 24-mo MLV (3.98±0.08 Unit/ml) compared to 9-mo vessels (4.36±0.04 Unit/ml; Fig. 1A). Additionally, we analyzed TBARS as an indicator of lipid peroxidation and found that the concentration of TBARS was significantly elevated in 24-mo MLV (53.7±10.8 nM/mg wet tissue) when compared to 9-mo vessels (3.4±0.8 nM/mg wet tissue; Fig.1B).
Determination of total cellular superoxide and mitochondrial ROS production in mesenteric lymphatic vessels
We measured the total levels of cellular superoxide by DHE fluorescence in live adult and aged MLV. After 30 min of incubation with this superoxide dye, numerous fluorescently labeled cells were identified in MLV, and the density of fluorescent signal was significantly higher (∼97%) in the 24-mo MLV (24.7±2.9 arbitrary units) when compared to 9-mo vessels (12.5±0.8 arbitrary units; Figs. 2A–2C). Additionally, we assessed mitochondrial ROS production using the mitochondria specific ROS-sensitive fluorescent dye MitoTracker Red CM-H2XRos and identified significantly elevated (∼61%) production of mitochondrial ROS in 24-mo MLV (21.3±1.9 arbitrary units) compared to 9-mo vessels (13.2±2.1 arbitrary units; Figs. 2D–2F).
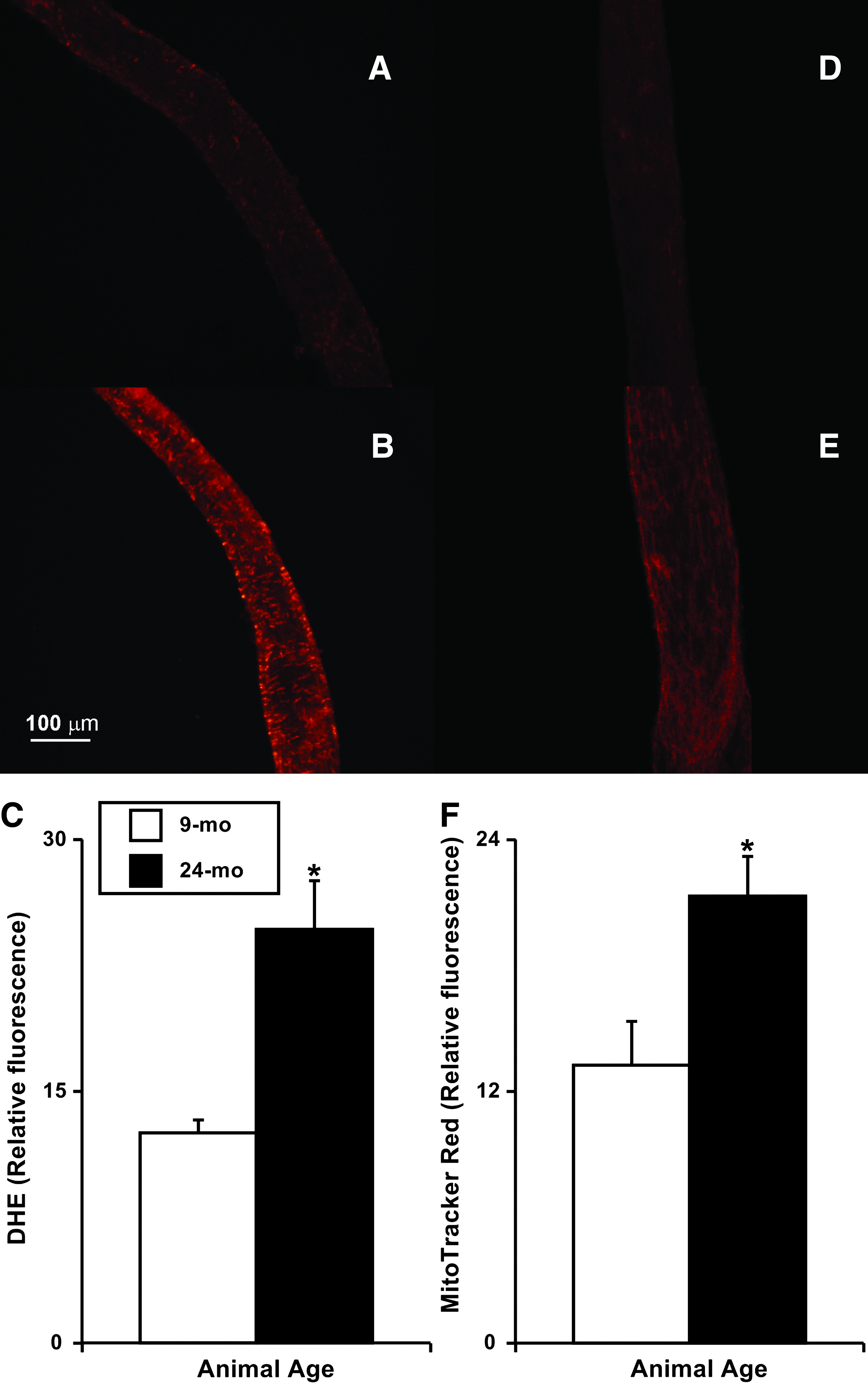
Representative images of the in situ detection of O2- in live 9-mo
Protein expression of SOD isoforms and nitro-tyrosine in the mesenteric lymphatic vessels
We performed Western blot analyzes to compare the protein expression of SOD isoforms (Cu/Zn- SOD, Mn-SOD, and EC-SOD) and nitro-tyrosine (as an indicator of oxidative damage) in 9-mo and 24-mo MLV. We found that the Cu/Zn-SOD isoform was expressed significantly lower (∼51%) in 24-mo MLV (normalized signal intensity (NSI): 2.53±0.33) compared to 9-mo MLV (NSI: 5.16±0.53), while the EC-SOD isoform was expressed significantly higher (∼78%) in 24-mo MLV (NSI: 5.42±0.53) compared to 9-mo MLV (NSI: 3.04±0.40; Fig. 3). We did not find significant differences in the expression of the Mn-SOD protein isoform between 9-mo and 24-mo MLV samples (NSI: 0.96±0.18 versus 1.60±0.31, respectively; Fig. 3). Protein bound nitro-tyrosine formation is commonly used to demonstrate nitric oxide-dependent oxidative damage. In this study, we examined the quantity of nitro-tyrosine formation in 9-mo and 24-mo MLV via Western blot. We observed a significant increase (∼148%) in the expression of nitro-tyrosine protein corresponding to 215 kDa band in 24-mo MLV (NSI: 1.05±0.18) when compared to 9-mo MLV (NSI: 0.42±0.14; Fig. 3). Conversely, we did not observe significant differences in the 36 kDa band between age groups (data not shown).
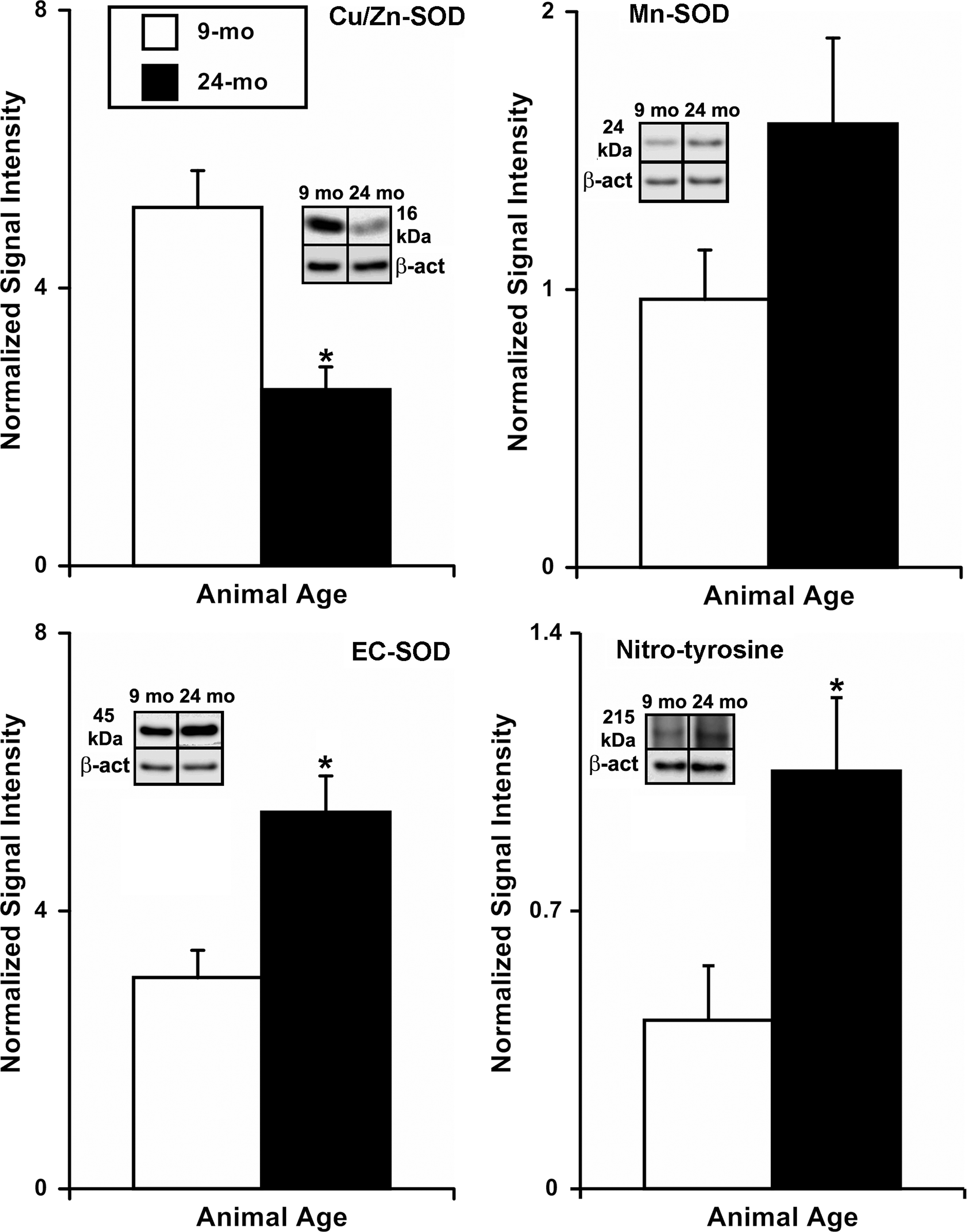
Western blot analysis of aging-associated changes in SOD isoform protein expression and nitro-tyrosine formation in rat mesenteric lymphatic vessels. Insets show representative 16 kDa Cu/Zn-SOD, 24 kDa Mn-SOD, 45 kDa EC-SOD, and 215 kDa nitro-tyrosine bands along with their corresponding β-actin bands in 9-mo and 24-mo samples. The number of samples of each age group (9 mo/24 mo): Cu/Zn-SOD: n=6/5, MnSOD: n=6/5, EcSOD: n=6/5, nitro-tyrosine: n=5/5. * indicates significant differences (p<0.05) between 9-mo and 24-mo age groups.
Immunohistochemical analysis of SOD isoforms and nitro-tyrosine
We performed immunohistochemical labeling of SOD isoforms (Cu/Zn- SOD, Mn-SOD, and EC-SOD) and nitro-tyrosine (as an indicator of oxidative damage) in 9-mo and 24-mo MLV. We found that the signal intensity (relative to background) for Cu/Zn-SOD isoform was significantly lower (∼28%) in 24-mo MLV (relative signal intensity (RSI): 6.31±1.32) compared to 9-mo MLV (RSI: 8.73±1.64; Fig. 4). At the same time, we did not find significant differences in the signal intensities of Mn-SOD and EC-SOD isoforms between 9-mo and 24-mo MLV samples (RSI: 9.93±4.86 versus 6.87±1.40 and 11.54±1.33 versus 11.87±2.67, respectively; Fig. 4). We also examined the signal intensity of nitro-tyrosine in 9-mo and 24-mo MLV via immunohistochemical labeling to demonstrate NO-dependent oxidative protein damage. We observed a significant increase (∼47%) in signal of nitro-tyrosine labeling in 24-mo MLV (RSI: 10.67±0.44) when compared to 9-mo vessels (RSI: 7.24±0.43; Fig. 4).
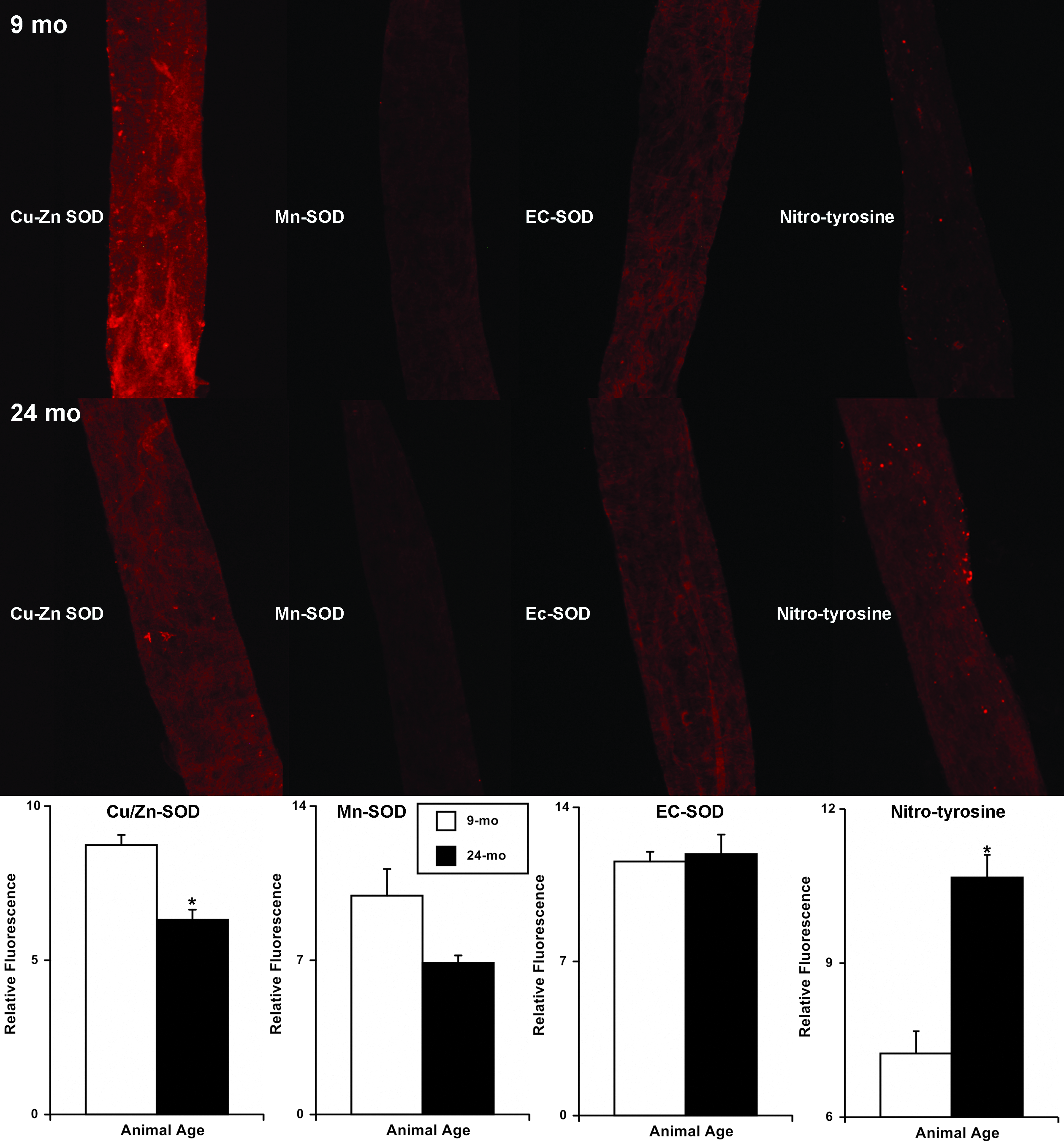
Immunohistochemical labeling (representative images) and analysis of aging-associated changes in SOD protein isoform expression and nitro-tyrosine formation in rat mesenteric lymphatic vessels. The numbers of animals (and used vessels) for each age group (9mo/24mo) are: Cu/Zn-SOD: n=5/4 (8/8), MnSOD: n=4/4 (7/8), EcSOD: n=3/3 (5/6), nitro-tyrosine: n=4/4 (7/7). * indicates significant differences (p<0.05) between 9-mo and 24-mo age groups.
Conclusions
Aging is commonly defined as a functional loss over time that is accompanied by an inability to withstand stress or insult. We recently demonstrated that, as with many other organs, there is an aging-associated decline in the contractile capacity of lymphatic vessels;9,11 however, the cellular events responsible for this functional loss remain undefined. The role of ROS such as O2- in the aging process was initially proposed as the “free radical theory” by Harman in the 1950s, 34 whereby ROS damage the cellular constituents resulting in a functional decline of the organ systems finally leading to cell death. The participation of ROS is well documented in many pathological conditions typical for the elderly such as cardiovascular diseases, diabetes, cancer, and arthritis. Fridovich 35 reported that under normal conditions, intracellular O2- concentrations are kept at low levels because eukaryotic cells contain large amounts of SOD (4–10*10−6 M). There are numerous reports suggesting that ROS at moderate concentrations act as signaling molecules and play an important role in the regulation of various vascular cellular functions.36–40 Acutely, ROS overproduction can interfere with important signaling cascades such as inactivation of bioavailable NO, alterations in prostaglandin metabolism, or deregulation of calcium and phosphorylation cascades.41,42 Chronically, ROS overproduction can lead to irreversible oxidation and accumulation of oxidized biological macromolecules, for example, increase in DNA mutations. 43 Despite the crucial involvement of lymph transport vessels in numerous physiological and pathological processes, there are no reports available on the involvement of oxidative stress in the aging of lymphatic vessels. In this present study, we provided the first evidence for aging-associated elevations in mesenteric lymphatic vessel O2- and oxidative cellular damage.
Endothelial cell membrane damage is thought to be an early event leading to microvascular dysfunction and may be initiated by several factors including lipid peroxidation. ROS such as hydrogen peroxide, superoxide, and hydroxyl radicals damage biomembranes and induce peroxidation of lipids, leading to an increase in cell permeability and loss of endothelial integrity. In this current study, we observed a significant increase in the concentration of TBARS in mesenteric lymphatic vessels of 24-mo compared to 9-mo rats. We considered this observed increase in the levels of TBARS (Fig. 1) to be an indicator of elevated aging-associated free radical-induced lipoperoxidation in MLV that could ultimately be associated with cellular membrane damage. These findings correlate with observations by Ohkuma 44 who proposed that interstitial accumulation of lipoperoxide (a breakdown product of cell membranes) may be potent toxic factor responsible for trophic changes associated with chronic lymphedema (a pathological condition also associated reduced lymphatic contractility).45–47 In 1993, Ohkuma 48 suggested that during the impaired lymph drainage, lipoperoxides, which are normally transported in lymph, may be deposited in the skin and contribute to the soft tissue changes characteristic of chronic lymphedema. With respect to Dr. Ohkuma findings and given the role of mesenteric lymphatics in transporting dietary lipids, we propose that aging-associated lipoperoxidative damage in the mesenteric lymphatic network observed in the present study and the impairment of the mesenteric lymph transport function in aged vessels reported by us earlier 11 may predispose the elderly to excessive mesenteric fat deposition, thereby potentially contributing to the lipid dysregulation commonly seen in aging-associated diseases. Potential inflammatory-related breakdown of antioxidant systems in mesenteric lymphatic vessels might be considered as an important trigger in the development of metabolic syndrome even at earlier stages of the life span, which requires further detailed investigations.
As discussed above, diminished NO bioavailability due to increased O2- production is one of the major mechanisms responsible for the impaired endothelium-dependent vasodilator responses observed during aging. The interaction between NO and superoxide depletes NO bioactivity, thereby altering several key vascular functions of which NO is a pivotal mediator, including regulation of smooth muscle tone, platelet activation, and vascular cell signaling.36,41,42 In this study, we report the first evidence of a significant increase in superoxide production in live aged mesenteric lymphatic vessels (Fig. 2), thus confirming the existence of the aging-associated oxidative stress in MLV, which may be a key factor in the cascade of events occurring in the mesenteric lymphatic network as a biological consequence of aging.
In normal physiological conditions, antioxidants enzymes prevent the detrimental effects of O2-. In studying the role of antioxidant defenses in aging, attention has been given to the role of superoxide dismutase, which efficiently and specifically catalyzes the dismutation of O2- to H2O2 and O2. SODs are also involved in the modulation of NO bioactivity. Normally, tissues express three isoforms of SODs including Cu/Zn-SOD (SOD1), Mn-SOD (SOD2), and extracellular SOD (EC-SOD or SOD3). Cu/Zn-SOD is an abundant copper- and zinc-containing cellular protein that is present in the cytosol, nucleus, peroxisomes, and mitochondrial inner membrane. Its primary function is to lower the intracellular steady-state concentration of O2-. 49 Mn-SOD is a mitochondrial enzyme that disposes of O2- generated by respiratory chain activity. It can be induced to protect against pro-oxidant insults. Conversely, Mn-SOD activity is decreased in physiologic aging and in diseases such as cancer, asthma, and transplant rejection. 50 EC-SOD plays an important role in regulating blood pressure and vascular contraction, at least in part through modulating the endothelial function by controlling the levels of extracellular O2- and nitric oxide bioactivity in the vasculature.51,52 Aging has been shown to produce alterations in the expression and activity of SOD in several tissues.53,54 In blood vessels, SOD activity and/or expression has been shown to be altered with aging.55,56 Zawieja et al. 10 reported that in young mesenteric lymphatic vessels, decreases in ejection fraction, contraction frequency, and lymph pump flow caused by high dose superoxide anion treatment were attenuated by SOD application. In our current study, we observed that total SOD activity was significantly decreased in the aged mesenteric lymphatic vessels compared to adult vessels (Fig. 1). This result may be a contributing factor to the elevated ROS in aged vessels (Fig. 2) and/or may augment the oxidative stress on the vessels during aging leading to elevated levels of oxidative damage (as seen in Figs. 1B, 3, and 4). In parallel with data obtained by Zawieja et al., 10 we propose that diminished SOD enzyme activity may play an important role in development of the aging-associated impairment of the mesenteric lymph transport function reported by us earlier. 11 However, further functional investigations are necessary to confirm the role of SOD protein dysfunction during aging in mesenteric lymphatic vessels.
The relative expression of different SOD isoforms in cells and tissues has been investigated extensively and provides clues as to the sources of O2- in pathophysiologic states. Cu/Zn-SOD is the predominant isoform in microvessels where it may scavenge O2- to increase the bioavailability of NO, which in turn improves endothelium-dependent vascular function. Didion et al. 25 suggested that the release of NO from the endothelium is dependent on Cu/Zn-SOD, whereas EC-SOD activity is thought to be required for the protection of NO as it diffuses through the vascular wall. In both large arteries and microvessels, deficiency in Cu/Zn-SOD results in increased levels of vascular superoxide and peroxynitrite, increased myogenic tone, augmented vasoconstrictor responses, and impaired endothelium-dependent NO-mediated relaxation.24,57,58 In the current study, we confirmed by Western blot analyses and immunohistochemical labeling significantly lower levels of Cu/Zn-SOD isoform expression in 24-mo compared to 9-mo MLV (Figs. 3 and 4, respectively). Such changes may be a contributor to the aging-associated impaired endothelium-dependent NO-mediated regulation on the lymphatic vessels.9,11 While we did observe an increase EC-SOD isoform expression in 24-mo compared to 9-mo MLV via Western blot (Fig. 3), we were not able to see any difference in expression of EC-SOD via immunohistochemical labeling (Fig. 4). While these differing results for EC-SOD may simply reflect the detection abilities of these two techniques, it is important to note that total SOD activity in 24-mo MLV is still decreased relative to the 9-mo vessels (Fig. 1A). Thus, even if EC-SOD protein expression is indeed elevated in the 24-mo vessels (as indicated via Western blot), the total antioxidant activity of the aged vessels is still depressed relative to the adult vessels, presumably due to the consistent depression in Cu/Zn-SOD protein expression. Follow-up studies are needed to assess the specific importance of the Cu/Zn-SOD isoform depletion and the possible EC-SOD enrichment in the development of the aging-associated oxidative damage in lymphatic vessels.
The increased levels of O2- in aged MLV (Fig. 2) may result in increased NO scavenging and subsequent ONOO- formation, which is known to initiate oxidative modification of proteins ultimately leading to lipid peroxidation or DNA damage.20,59,60 It is known that peroxynitrite at submicromolar concentrations causes the nitration of protein-bound tyrosine residues in Mn-SOD or prostacyclin synthase. 61 Our data clearly demonstrate significantly increased nitro-tyrosine levels in aged mesenteric lymphatic vessels (Figs. 3 and 4), thus indicating increased formation of ONOO-. This last observation suggests that there may be diminished levels of basal NO in the vessel due to the inability of SOD to completely scavenge the elevated O2- formation during aging. Consequently, the loss of NO bioactivity associated with increased vascular O2- in the mesenteric lymphatic vessels during aging may play a potentially important role in the pathogenesis of lymphatic endothelial dysfunction. We also found that mitochondrial ROS increased in the aged MLV compared to their adult counterparts. This also might enhance the reaction between O2- and NO. It is demonstrated 62 that protein tyr-nitration mediated by ONOO-, increases with age and ultimately leads to the inhibition of mitochondrial energy production by causing site-specific lesions in the electron transport chain, in enzymes involved in the citric acid cycle, or in enzymes necessary for energy transfer. In connection with these findings, aging-associated weakening of lymphatic pumping and disturbances in its NO-dependent regulatory mechanisms9,11 may be linked to increased nitro-tyrosine levels in aged lymphatic vessels.
In conclusion, in this study, we demonstrate for the first time evidence of aging-associated oxidative stress in mesenteric lymphatic vessels, including increased levels of the superoxide and mitochondrial reactive oxygen species. Additionally, we demonstrated reduced antioxidant activity within the cell via reduced total SOD enzymatic activity concomitant with decreased protein expression and immunohistochemical labeling of Cu/Zn-SOD protein in 24-mo old mesenteric lymphatic vessels. While we did find evidence of increased extracellular SOD protein expression in aged mesenteric lymphatic vessels, the impact of this finding on the oxidative environment of aged lymphatics needs further investigation. Finally, we demonstrated elevated levels of oxidative damage to proteins and lipids in the aged mesenteric lymphatic vessels. Cumulatively, these data suggest that oxygen-derived radicals such as O2- and ONOO- may be at least partially responsible for the aging-associated lymphatic pump dysfunction previously reported by us. Subsequent follow-up studies are necessary to investigate in-depth the nature and functional importance of these present findings.
Footnotes
Acknowledgments
The authors thank Kelci Jackson and Stacy Henderson for their help with data analysis.
Author Disclosure Statement
For all authors, no competing financial interests exist.
This work was supported in parts by the National Institutes of Health (NIH RO1 AG-030578 and HL-094269) and by Texas A&M Health Science Center College of Medicine and Department of Systems Biology and Translational Medicine.