
Abstract
Abstract
Sphingosine-1-phosphate (S1P) is a potent sphingolipid metabolite that regulates a number of biological processes critical for cancer. S1P produced inside cancer cells is exported and exerts its extracellular functions by binding to its specific receptors in an autocrine, paracrine, and/or endocrine manner, which is known as inside-out signaling. S1P is also known to exert its intracellular functions especially in the inflammatory process, but its relevance to cancer biology remains to be elucidated. Recently, there have been growing interests in the role of S1P in breast cancer progression, including angiogenesis and lymphangiogenesis. Our group demonstrated that activation of sphingosine kinase 1, the enzyme that catalyzes the phosphorylation of sphingosine to S1P, is a key step of this process. In this review, we will cover our current knowledge on the role of S1P signaling pathway in breast cancer progression with an emphasis on its role in tumor-induced lymphangiogenesis.
Lymph Node Metastasis Is a Major Prognostic Predictor in Breast Cancer
Over the past one and a half decade, understanding of the cellular and molecular aspects of physiologic and tumor-induced lymphangiogenesis has advanced remarkably after the discovery of vascular endothelial growth factor (VEGF)-C/D and its function in promoting the growth of lymphatic vessels. 6 Initially, the study of lymphangiogenesis largely focused on the primary site of tumor growth and adjacent tissues, which is known as “tumoral lymphangiogenesis”.7,8 However, lymphangiogenesis was also observed around regional lymph nodes, in particular the sentinel nodes where tumor cells first metastasize, a phenomenon now known as “lymph node lymphangiogenesis”.9,10 Lymph node lymphangiogenesis and increased lymph flow through tumor-draining lymph nodes are speculated to actively promote lymphatic metastasis. 2 Recent evidence indicates that tumor cells can also induce lymph node lymphangiogenesis even before they metastasize, and that metastatic tumor cells continue to induce lymphatic vessel growth within sentinel lymph nodes, which is thought to promote their further metastatic dissemination. 11 Our recent findings indicate that sphingosine-1-phosphate (S1P) play a key role in this process, which we will discuss later in detail. 12
What is Sphingosine 1-Phosphate (S1P)?
Sphingosine 1-phosphate (S1P) is a potent lipid mediator that regulates a number of biological processes critical for cancer, such as cell survival, growth, migration, invasion, angiogenesis, vascular maturation, and immune reactions.13–15 Increased S1P production has been implicated in various physiological and pathological processes such as cancer, atherosclerosis, allergy, and autoimmune diseases such as multiple sclerosis.13,16,17 S1P exerts its functions either by binding to its specific receptor in autocrine, paracrine, and/or endocrine manner after it is exported from the cells, also known as “inside-out signaling”, or by binding to its intracellular targets.
S1P, ceramide (N-acyl sphingosine), and sphingosine are metabolites of sphingolipids, which are structural components of cell membranes. Sphingolipid metabolites are now recognized as important players in cancer biology. In contrast to S1P, which is associated with growth and survival, its precursors, sphingosine and ceramide, are associated with cell growth arrest and apoptosis. 18 Ceramide can be produced de novo, or by breakdown of sphingomyelin and more complex sphingolipids. Sphingosine is only formed as a result of ceramidase-catalyzed hydrolysis of ceramide and can be either deacylated to ceramide or phosphorylated to S1P. Sphingosine kinases (SphKs) convert sphingosine to S1P, thereby enhancing cell growth and survival, and playing a crucial role in regulation of this balance.19,20 Conversely, S1P phosphatases tilt the balance from S1P to sphingosine and ceramide. S1P levels in the cells are tightly regulated by the balance between its synthesis by SphKs and its degradation. Indeed, it is found that SphK1-specific inhibitor not only decreased S1P levels but concomitantly increased levels of its proapoptotic precursor ceramide 21 (Fig. 1).
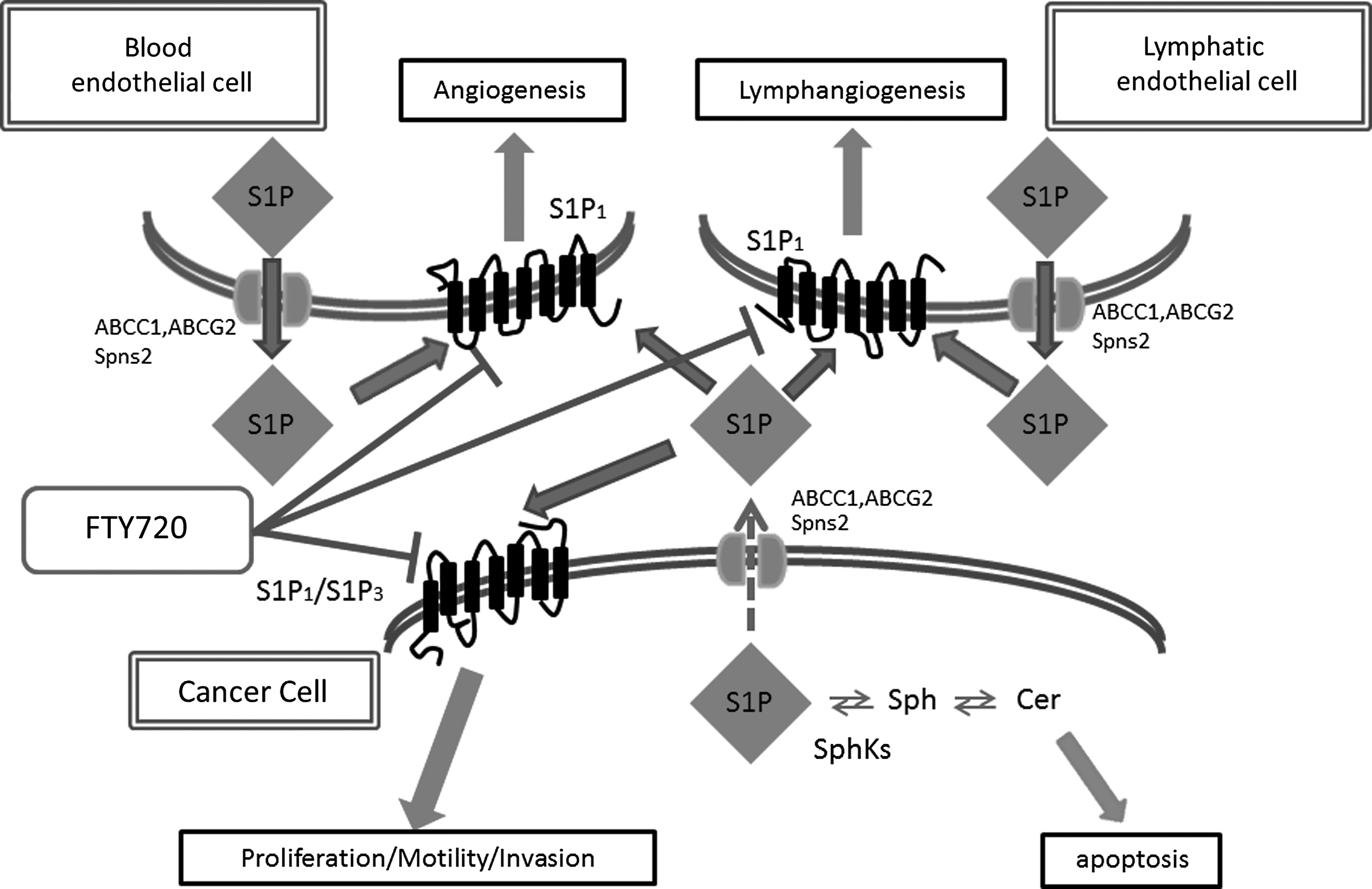
S1P produced and released from cancer cells acts in an autocrine/paracrine manner to activate S1P receptors present on the cancer cells, blood endothelial cells, or lymphatic endothelial cells, and stimulates angiogenesis and/or lymphangiogenesis.
Sphingosine Kinases 1 and 2
Currently two isoforms of sphingosine kinases (SphKs), SphK1 and SphK2, are identified, which may at least partially compensate for the lack of the other. Double knock-out mice of SphK1 and SphK2 that completely eliminate S1P are embryonically lethal due to severely disturbed angiogenesis and hemorrhage during development. On the other hand, either SphK1 or SphK2 single knock-out mice are alive and healthy. 22
Many studies were conducted on the role of SphK1 in cancer, however, the role of SphK2 in cancer remains a subject of debate. 23 Although both SphK1 and SphK2 share overall homology and produce the same product, S1P, they display different catalytic properties, subcellular locations, tissue distribution, and temporal expression patterns during development and have unique and specific functions. SphK1 translocates to the plasma membrane when activated by growth factors. This translocation enables localized production of S1P in the vicinity of its cell surface receptors, which enables the “inside-out signaling” of S1P.13,23,24 In contrast, SphK2 is localized in the nucleus of many types of cells, 25 and in the inner mitochondrial membrane 26 and thought to control cellular functions.
SphK1 is a critical regulator of the balance between the pro-growth and anti-apoptotic S1P and its pro-apoptotic precursors, ceramide and sphingosine. Numerous previous studies have shown that overexpression of SphK1 promotes tumorigenesis and cancer progression. 27 For instance, MCF-7 cells that overexpress SphK1 formed more and larger tumors than vector transfectants with higher microvessel density in their periphery when implanted into nude mice. 28 In accordance, downregulation of SphK1 in cancer cells reduces growth, increases apoptosis, and enhances chemosensitivity. 29 Growth factors, hormones important for progression of cancer, and cytokines stimulate SphK1 and production of S1P.20,30 SphK1 can be stimulated by a wide variety of growth factors, including estradiol (E2), platelet-derived growth factor (PDGF), vascular endothelial growth factor (VEGF), epidermal growth factor (EGF), hepatocyte growth factor, cytokines (tumor necrosis factor-alpha (TNFα)), steroid hormones, and GPCR ligands (acetylcholine), lysophosphatidic acid, S1P itself, and many other factors.23,31–42 Furthermore, it has been shown that SphK1 is upregulated in a variety of solid tumors, including breast, colon, lung, ovary, stomach, uterus, kidney, and rectum. 43 A study using gene expression microarray analysis of 1269 human breast tumor samples revealed a worse outcome of patients with high SphK1 expression, suggesting its value as a prognostic marker. 44
Compared with SphK1, the functions and regulation of SphK2 in cancer have been unclear for a long time. Recently, SphK2 is identified to be primarily located in the nucleus and mitochondria and is activated by phosphorylation by extracellular signal-regulated kinase (ERK) ½. 25 Endogenous SphK2 is mainly associated with isolated chromatin and was not detected in the nucleoplasm. SphK2 associates with histone deacetylases (HDAC) 1 and 2 in repressor complexes and is selectively enriched at the promoters of the genes encoding the cyclin-dependent kinase inhibitor p21 or the transcriptional regulator c-fos, where it enhances local histone H3 acetylation and transcription. 25 SphK2 can be stimulated by EGF and the protein kinase C activator, phorbol ester.45,46 Both of these activate ERK1, which in turn binds to SphK2 and phosphorylates it on Ser351 and Thr578, increasing its enzymatic activity. 46 It is reported that phosphorylation of SphK2 is catalyzed by protein kinase D, which leads to its nuclear export for subsequent cellular signaling. 47
S1P Receptors
Many of the biological effects of S1P are mediated via specific G protein-coupled receptors (GPCRs), now designated S1P receptors. They were formerly called endothelial differentiation gene (EDG) receptors; to date, five members, EDG-1/S1P1, EDG-5/S1P2, EDG-3/S1P3, EDG-6/S1P4, and EDG-8/S1P5 have been identified.
S1P1 receptor was the first identified S1P receptor. 48 S1P1 receptors are ubiquitously but differentially expressed on a vast majority of cells. Their physiological function includes cell motility, lymphocyte trafficking, angiogenesis, vascular maturation, and vascular tone.32,49–52 Homozygous S1P1 null mice are embryonically lethal due to incomplete coverage of blood vessels by smooth muscle cells and massive hemorrhage at 12.5–14.5 days of gestation. 51 S1P1 expression is strongly induced in tumor vessels, and injection of S1P1 small interfering RNA into xenograft tumors in mice suppressed vascular stabilization, angiogenesis, and tumor growth in vivo. 53 Recently, there have been a great interest in this receptor due to the FDA approval of FTY720 (fingolimod, trade name Gilenya, Novartis), a functional antagonist of S1P1 that sequesters lymphocytes in lymph nodes, as a treatment of multiple sclerosis. 13 Further, there are accumulations of data on the role of S1P1 in lymphangiogenesis that we will describe in detail later.
S1P3 receptors have been shown to stimulate cell migration as well. It has been shown to exert its effects on the cardiovascular system, regulating bradycardia and hypertension, pulmonary epithelial barrier, and hearing.54,55
The other S1P receptors have different phenotypes, such as proper functioning of the auditory and vestibular systems,56–59 coronary vasoconstriction60–62 by S1P2 receptor, immune compartments, and leukocytes,63,64 and the mobilization of NK cells to inflamed organs 65 by S1P4 receptor.
Extracellular Target of S1P
It has been well demonstrated that the “inside-out” signaling by S1P plays a role in many human diseases, including cancer and inflammation. 13 How “inside-out” signaling via S1P receptors is accomplished has been an intriguing question of the field since S1P does not spontaneously traverse the lipid bilayer due to its polar head group, where the SphKs and S1P production are both on the cytosolic side of the plasma membrane, and S1P receptors bind S1P on the exoplasmic side. While it has been suggested that SphK proteins themselves may be secreted and produce S1P extracellularly, our group demonstrated that the ATP-binding cassette (ABC) transporters, ABCC1 and ABCG2, mediate secretion of intracellularly produced S1P. In MCF-7 breast cancer cells, estradiol stimulates ERK1/2 by activation of SphK1, which results in generation of S1P, leading to export of S1P via ABCC1 and/or ABCG2, and activation of S1P receptor 31 (Fig.1). Our result was consistent with prior studies from several laboratories, including ours, which have suggested the involvement of ABC transporters in S1P export from mast cells,66,67 platelets, 68 endothelial cells, 69 astrocytes, 70 thyroid follicular cells, 71 fibroblasts, 72 and erythrocytes. 73
Recently, a new transporter, Spinster 2 (Spns2), was identified as a transporter of S1P as well (Fig.1). Spns2 rescued the zebrafishcaridia bifida phenotype and has been shown to be responsible for S1P release. 74 The phosphorylated form of FTY720 (FTY720-P) is also reported to be transported by Spns2 through the same pathway as S1P, 75 and Spns2 is responsible for egress of mature T cells and immature B cells from thymus and bone marrow. 76 Pham et al. reported that transcripts for the mouse Spns2 orthologue were abundantly expressed in lymphatic endothelial cells. 77 This report implicates a role of Spns2 in lymphatic vessel development or function; however, there are no data published to date in this regard and further investigation on this topic is awaited.
In addition to anti-apoptotic effects, extracellular S1P also enhances cell proliferation in many cell types, including endothelial cells, 78 vascular smooth muscle cells,79,80 and fibroblasts. 81 As described above, SphK1, which is mainly responsible for the production of secreted S1P in response to receptor activation, is overexpressed in many types of cancers and possesses many characteristics of an oncogene. 82 Finally, despite a multitude of studies focusing on physiological functions of S1P receptors, some actions of S1P resulting from activation of SphKs have been shown to be independent of S1P receptors.
Intracellular Target of S1P
Over the years, most of the biological effects of S1P were thought to be attributed to the “inside-out” signaling, with exported S1P binding to its five receptors. Direct intracellular targets of S1P were not identified until recently, despite the fact that S1P has been implicated to function inside the cell independent of S1P receptors. 83 The suspicion that S1P may have direct intracellular targets stems from the fact that S1P elicit its effects in organisms lacking known S1P receptors.84–87 Recently, it was found that S1P has intracellular sites of action and regulates fundamental biological processes, including gene expression, mitochondrial functions, protein processing, and inflammation.25,26,88 Dihydro-S1P, generated exclusively via de novo sphingolipid biosynthesis pathway, binds to extracellular S1P receptors via inside-out signaling, but not to any of the known intracellular targets of S1P; thus, comparison of S1P and dihydro-S1P levels provides us some clue on the involvement of de novo synthesis and the target of S1P.
As this is a new development in S1P research, its impact on cancer has yet to be explored. However, due to the nature of the currently identified intracellular targets, including HDACs, 25 E3 ubiquitin ligases, 88 and prohibitin, 89 it seems very plausible that future research will discover direct links to cancer. For example, pharmacological inhibitors of HDAC are currently under investigation in clinical trials as anti-cancer drugs. Given recent finding that endogenous SphK2 inhibits HDAC1 and 2, it is tempting to speculate that it is functioning as a cancer suppressor. In agreement with this notion, nuclear SphK2 has been shown to inhibit cell proliferation, likely via p53-independent upregulation of p21 expression. 25 Another hint is that some of the effects mediated by TNFα depend on the activation of SphK1and production of S1P. 88 It is well established that TNFα signaling pathway is important in inflammation-induced tumorgenesis, and it is speculated that S1P may play a critical role in this process.
S1P and Breast Cancer
Breast cancer is the most common cancer and the second leading cause of death from malignancy in women in the United States, and thus has been the object of intensive research that is now revealing the complexity of this disease. SphK1 expression was generally higher in breast cancers than normal breast tissue, whereas low levels in tumors compared with the normal specimens were rare. 43 SphK1 has also been shown to be associated with tumor angiogenesis and resistance to radiation and chemotherapy. Gene expression microarray analyses of 1269 breast tumor samples revealed a worse outcome for patients with high SphK1 expression, further supporting the notion that overproduction of S1P results in a worse prognosis for cancer patients and suggesting its value as a prognostic marker.18,23,44
17β-Estradiol (E2), a major naturally occurring estrogen, has dual actions to increase SphK1 activity: a rapid and transient effect and a delayed but prolonged effect. The rapid activation of SphK1 induced by E2 is apparently mediated by membrane-associated ER coupled with Gi proteins (G-protein-coupled receptor 30), whereas the delayed action relies on transcriptional activity of nuclear ERα. 39 Recently, it was reported that SphK1/S1P pathway mediates the E2-induced transactivation of EGFR that is associated with carcinogenesis, including lung cancer and glioblastoma multiforme. 39 Moreover, we have reported that E2 induces export of S1P from breast cancer cells via ABCC1 and ABCG2 transporters. 31 Thus, the accumulating evidence suggests that SphK1/S1P pathway may contribute to E2 signaling.
Estrogen receptor (ER) status is a major determinant of breast cancer. ER-negative breast cancer patients have earlier disease recurrence and worse prognosis compared with ER-positive breast cancer patients. This is partly because pharmacological inhibition of E2 synthesis (such as aromatase inhibitors) or E2 binding to the ER (such as tamoxifen) is effective treatment for ER-positive tumors, as opposed to ER-negative tumors that do not have that option. Therefore, there is still a need of novel targets for therapeutic intervention that can provide better treatment options for ER-negative breast cancers. When comparing ER-positive and ER-negative tumors, SphK1 is more highly expressed in ER-negative tumors that are known for their higher proliferative activity. 44 Moreover, immunohistochemical analysis identified the carcinoma cells in breast tumors as the major source of SphK1 expression. 44 Even in the ER-positive breast cancer, higher expression of SphK1 correlated with poor patient survival rates, and it was reported to be associated with development of tamoxifen resistance and earlier disease recurrence while on tamoxifen.90,91 Therefore, Sphk1 expression levels in combination with ER status are proposed to be a good biomarker to predict response to tamoxifen. 91 Further, there was a report that a SphK1-selective inhibitor (SKI-II) abrogated ER signaling, possibly through direct binding to the ER similar to tamoxifen, diminished viability and proliferation of breast carcinoma cells without similar effects on normal breast epithelial cells. 92 Additionally, high levels of S1P1 and S1P3 expression in ER-positive breast cancer patients are also reported to be correlated with poor prognosis as well. 91 There have been several reports on the effect of a Sphk2-selective inhibitor ABC294640 on breast cancer that it altered ER signaling in MCF-7 cells, 93 induced apoptosis through the intrinsic pathway, 94 and decreased chemo-resistant breast cancer tumor growth in vivo.
HER2/neu is also known to correlate with poor prognosis and therapeutic resistance in breast cancer. HER2/ErbB2 was reported to interact functionally with S1P4 in ER-negative MDA-MB-453 breast cancer cells. S1P binding to S1P4 stimulates activation of ERK-1/2 and this is contingent on HER2. 95 S1P4 is linked with poor prognosis in ER-negative breast cancer patients as evidenced by shorter disease-free survival and disease-specific survival of patients who have high S1P4 expression in their tumors compared with those that have low receptor expression. 96 These reports implicate the important roles of SphK1 and S1P4 in prognosis of ER-negative breast cancer.
Potentiation of EGFR and type 1 insulin-like growth factor receptor (IGFR1) signaling by IGF-binding protein-3 (IGFBP-3) requires SphK1 activity and S1P1 and S1P3, where IGFBP-3 stimulates expression of SphK1 in breast epithelial cells, 97 which implicate that SphK1 may be a critical component of EGFR signaling in breast cancer progression as well. This has important implications as EGFR expression is associated with poor prognosis in breast cancer and correlates with lack of response to anti-estrogen therapy.
Compound Targeting S1P Signaling Pathway, FTY720
Interest in the functions of S1P in the immune system has recently increased exponentially due to the development of FTY720 for the treatment of multiple sclerosis. 13 FTY720 is derived from the myriocin (ISP-1) metabolite of the fungus Isaria sinclairii, and is a structural analogue of sphingosine. It is a pro-drug that is phosphorylated by SphK2 in vivo to a S1P mimetic, phospholylated-FTY720 (FTY720-P), which induces internalization and degradation of the S1P1 receptor and induces its prolonged downregulation 50 (Fig. 1). FTY720 sequesters lymphocytes in lymph nodes, preventing them from contributing to an immune reaction. FTY720 (fingolimod, trade name Gilenya, Novartis) is now approved by the United States Food and Drug Administration (FDA) for the treatment of patients with relapsing multiple sclerosis. It reduces the rate of relapses in relapsing-remitting multiple sclerosis by over half, but has adverse effects such as bradycardia, skin cancer, and potentially fatal infections. FTY720 has recently been investigated as a candidate therapeutic drug for the treatment of heart failure and arrhythmias as well. 98 Due to its immunosuppressive property, FTY720 was evaluated as an immunosuppressant in phase III clinical trial of kidney transplantation. However, it was found to be no better than the existing standard of care. 99
FTY720 has not been used in humans to treat cancer. It was reported that FTY720 inhibits the growth of breast cancer both in vitro and in vivo.100,101 FTY720 also demonstrated a strong anti-angiogenic effect and functional antagonism of vascular S1P receptors.102,103 Recently, oral administration of FTY720 in combination with sunitinib was shown to greatly decrease breast tumor growth with normalization of the reducing tumor angiogenesis in a rat breast cancer model, which suggests that this combination may improve the delivery of other chemotherapies. 104
S1P and Tumor-Induced Lymphangiogenesis
Angiogenesis is a crucial component of the growth and metastasis of tumors. Recent growing evidence implicates S1P as one of the most potent pro-angiogenic factors. Expression of S1P1 is upregulated in the tumor vasculature during angiogenesis, and downregulation of its expression was effective in inhibiting angiogenesis and tumor growth in vivo, suggesting that S1P1 receptor is also a critical component of the tumor angiogenic response. 53 Signaling via S1P2 negatively regulates S1P-mediated multiple responses of vascular endothelial cells. 105 The defective vascular maturation observed in S1P1-deficient mice highlights a fundamental role for S1P signaling on vasculogenesis. 51 Neutralization of the action of extracellular S1P shows significant inhibition of angiogenesis, tumor growth, metastasis, and lymphocyte transmigration, indicating that S1P is an important pathological regulator of inflammation and angiogenesis.27,102,106,107
Applying approaches similar to the ones used to understand angiogenesis is expected to identify molecular mechanisms that control lymphangiogenesis. 108 Interestingly, lymphatic endothelial cell (LEC)-specific deletion of SphK1 in the SphK2 knockout mouse inhibited lymphatic vessel maturation. 77 Taken together, SphK1 and S1P in LECs are required for the proper development of lymphatic vessels. 77
S1P stimulates lymphangiogenesis in both in vitro and in vivo models.12,109,110 S1P has been shown to induce in vitro lymphangiogenesis via the S1P1 receptor expression in LEC. 109 Tumor cells, which are characterized by high levels of SphK1 expression, can release S1P into the extracellular space, 13 which in turn can lead to paracrine-induced angiogenesis and lymphangiogenesis. 110 Yoon et al. found that S1P induced intracellular Ca2+ mobilization and formation of actin stress fibers in a pertussis toxin/phospholipase C-dependent manner in human lymphatic endothelial cells. 109 Anelli et al. showed that S1P can induce in vitro endothelial tube formation and cell migration in endothelial cell lines derived from vascular and lymphatic systems at nanomolar concentrations. 110
More recently, our group published that S1P generated by SphK1 is important not only for tumor progression but also for tumor-induced angiogenesis and lymphangiogenesis, therefore, targeting SphK1 and its product S1P would be a multipronged attack against breast cancer. 12 Previous studies demonstrated that SphK1 is overexpressed in breast cancer and its level of expression correlates with resistance to treatments and poor patient outcomes.14,23,44 However, several factors have hindered progress in determining the role of S1P in tumorigenesis and tumor-induced angiogenesis/lymphangiogenesis in vivo. Since S1P has such a profound role in immune function, 15 studies of traditional in vivo metastatic breast cancer models that utilize xenografts of human breast cancer into immune compromised mice that ignore the host immune response to cancer was deemed an inappropriate model to interpret the role of S1P in cancer biology. Thus, we utilized an improved syngeneic breast cancer cell implantation method, which mimics human breast cancer biology better than conventional xenograft subcutaneous implantation, to explore the role of SphK1 and S1P in angiogenesis and lymphangiogenesis. We have found that this chest orthotopic model readily develops lymph node metastases, which more accurately recapitulates human breast cancer progression than subcutaneous models. 12
Tumor-induced angiogenesis or lymphangiogenesis is usually evaluated by histological determinations of microvessel density or lymphatic vessel density, which rely on selective morphometric analysis (eg, vessel counts, vascular morphology, etc). 3 The strengths of the morphological approach are that it can evaluate the location of the vessels in relationship to the tumor and identify/quantify morphological changes that lymphatic vessels undergo during tumor progression. The limitations include variable sites of tissue sectioning, variable immunostaining techniques, different vessel density quantification methods, and the lack of standardization in the estimation of angiogenesis and lymphangiogenesis. 3 To complement this approach and overcome some of these limitations, we used a flow cytometry method to quantify both blood endothelial cells (BECs) and LECs in the same sample to evaluate both angiogenesis and lymphangiogenesis simultaneously. Flow cytometric analysis can provide supportive data to quantify the changes in BECs and LECs.
Directed in vivo angiogenesis assays (DIVAA) provide a simple and quantitative method to measure angiogenesis in vivo. 111 In contrast, one of the challenges of studying lymphangiogenesis in vivo is the lack of assays for objective quantification. 3 To this end, we developed a similar approach to quantify both angiogenesis and lymphangiogenesis simultaneously by combining DIVAA with fluorescence activated cell sorting (DIVAA/FACS). 12
Utilizing the animal model described above, we found not only that circulating S1P levels correlated with tumor burden, but also targeting SphK1 with a specific inhibitor, SK1-I, reduced tumor growth, lymph node and lung metastasis. 12 Consistent with both the in vitro tube formation and DIVAA/FACS assays, we demonstrated that exogenous S1P enhanced angiogenesis and lymphangiogenesis while SK1-I inhibited each process. Furthermore, pharmacological inhibition of SphK1 with SK1-I decreased angiogenesis and lymphangiogenesis not only around the primary tumor, but also in lymph nodes that are distant from the tumor. This result indicates that S1P play a key role not only in “tumoral lymphangiogenesis”, but also in “lymph node lymphangiogenesis”, which we speculate to promote metastasis actively via the lymphatics. Based upon our findings, we believe that targeting SphK1 and S1P signaling pathways may be a useful additional modality for treatment of breast cancer by suppression of tumor-induced lymphangiogenesis progression.
Conclusions
In conclusion, S1P generated by SphK1 is important not only for tumor progression but also for tumor-induced lymphangiogenesis and lymph node metastasis. Targeting S1P signaling pathway is expected to suppress tumor-induced lymphangiogenesis, thus, breast cancer progression.
Footnotes
Author Disclosure Statement
Kazuaki Takabe is supported by United States National Institute of Health Grants R01CA160688 and K12HD055881, and by Susan G. Komen for the Cure (Investigator Initiated Research Grant). Masayuki Nagahashi is a Japan Society for the Promotion of Science Postdoctoral Fellow.
No competing financial interests exist.