
Abstract
Abstract
Background:
The present work deals with innervation patterns along collector lymphatic vessels from cervical, mesenteric, and femoral regions, and lymph capillaries in young and elderly subjects.
Methods and Results:
Morphological and morphometric analysis of nerve fibers along lymph vessels was performed by immunohistochemistry for PGP 9.5, NPY, TH, ChAT, VIP, SP, and dopamine. Nerves containing NPY and TH were frequent, whereas immunoreactivity for ChAT and VIP were few. SP-positive fibers were widely distributed in the medial and endothelial layers. Dopamine neurotransmitters were observed in a few short nerve fibers. A more diffuse presence of nerve fibers in mesenteric and femoral lymph vessels, compared to cervical ones, was detected. In lymph capillary vessels, a few nerve fibers positive for neuropeptides and neurotransmitters were detected, whereas no dopamine and VIP immunoreactive fibers were detected. A wide reduction of all specific nerve fibers analyzed was detected in lymph vessels from elderly subjects.
Conclusions:
The presence on lymph vessels of sympathetic and parasympathetic nerve systems can be declared. The differences observed in lymphatic vessel innervation patterns may note the involvement in lymph flow regulation, calling attention in aging, when nerve fibers reduction may cause functional default of lymph vessels.
Introduction
The absence of a central pulsating organ in the lymphatic system makes lymph drainage in lymphatic vessels rather enigmatic. Initially an extrinsic or passive mechanism was proposed, consisting in the transmission of mechanical tissue forces elicited by the lymph formation rate (as a consequence of pressure gradient produced by cardiac or respiratory displacements), pulsating vessels, and/or skeletal muscle contraction acting on the lymphatic vessels (passive lymph progression).1,2 Now an intrinsic mechanism was also proposed that consists of spontaneous contraction of lymphatic vessels capable of inducing fluid propulsion (intrinsic activity or active lymph progression).3,4 Contractions in lymphatic vessels are started by action potentials that originate in pacemaker cells residing within the muscle layer, 5 even if the nature of these pacemaker cells is still unclear.6,7
On the basis of functional studies on lymph flow in various body regions, differences have been found that must take into account the different responses to local lymph production rate and drainage. 8 Therefore, the lymph formation rate appears to be another important force in determining pressure gradient along lymph vessels. In summary, lymph flow is the result of a combination of several local driving forces.
Considerable evidence suggests that lymphatic vessels have a nerve supply.9,10 These nerve fibers are able to produce an increase in the frequency and strength of lymphatic contraction.11–13 Vagal cholinergic nerves of lymphatic vessels also have shown experimental evidence as an important factor for active lymph flow. 14 Furthermore, humoral factors and neuropeptides also act as modulators of the lymphatic tone and contractility and thus of the lymphatic functions.1,15–17 The complex regulation of lymph flow may make modulation of lymph vessel nerve fibers important in assisting lymphatic vessels functionality. 10 Neuropeptides are well-known modulators of the central and peripheral nervous systems. In particular, neuropeptides such as NPY and VIP are recognized markers of sympathetic and parasympathetic nerve fibers, respectively.18,19 We previously analyzed, in young and old men, some neuropeptides and neurotransmitters localized in nerve fibers running in human thoracic duct, the final collector of lymphatic vascular system. 20 In the present study, we extend the analysis to human collector lymphatic vessels taken from different body regions and lymph capillaries. Similarly, the differences of morphometric distribution of nerve fibers has been analyzed among young and old men.
Materials and Methods
Ethics
All the experiments performed in this work were previously approved by the local Ethical Committee. All experimental procedures were conducted in accordance with the Declaration of Helsinki in its revised edition, in the Guidelines of Good Clinical Practice (CPMP/ICH/135/95), and international and local regulatory requirements. To each sample was assigned a progressive number followed by a letter indicative of the participating unit. For each case, a report was prepared indicating age sex and other general notices of each single subject. The anatomical units (Camerino, Sapienza, and Novara) knew only the number and letter of each sample. Therefore, the so-called procedure of the “double blind” was used in all experiments.
Samples
Specimens were obtained from autopsies on 16 dead subjects and immediately processed. Eight male subjects (age range 28–35 years) were considered for the young group; 8 male subjects (age range 68–75 years) were considered for the elderly group. All subjects were free of any disease.
For each subject, samples were harvested from different regions of the lymphatic system: cervical, mesenteric, and femoral lymphatic collectors. All lymphatic collector vessels were initially identified as vessels linked to regional lymph nodes. Moreover, peribronchiolar connective tissue was chosen for analysis of lymph capillaries, due to easy contrast between blood capillaries bringing erythrocytes into lumen and lymph capillaries without them. For lymph collectors and capillaries, the morphological criterion was used together with anti-LYVE-1 immunohistochemistry to identify them as lymphatic vessels. 21 Three random choice collector vessels from each region were taken under exam. Each sample was rapidly washed in cold PBS. Then vessel samples were divided into halves, one oriented along the transversal plane, the other oriented along the coronal plane. All specimens were embedded in a cryoprotectant medium and frozen in isopentane cooled with liquid nitrogen. Serial sections (25 μm thick) were cut on a cryostat at −20°C. Owing to the considerable thickness of the slides, immunohistochemical experiments were performed by the free-floating method.
The first section of each specimen (collector vessels and peribronchiolar tissue) was stained for the immunohistochemical detection of nerve fiber profiles using a general marker for nervous tissue, PGP-9.5 (rabbit polyclonal, diluted 1:600 in PBS; Cambridge Research Biochemicals, UK, code ABIN704396).
The second section was stained for the immunohistochemical detection of TH to visualize catecholaminergic nerve fibers (mouse monoclonal, dilute 1:10 in PBS, Novocastra Laboratory, UK, code NCL-TH).
The third section was stained for the immunohistochemical detection of ChAT to visualize cholinergic nerve fibers (goat polyclonal, diluted 1:30 in PBS, Chemicon, CA, code AB144P).
The fourth section was processed for immunohistochemical detection of VIP (rabbit polyclonal, diluted 1:1000 in PBS; Peninsula Laboratories, San Carlos, CA, code T4246).
The fifth section was stained for immunohistochemical detection of NPY (rabbit polyclonal, diluted 1:600 in PBS; Cambridge Research Biochemicals, code ABIN112908).
The sixth section was stained for the immunohistochemical detection of SP (rabbit polyclonal, diluted 1:400 in PBS; Cambridge Research Biochemicals, code ABIN643829).
The seventh section was stained for the immunohistochemical detection of dopamine positive fibers (mouse monoclonal antibody, diluted 1:50; Abcam, Cambridge, UK, code ab8892).
To visualize the immune reaction, a secondary fluorescein isothiocyanate-conjugated antiserum was used (Vector Laboratories, group code FI-1000, Burlingame, CA). Furthermore, to better visualize the tissue detail, the slides from peribronchiolar connective tissue specimens were counterstained with DAPI. Control sections were processed as above, but using a nonimmune rabbit IgG instead of the primary antibody. No positive reaction was observed under these conditions.
The eighth section of coronal section collector vessels and peribronchiolar tissue was stained for LYVE-1 immunohistochemistry (rabbit polyclonal antibody, diluted 1:100; Abcam, Cambridge, code ab10278). It was visualized by anti-rabbit biotinylated secondary antibody and avidin/biotinylated enzyme complex (Vectastaine Elite kit, code PK6101, Vector Laboratories, Burlingame, CA) together with DAB as chromogen (DAB chromogen kit, Vector Laboratories).
The ninth slide of coronal section collector vessels and peribronchiolar tissue was processed for hematoxylin and eosin staining to assess general microanatomy.
Nerves fiber morphometry
Qualitative morphological display evaluation of nerve fibers along collector lymphatic vessels was performed on coronal and sagittal sections.
Quantitative morphometric analysis of nerve fibers along collector lymphatic vessels was performed on sagittal sections only, because the long fibers observable provided a better reliability of morphometric analysis. In sagittal specimens, a total thickness of 175 μm were analyzed for each vessel, highly compatible with the total vessels wall thickness (∼240 μm). In this way, we performed morphometric analysis exclusively inside the vessel wall. For each slide, analysis was performed on 5 randomly fields (each field=100×103 μm2).
The stereological method for counting fibers was adopted in accordance with the evaluation of length fiber density (Lv) parameter. 22 In each field under analysis, Lv represents the density of nerve profile per unit volume. The volume of analysis consisted in total area of analysis field×slides thickness (125×105 μm3).
A nerve profile was defined as an immunopositive segment seen regardless of its size and length (isotropic distribution). The shift of focus plane was used to better observe the several nerve profile included into volume of analysis. Experimental data were obtained using following formula: Lv=Q/V, where Q represents the number of immunopositive nerve profiles, and V represents the volume of analysis. Morphometric analysis of nerve fibers profile along lymphatic capillaries was performed similarly to the above methodology, but, owing the absence of vessel wall, adopting specific expedients. In each slides, three capillary profiles were chosen on which a part of longitudinal profile was appreciable (≥5×103 μm2). The LA parameter was evaluated as LA=Q/A, where Q represents the number of immunopositive nerve profiles, and A represents the area of analysis of each capillary.
The nerve fiber branching profile along collector lymph vessels and capillaries was detected by fluorescence microscope connected to a digital camera, using image analysis computer software (Qwin, Leica).
Data analysis
Data are presented as mean±S.E.M. calculated from values detected in the individual samples of each experimental group. The normal distribution of nerve fiber data were assessed by means of the Kolgomorov-Smirnov test. Statistical differences among experimental groups for different regions of lymphatic system were then assessed by the ANOVA test, followed by the Newman-Keuls post hoc test. Statistical procedures were performed with the Prism 4 statistical software (GraphPad Software Inc., CA). The level of significance was p<0.05.
Results
Collector lymphatic vessels
All collector lymphatic vessels were stained positively to LYVE-1 immunohistochemistry. Examining coronal sections, any differences were observed in the morphological pattern of the nerve fibers distributed in lymphatic collectors from the various regions of the human body, both in young and elderly subjects. The microanatomical structure of lymphatic collectors (with a diameter ∼200 μm) was identified as composed by an external connective, thin layer, a medial layer formed by smooth muscle cells, and the internal layer formed by endothelial cells. In young subjects, the PGP 9.5 immune positive nerve fibers, running in perivascular connective tissues, reach the lymphatic vessels from the external layer and are distributed through the intermedial layer in perpendicular and transversal orientation in respect to the longitudinal axis. These nerve fibers are frequently connected by longitudinally or obliquely orientated interconnections. Some fibers also seemed to reach the internal layer of the lymphatic vessel wall (Fig. 1A). In the external layer, immune-positive nerve fibers were noted following TH (Fig. 1B) and ChAT immunohistochemistry; few fibers were noted after VIP and SP immunohistochemistry. In the medial layer of vessels, after NPY, ChAT, TH, and SP immunohistochemistry, positive fibers were noted, displayed similarly to the PGP9.5 positive ones. In particular, TH and SP immunohistochemistry revealed less interconnections than NPY and ChAT immunohistochemistry. Furthermore, NPY, TH, and SP positive fibers were noted to reach the internal layer of vessel, TH and SP positive fibers displayed occasional dot spot signals into the internal layer (Fig. 1B). Instead following VIP and dopamine Immunohistochemistry, rare and short positive nerve fibers were observed perpendicularly oriented, more rarely transversally oriented. Thick fibers (∼12 μm thick) were the VIP positive fibers, thin fibers (≤7 μm thick) were the dopamine positive ones. Both types of these positive nerve fibers did not reach the vessel internal layer (Fig. 1C and 1D). No differences were detected in the immunohistochemical pattern of specific nerve fibers in coronal lymph vessels among young and elderly subjects (data not shown).

Microphotograph showing immune positive nerve fibers in transverse sections of lymphatic vessel mesenteric collector from young patients. Immunochemistry for
Analyzing sagittal sections, any differences in the nerve fibers morphological pattern were observed among lymphatic vessels from the various regions of the human body, both in young and elderly subjects. Moreover, immune staining for the general neural marker PGP 9.5 shows that nerve fibers reach lymphatic vessels and run as numerous thick (∼12 μm thick) and thin (≤7 μm thick) immune positive fibers along the wall of same vessels, interconnected, and displaying a dense fiber network appearance.
TH and NPY immunohistochemistry revealed a dense fiber network, characterized by thick immune positive nerve fibers (Fig. 2A–2C), similarly SP Immunohistochemistry showed thin immune-positive fibers (Fig. 2D–2F). Instead, a less dense fiber network appearance was observed after ChAT immunohistochemistry, characterized by immune positive thick nerve fibers (Fig. 2G–2I). Longitudinal immune positive nerve fibers with dot spot appearance were observed following VIP immunohistochemistry (Fig. 2L–2N); following dopamine immunohistochemistry, transversal and/or longitudinal short immune positive nerve fibers were noted, not arranged in a clear dense fiber network appearance. In elderly subjects, the display of the immune positive fiber profile was similar to that of young subjects.
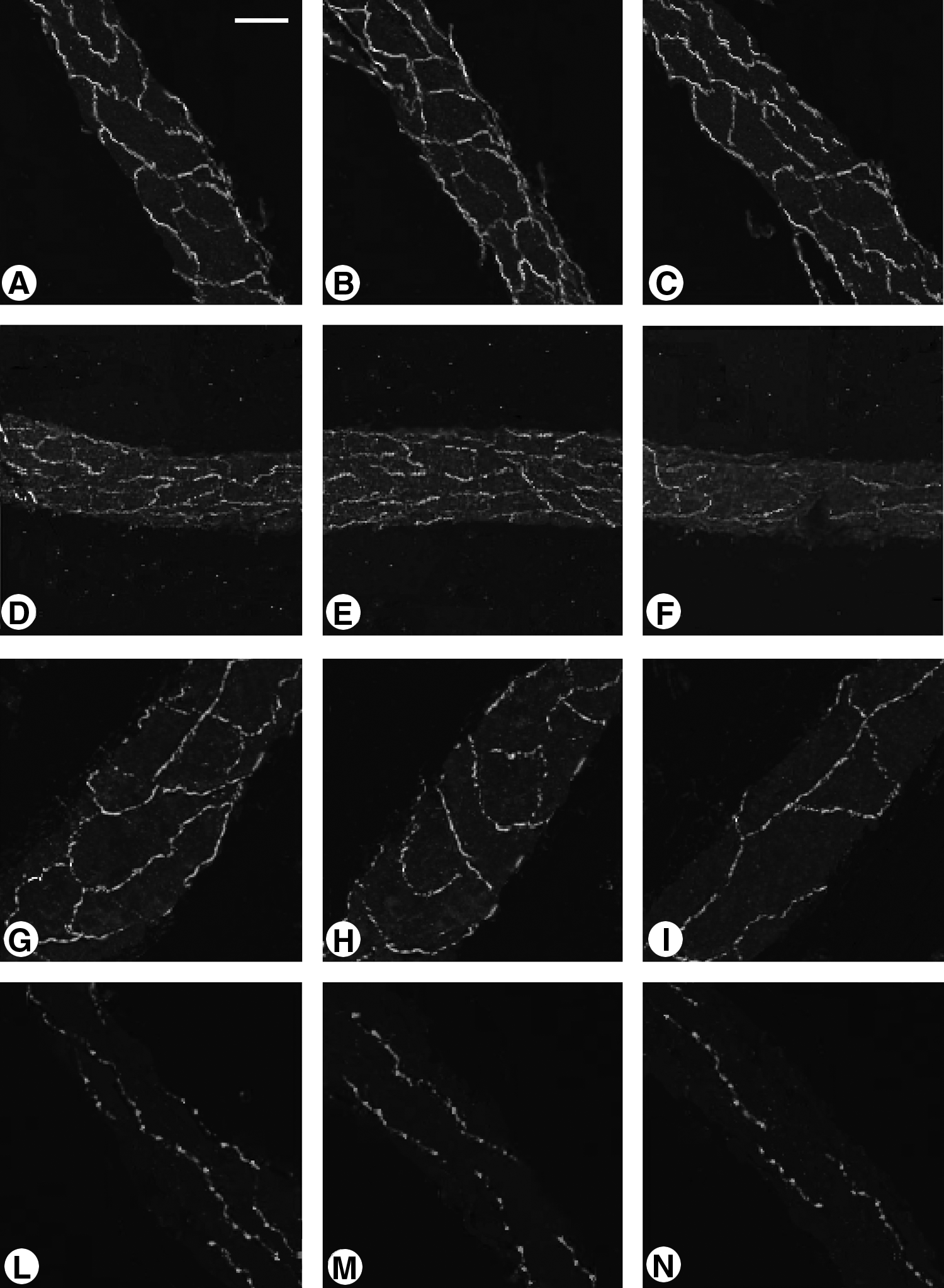
Microphotograph showing comparison of several immune positive nerve fibers in longitudinal sections of lymphatic vessel collectors (l.v.) taken from cervical, mesenteric, and femoral body zones of young and old subjects (s.).
Quantitative analysis of immunohistochemical positive nerve fibers detected in young subjects showed a significant higher number of PGP 9.5 positive fibers in mesenteric and femoral lymphatic vessels than in cervical ones (Fig. 3A). The analysis of specific immunohistochemical signals showed that the differences were due to a higher number of all specific positive nerve fibers detected, with the exception of SP positive ones. No significant differences were detected between mesenteric and femoral lymphatic vessels in young subjects (Fig. 3A). In the elderly subjects, less nerve fibers were detected for all different immune positive fibers analyzed in all regionally different lymphatic vessels (Fig. 3B).
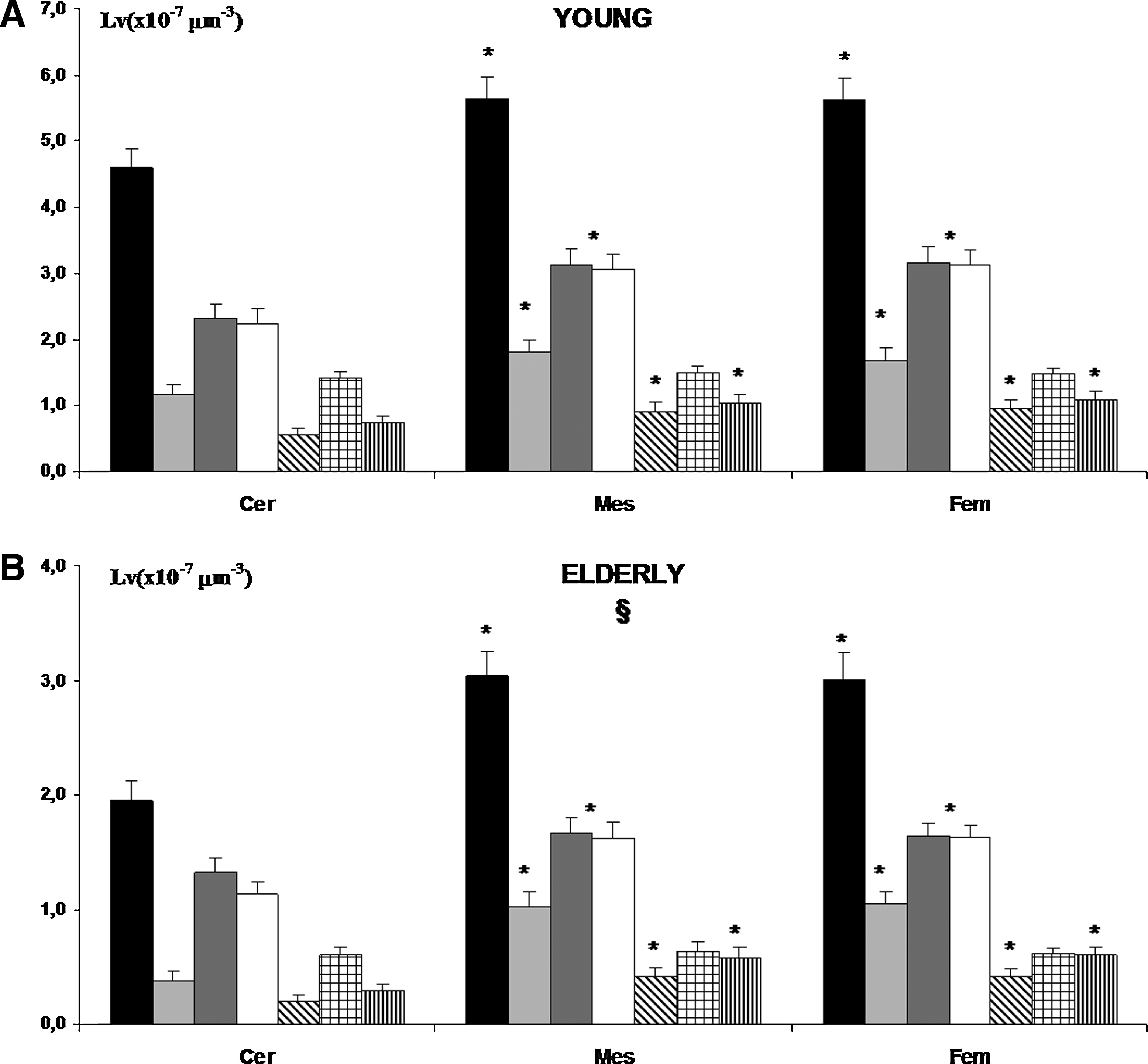
Histogram showing morphometric evaluation of several immune positive nerve fibers (p.f.) detected in lymphatic vessel collectors taken from cervical (Cer), mesenteric (Mes), and femoral (Fem) body zones of young and old subjects. Black bar, PGP 9.5 p.f.; light gray bar, ChAT p.f.; dark gray bar, TH p.f.; white bar, NPY p.f.; diagonal band bar, VIP p.f.; square bar, SP p.f.; vertical band bar, dopamine p.f. *p<0.05 vs. Cer; in B §p<0.05 elderly vs. young.
Lymphatic capillaries
Lymphatic capillaries were morphologically identified as a large capillary profile where no blood cells can be observed and immunohistochemically positive to LYVE-1 (Fig. 4A). In capillaries from both young and elderly subjects, no VIP-positive and/or dopamine-positive nerve fibers were observed. On the contrary, longitudinal nerve fibers running along the capillary wall were observed positive for the other neurotransmitters studied (Fig. 4B and 4C). In elderly subjects, a significant decrease of positive fibers were detected (Fig. 5).

Microphotograph showing immune positive nerve fibers in the lymphatic capillaries from peribronchial connective tissue in transverse sections.
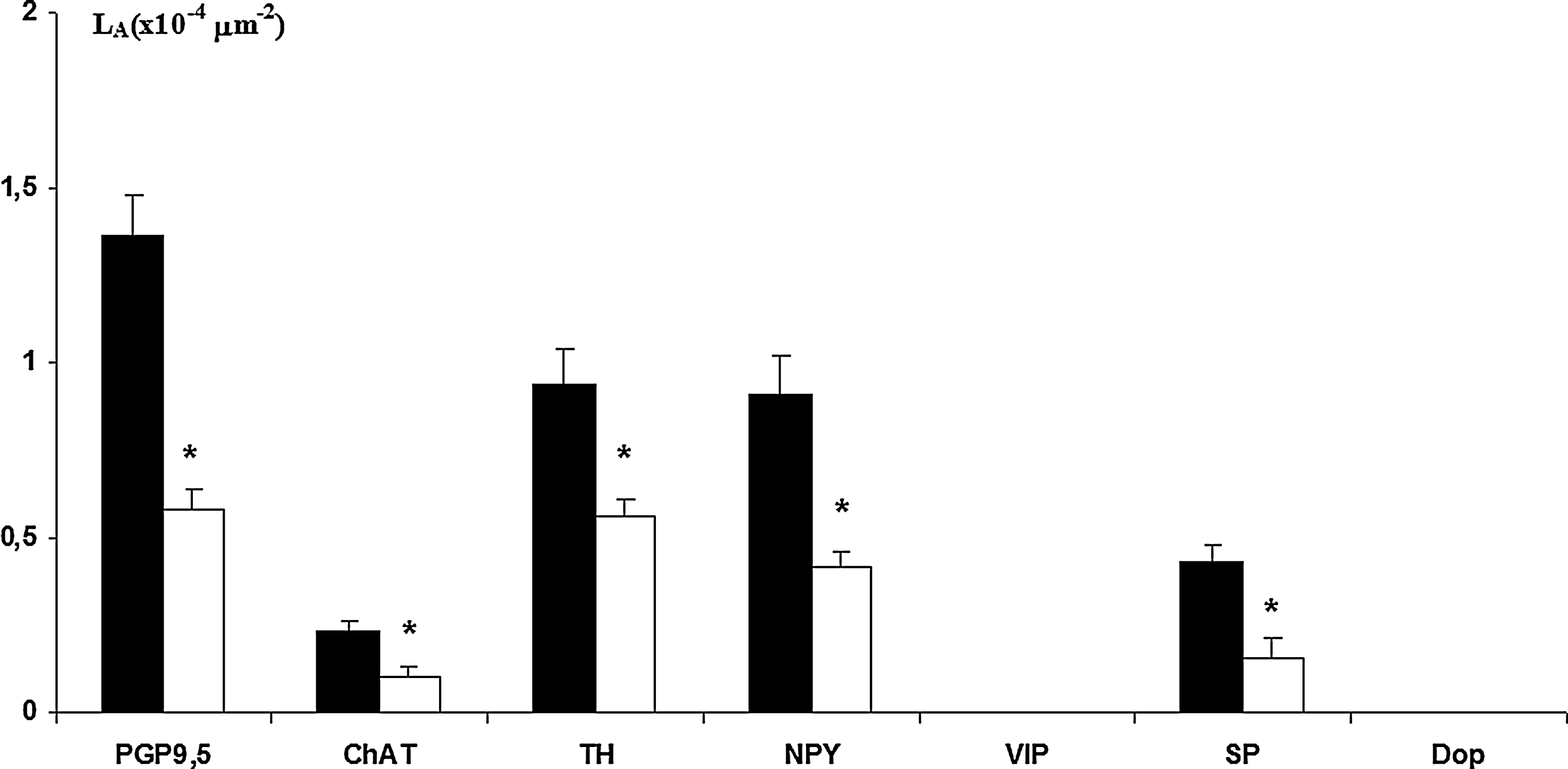
Histogram showing morphometric evaluation of several immune positive nerve fibers detected in lymphatic capillaries of young (■) and elderly (□) subjects. *p<0.05 vs. young. VIP and dopamine were not detected.
Conclusion
In our study, the PGP9.5 marker provided a clear immunocytochemical evidence of well-characterized populations of nerve fibers running around collector and lymphatic capillaries in humans. PGP9.5 is a soluble protein present in nervous and neuronal and endocrine cells. It has been identified as ubiquitin carboxy-terminal hydrolase and has been observed involved in axonal transport physiology. 23 Its immunohistochemical evidence is particularly useful for identifying small diameter unmyelinated neurons and/or investigation of nerve fibers. 24 It may be of interest to note that the disappearance of PGP9.5 revealed a functional default of nerve fibers not necessarily followed by an anatomical disappeared of fibers themselves.
In the present work, a specific distribution of neurotransmitters contained in lymphatic nerve fibers has been described, suggesting their specific involvement in regulating the capacity of the lymphatic pumping activity. It was shown that nitric oxide formation represents the major regulatory component of lymphatic pumping, 25 but also that hydrodynamic factors such as stretch/pressure and shear/flow play crucial roles in modulating lymphatic pump function. 4
In this context, the lymph vessel innervation may be seen as a modulator system that assists lymph pump activity and better coordinates the lymph function with the physiological status of the body. Otherwise, orthostatic position, body movement, visceral movements, and the lacteal lymph postprandial production are all conditions responsible for influencing lymph flow, that in different body regional district satisfy different physiological needs.
In collector and capillary nerve lymph vessels, fibers containing NPY and/or TH were the most frequent; they showed a similar distribution pattern, consistent with identification of postganglionic sympathetic nature of these nerve fibers. 15 Immune reactivity for ChAT and VIP, markers of cholinergic parasympathetic nerve fibers, 26 was evident but sparse. Therefore, the influence of the sympathetic and parasympathetic nervous systems on the collector lymphatic vessels and lymphatic capillary in humans can be confirmed.
NPY is a modulator of TH, the main transmitters of the sympathetic nerve fibers inducing vessels contraction. 27 We have observed nerve fibers containing these modulators to represent the largest amount of lymph vessels innervating fibers both in young and elderly subjects; these findings may indicate a prevalence of contractile tone modulation offered by lymph vessels innervation. With this point of view, the occurrence of dopamine may be also considered; in fact, dopamine in visceral and vascular nerve fibers was seen strictly associated with NPY and TH modulators.28,29 The appearance of dopamine in short fibers may suggest its content in nerve varicosity with neurotransmitters releasing function (boutons en passant) such as observed in other organs.30,31
SP was the most represented neuropeptide in collectors and capillaries lymph vessels after NPY. SP is a vasodilator in blood vessels, 32 but in lymph vessels, it was observed to exert a substantial positive inotropic and chronotropic effects improving pump efficiency,17,33 and its specific involvement in the lymph vessels mechanical response during inflammation has been documented. 34 These findings are well in accord with our morphological observation that SP nerve fibers run widely along the contractile wall of the lymph collectors.
VIP is the main neurotransmitter located in the parasympathetic nerve fibers and represents the main modulator of cholinergic synapses; it induces smooth muscle relaxant activity.9,18 Indeed, VIP is very low represented in lymphatic collector nerve fibers such as ChAT. We hypothesize that these cholinergic nerve fibers are involved rather in modulating the elastic performance of lymph vessels, favoring the relaxing status of smooth muscle cells. 35
The occurrence of SP and TH positive nerve fibers in the internal layer of lymph collectors may indicate their sensorial function. Indeed, the sensorial activity of subendothelial SP positive fibers in lymph vessels was already documented in previous works;9,36 and a role for NPY as modulator of various immune functions and lymphocytes migration has been also observed.29,37
The higher distribution of nerve fibers in mesenteric and femoral lymph vessels with respect to cervical ones allow us to hypothesize an important involvement of vessel innervation in lymph flow regulation of body lower regions, where antigravity force action is more evident, and may generate the need to assist the lymph pump activity in a more complicated way.
Indeed, aging seems to affect differently the lymph pump functions in the several regions investigated. The relaxant tone may be mediated by ChAT and co-modulator VIP, and was mainly affected by aging in cervical collector lymphatic vessels (ΔLV∼−67%), in comparison to contractile tone as may be mediated by TH and its co-modulators such as NPY, SP, and dopamine, (ΔLV∼−42%). On the contrary, it has been observed in femoral lymphatic vessels (dilatory nerve fibers ΔLV∼−37%; contractile nerve fibers ΔLV∼−48%), while in mesenteric lymphatic vessels, the change was similar between relaxant and contractile nerve fibers (ΔLV∼−44%). These findings indicate an imbalance in lymph pump modulation activity with consequent severe impairment of lymph flow progression in elderly subjects.
ChAT-, TH-, NPY- and SP-positive nerve fibers were found also running along lymph capillaries profile. Here the absence of dopamine and VIP positive nerve fibers, supposed as modulators of contractile/relaxant response of smooth muscle cells of lymph collectors, may follow the absence of smooth muscle cells.
The fundamental physiologic work by lymph capillaries consists of the trans-endothelial permeability to lymph flow and leukocytes transit. Our morphological findings may support the hypothesis of a role of nerve fibers located along lymph capillaries on leukocytes transit. In fact, in leukocytes, neurotransmitters and neuropeptides receptors are well represented; 38 furthermore, innervation might be involved in sensorial functions on shear/flow hydrodynamic factors to modulate lymph drainage during several physiological needs.
The age-related changes affect the lymph vessels and induce a decrease of the number of its nerve fibers. Following the supposed role of these nerve fibers, we may argue about a functional default of capillaries functionality in elderly subjects that may impair not only the tissue lymphatic drainage but also the lymphocyte circulation.
In our previous work, 20 the pattern of the same neuropeptides and neurotransmitters studied in the present one, were analyzed in thoracic ducts resected from the same subjects used in the present work. In the thoracic duct, TH, NPY, and SP fibers were also observed as the most represented fibers. We described a quantitative decrease of all nerve fibers in thoracic duct of old men, with the ChAT positive fibers the most affected. Thoracic duct also revealed a prevalent impairment of relaxing tone such as observed in collector vessels. All these findings may lead to the hypothesis of a functional stiffness of lymphatic vessels in elderly subjects. Our studies on innervation patterns of the lymphatic vascular system are shown in Table 1.
Data are presented as Mean±SEM, all values presented in Old are statistically significant vs. Young ones. TD, Thoracic Duct (data from Mignini et al. 20 ); CerLv, cervical lymphatic collectors; FemLC, remoral lymphatic collectors; LCp, lymphatic capillaries; MesLC, mesenteric lymphatic collectors.
Our findings show a rich innervation of the vascular lymphatic system in which several neurotransmitters and neuropeptides are present. In this context, lymph innervation may support the hydrodynamic forces and humoral factors known to be the fundamental regulatory factors of lymph function. Otherwise, important regional variations in the hydrodynamic sensitivities of lymph function have been observed. We argue that innervation may have a function of linking and coordinating the several local responses of lymphatic function, and linking them with the physiological status of the body. An alteration of this complicated lymphatic function may have serious consequences on physiological homeostasis of the body; therefore, the reduction of innervation in aging calls attention to a compromising of lymphatic function that may be the origin of pathology or worsen existent pathology.
Footnotes
Author Disclosure Statement
No competing financial interests exist for any author.