
Abstract
Abstract
LYVE-1 is a marker expressed by lymphatic endothelial cells (LECs) that line the lymphatic endothelium. Through studies designed to examine potential changes in expression of LYVE-1 in cynomolgus macaque colon tissues during the course of simian immunodeficiency virus (SIV) infection, we discovered that LYVE-1 was expressed by heterogenous populations of cells. As revealed by in situ hybridization (ISH), LYVE-1 mRNA levels in colon were decreased in macaques with AIDS compared with acutely infected or uninfected macaques. In the submucosal layer of the colon, approximately half of the LYVE-1-expressing cells co-expressed the dendritic cell (DC) marker, DC-SIGN/CD209, and this percentage did not change appreciably during infection. Subsets of cells expressing LYVE-1 also co-expressed macrophage markers, such as CD68 and the macrophage mannose receptor (MMR)/CD206, in both the colon and lymph nodes. LECs, DCs, and macrophages that co-expressed LYVE-1 were observed in colon and lymph node from uninfected, healthy animals as well as in tissues with SIV-driven inflammation. These findings provide further definition of the phenotypic overlap between LECs and antigen presenting cells, reveal the heterogeneity within the population of cells expressing the lymphatic marker LYVE-1, and show that SIV modulates this population of cells in a mucosal surface across which the virus is acquired.
Introduction
LYVE-1 is expressed by the efferent lymphatic vessels in the medulla of normal lymph node (LN), contributing to hyaluronan homeostatis and turnover. 11 It is related in sequence and function to CD44, and is functionally regulated by sialylation such as CD44, 12 but its set of functions is not fully understood, given that mice in which the lyve1 gene has been knocked out show no evident phenotype. 13 In intestinal tissues, central lacteals in the villi of the small intestine drain into the lymphatic vessels in the submucosa, and mucosal lymphatic vessels in the cecum and colon drain into the submucosal lymphatic vessels. The central lacteals and mucosal lymphatic vessels in the rat intestine express LYVE-1. 14 Despite the observation in a number of model systems that LYVE-1 is expressed by lymphatic endothelium, the phenotype and distribution of cells expressing LYVE-1 in normal and inflamed intestinal tissues has not been examined in detail. Differential expression of LEC markers has been observed in the LNs from cynomolgus macaques infected with simian immunodeficiency virus (SIV) and Mycobacterium tuberculosis. 15
A complex network of immune cells that includes dendritic cells (DC), macrophages, T cells, and LECs resides in the gastrointestinal tract. Recent reports have investigated the function,16,17 and subtypes18,19 of DCs and macrophages in the lamina propria. In our previous report, expression of the DC-specific ICAM3-grabbing non-integrin (DC-SIGN) was observed in the lamina propria and submucosal layer of macaque intestine, and the expression levels of DC-SIGN in intestine did not change appreciably during pathogenic simian immunodeficiency virus (SIV) infection. 20 Most studies of immune cells in the intestine have focused on cells localized to the lamina propria, as compared to the submucosal layer. Moreover, despite the likely importance of LECs in homeostatic, inflammatory, and infectious processes in intestinal tissues, the amount of information on LECs in the intestine is limited, especially during infections. In this study, we focused on the large intestine of macaques during the course of SIV infection because the colon is a likely site of microbial translocation 21 with associated systemic immune activation, and lower colon and rectum are major sites of mucosal transmission during HIV-1 and SIV infection.22,23 We have investigated the populations of cells expressing LYVE-1 in cynomolgus macaque colon as well as the effects of SIV infection on the levels and distribution of these cells. These studies revealed heterogeneity in the types of cells expressing LYVE-1 in colon and secondary lymphoid tissues.
Methods
Animals and tissues
The previous studies with macaques from which archival tissues were available were performed under the guidance and approval of the University of Pittsburgh Institutional Animal Care and Use Committee. Archival necropsy tissues from 18 adult cynomolgus macaques (Macaca fascicularis) were included in these studies. At study entry, the macaques were negative for SIV, simian retrovirus (type D), and simian T-lymphotropic viruses -1, -2, and -3. Macaques were infected via the intrarectal route with the SIV/DeltaB670 primary isolate as described previously.24,25 Tissues were obtained at necropsy from six naïve, SIV-uninfected macaques, six acutely (2 weeks post-infection [wk PI]) SIV-infected macaques, and six long-term (36–75 wk PI) SIV-infected macaques with acquired immunodeficiency syndrome (AIDS). Tissues were fixed by immersion in fresh 4% paraformaldehyde in phosphate buffed saline (PBS) and processed as described previously.20,25
In situ hybridization and immunohistochemical staining
The generation of the rhesus macaque LYVE-1 partial cDNA has been described. 15 In situ hybridization (ISH) with [ 35 S]-UTP-labeled riboprobes, immunohistochemistry (IHC), and combined ISH/IHC were performed as described.15,20,25 Autoradiographic exposure times were 20 days for LYVE-1. Antibodies used for combined ISH/IHC were specific for anti-human DC-SIGN (DCN46; BD Pharmingen, San Diego, CA) and anti-human LYVE-1 (AF2089; R&D Systems, Minneapolis, MN). The percentages of LYVE-1 mRNA+ cells that expressed DC-SIGN were determined by examining 100 LYVE-1 mRNA+ cells per microanatomic location at a magnification of 400X and categorizing each cell as DC-SIGN-positive or -negative.
Quantitation of ISH signals
For quantitation of the signals generated after ISH with 35 S-labeled probes and emulsion autoradiography, we used the approach described. 20 Briefly, the numbers of LYVE-1 mRNA+ cells were enumerated per 5.46 mm2 of tissue from each tissue section at a magnification of 400X in two independent experiments.
Immunofluorescence staining and confocal microscopy
Immunofluorescence staining of LYVE-1, DC-SIGN, CD68, and CD206 was performed in colon tissue sections. Cryosectioned specimens were incubated with goat anti-human LYVE-1 (R&D Systems) and either mouse anti-human DC-SIGN (DCN46; BD Pharmingen), mouse anti-human CD68 (KP-1; Dako Cytomation, Carpinteria, CA), mouse anti-human CD206 (555953; BD Pharmingen), or mouse anti-human podoplanin (11-003; AngioBio, Del Mar, CA) for 60 min, followed by washing three times in PBS. Sections then were incubated for 30 min with Alexa Fluor 488-conjugated donkey anti-mouse IgG (Invitrogen, Carlsbad, CA), and Alexa Fluor 647-conjugated rabbit anti-goat IgG (Invitrogen), followed by washing three times in PBS. A drop of Prolong Gold antifade reagent with 4',6-diamidino-2-phenylindole (DAPI; Invitrogen) was added and the coverslipped sections stored at 4°C until visualization. Negative control staining was performed by either omitting the primary antibodies or using isotype-matched control antibodies at similar concentrations. Tissue sections were examined using a FluoView FV1000 confocal microscope (Olympus, Tokyo, Japan) at the University of Pittsburgh's Center for Biologic Imaging.
Statistical analysis
Significant differences between the groups were evaluated using an unpaired Student's t-test. Statistical calculations and analyses were performed with the use of Prism 5 software (GraphPad Software, San Diego, CA). P values <0.05 were considered significant.
Results
SIV infection alters levels of expression of LYVE-1 mRNA in the macaque colon
To determine whether in colon, the levels of expression of LYVE-1 change during the course of SIV infection, LYVE-1 mRNA+ cells were identified by ISH and their numbers were enumerated in three microanatomic regions of colon, including the epithelium/lamina propria, submucosa, and muscle layers. The tissues examined were collected at necropsy from cynomolgus macaques that were either uninfected, or infected intrarectally with the SIV/DeltaB670 pathogenic strain 26 and sacrificed during acute infection (2 weeks post-infection [wk PI]) or upon development of AIDS (Table 1). Cells expressing LYVE-1 mRNA (Fig. 1A) and protein (Fig. 1B and 1C), were heterogenous and included vascular-associated and nonvascular cells. As a specificity and sensitivity control, we performed ISH for LYVE-1 mRNA and IHC for LYVE-1 protein simultaneously on multiple macaque colon sections and found that nearly all cells staining by one method also stained positive by the other method (not shown). LYVE-1 mRNA+ cells were more abundant (per mm2) in the submucosa (8.1-fold; p<0.05) and muscle layer (6.2-fold; p<0.05) than in the epithelium/lamina propria, regardless of infection status (Fig. 1D–F; note y-axis scale differences). These findings indicate that the extensive network of LYVE-1-expressing cells is mainly located in the submucosa and muscle layers of the large intestine. Upon development of AIDS, the densities of LYVE-1 mRNA+ cells in the lamina propria and muscle layers of the colon decreased significantly (P<0.05) in comparison to acutely infected macaques and uninfected macaques (Figs. 1D and 1F). The numbers of LYVE-1 mRNA+ cells in the submucosal layer of colon decreased during AIDS, but the differences were not significant. As expected, and as a control for detection of LYVE-1+ LECs, LYVE-1 mRNA+ cells were located in the central lacteals of the jejunum in all animals examined, regardless of SIV disease state (not shown).
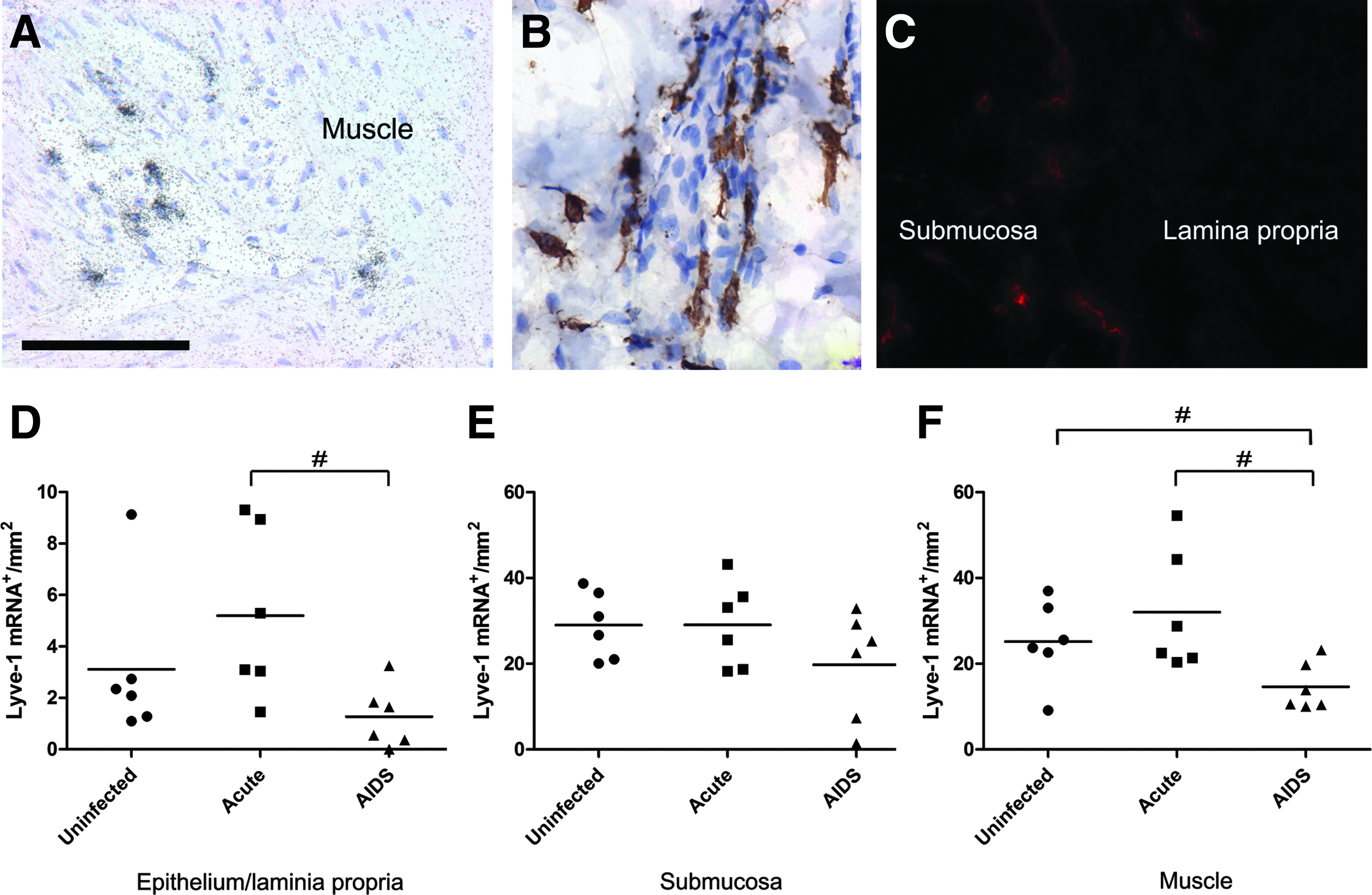
LYVE-1 mRNA expression in macaque colon. The distribution of LYVE-1 mRNA
The data are presented as copies of viral genomic RNA per ml of plasma.
This animal was determined after sacrifice to have been exposed but the infection was either unsuccessful or at such a low level as to be undetectable by 14 d PI.
Co-expression of DC-SIGN by nonvascular LYVE-1 mRNA+ cells in macaque colon
To begin to determine the phenotypes of the nonvascular population of LYVE-1+ cells and to determine whether these phenotypes changed during the course of SIV infection, we combined ISH for LYVE-1 mRNAs with IHC using an anti-DC-SIGN antibody (Figs. 2A and 2B). DC-SIGN/CD209 is a cell adhesion molecule that can capture HIV-1 and SIV, transport it to a long-lived intracellular compartment, and allow it to be presented in trans to T cells. 27 Although considered a DC marker, we and others have found it expressed predominantly in macrophage-rich areas of LNs.25,28 Of the LYVE-1 mRNA+ cells in the submucosal layer of the colon, 49.4% were also DC-SIGN+ and this percentage did not change appreciably during the course of infection (Fig. 2C). In contrast, LYVE-1 mRNA+ cells in the epithelium/lamina propria and muscle layers did not co-express DC-SIGN (100% and 99.4%, respectively) and this did not change during the course of SIV infection. In the central lacteals of the jejunum the abundant LYVE-1+ cells were all negative for co-expression of DC-SIGN protein (not shown). These data indicate that there are at least two distinct LYVE-1 mRNA+ populations in the submucosa of the colon, with individual, nonvascular LYVE-1+ cells co-expressing DC-SIGN, and vasculature-associated LYVE-1+ cells not co-expressing DC-SIGN.
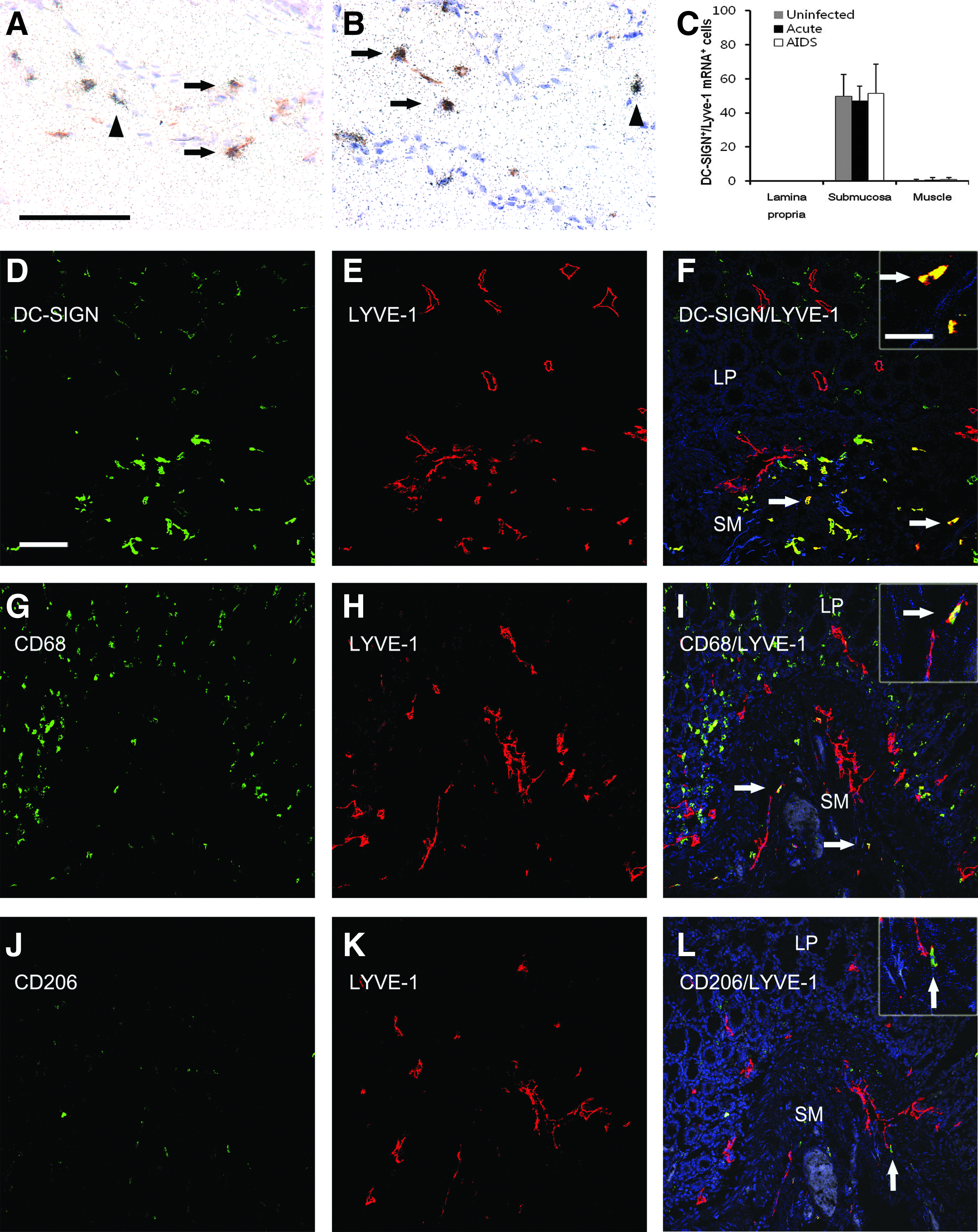
Co-expression of monocyte/macrophage/DC markers by LYVE-1 cells in macaque colon.
Co-expression of other macrophage and DC markers by cells expressing LYVE-1 protein
To explore further the different populations of cells expressing LYVE-1, we co-immunostained for LYVE-1 protein and other immune cell markers and performed confocal microscopy. In addition to DC-SIGN, we stained for CD68 or CD206. CD68 is expressed by macrophages and subpopulations of DCs and CD206 is the macrophage mannose receptor (MMR) expressed by macrophages and DCs. 29 CD206 is also a marker for macrophages with the M2 phenotype. 30 In the lamina propria of the colon, co-immunostaining for LYVE-1 and either DC-SIGN, CD68, or CD206 revealed that the LYVE-1+ lymphatic vasculature did not co-stain for DC-SIGN, CD68, or CD206, regardless of the stage of SIV infection. In contrast, in the submucosal and muscle layers of the colon, subpopulations of individual LYVE-1+ cells co-expressed DC-SIGN, CD68, or CD206 (Figs. 2 and 3). With regard to the extent of co-expression of these markers, nonvascular LYVE-1+ cells co-expressed DC-SIGN to a much greater extent than CD68 or CD206 (Figs. 2 and 3). The co-localization of the DC-SIGN fluorescence signals was particularly strong, compared to the CD68 and CD206 signals, and was almost exclusively in individual, nonvascular LYVE-1+ cells. Altogether these findings indicate that in macaque colon, in addition to afferent lymphatic vessels, macrophages and/or DCs also express LYVE-1.
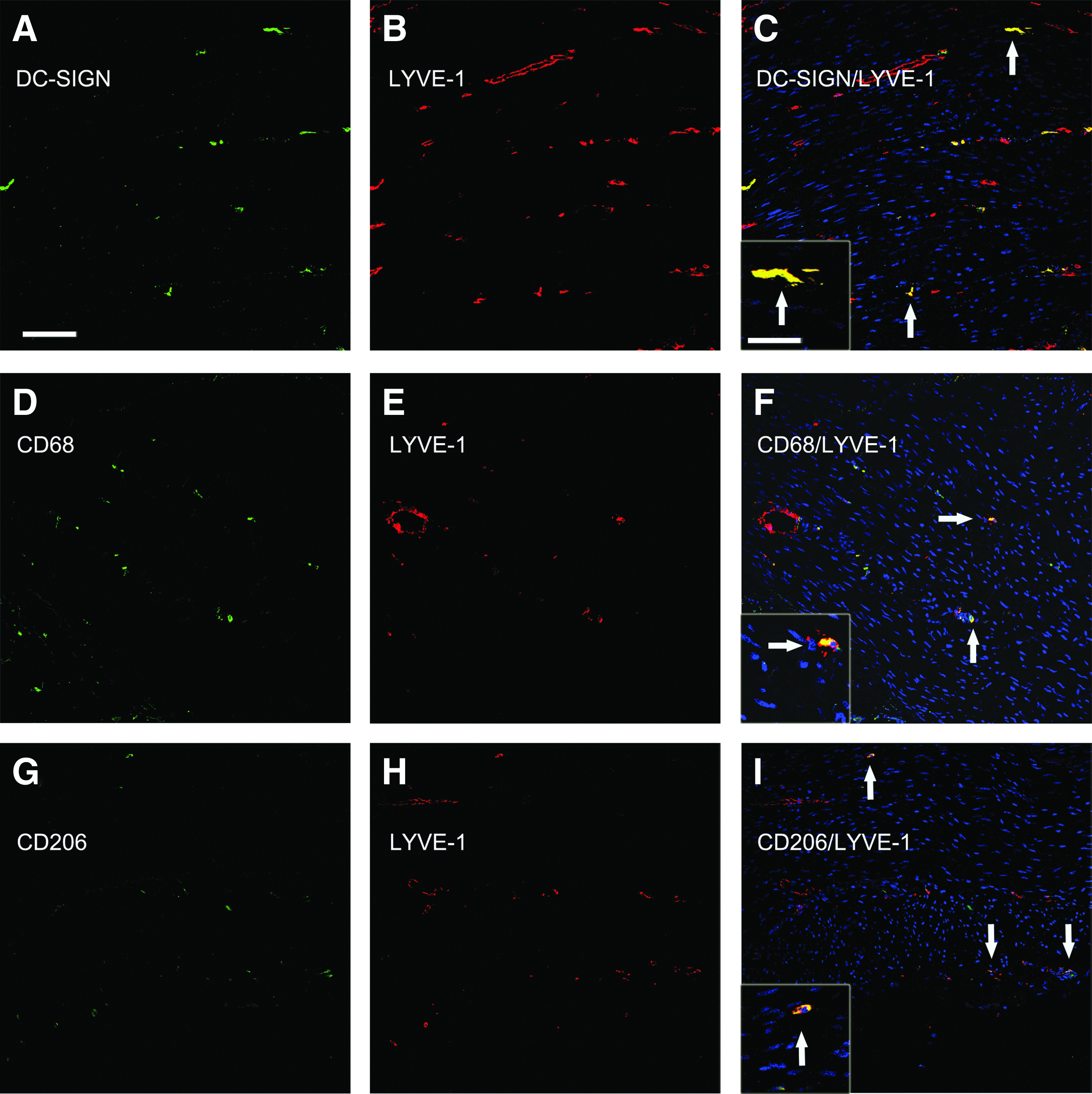
Simultaneous detection of LYVE-1 and monocyte/macrophage/DC markers in the muscle layer of macaque colon. Co-immunostaining for LYVE-1 and either DC-SIGN, CD68, or CD206 was performed as described in the legend to Figure 2. Shown are representative images from the muscle layer of the colon. Tissue sections were obtained from the following animals and disease states:
To determine whether in other tissues intimately involved in the immunology and pathogenesis of HIV-1/SIV infection, LYVE-1+ cells co-expressed DC-SIGN, CD68, or CD206, we examined LN and spleen tissues, again using confocal microscopy. LYVE-1 signal was mainly observed in efferent lymphatic endothelial cells located in sinusoidal latices in LN medullary areas as our previous study, 15 and in sinusoidal-like networks surrounding the lymphocyte-rich white pulp in spleen (Figs. 4 and 5). A large proportion of the LN sinusoidal LYVE-1+ cells co-expressed DC-SIGN, whereas a smaller proportion co-expressed CD206 (Figs. 4C and 4I), and very few co-expressed CD68 (Fig. 4F). Interestingly, many of the DC-SIGN+ and CD68+ cells were located immediately juxtaposed to LYVE-1+ cells, frequently appearing to localize within the lumens of the LYVE-1+ defined sinuses (Figs. 4C and 4F). In contrast, in spleen, cells expressing LYVE-1 did not appreciably co-express DC-SIGN, CD68, or CD206 (Fig. 5).

Simultaneous detection of LYVE-1 and monocyte/macrophage/DC markers in the medullary regions of macaque lymph nodes. Co-immunostaining for LYVE-1 and either DC-SIGN, CD68, or CD206 was performed on macaque LN tissue sections as described in the legend to Figure 2 and representative images are shown. Tissue sections were obtained from the following animals and disease states:
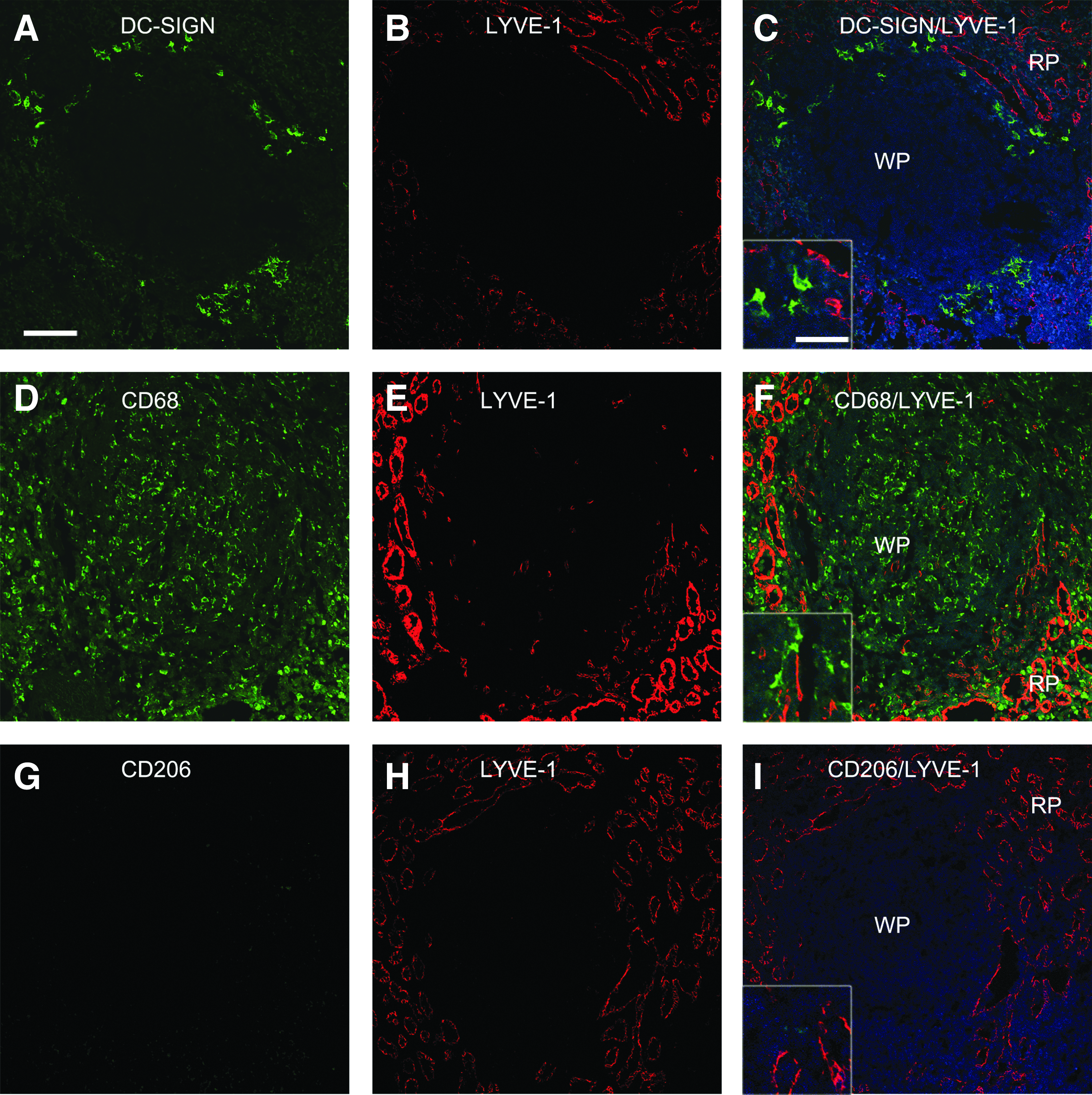
Simultaneous detection of LYVE-1 and monocyte/macrophage/DC markers in macaque spleen. Co-immunostaining for LYVE-1 and either DC-SIGN, CD68, or CD206 was performed on macaque spleen tissue sections as described in the legend to Figure 2 and representative images are shown. Tissue sections were obtained from the following animals and disease states:
Finally, to determine whether LYVE-1+ cells co-expressed other lymphatic markers, we co-stained tissue sections for podoplanin. Most cells expressing podoplanin were associated with vascular structures in the epithelium/lamina propria of the jejunum (Fig. 6A), and they co-expressed LYVE-1. Although much less abundant, individual cells expressing podoplanin were present apart from vascular structures and they did not co-stain for LYVE-1. In colon, podoplanin and DC-SIGN were not co-expressed by cells of the lymphatic vasculature or by individual nonvascular cells (Fig. 6B). Therefore, in macaque colon podoplanin was almost exclusively expressed by the lymphatic vasculature.
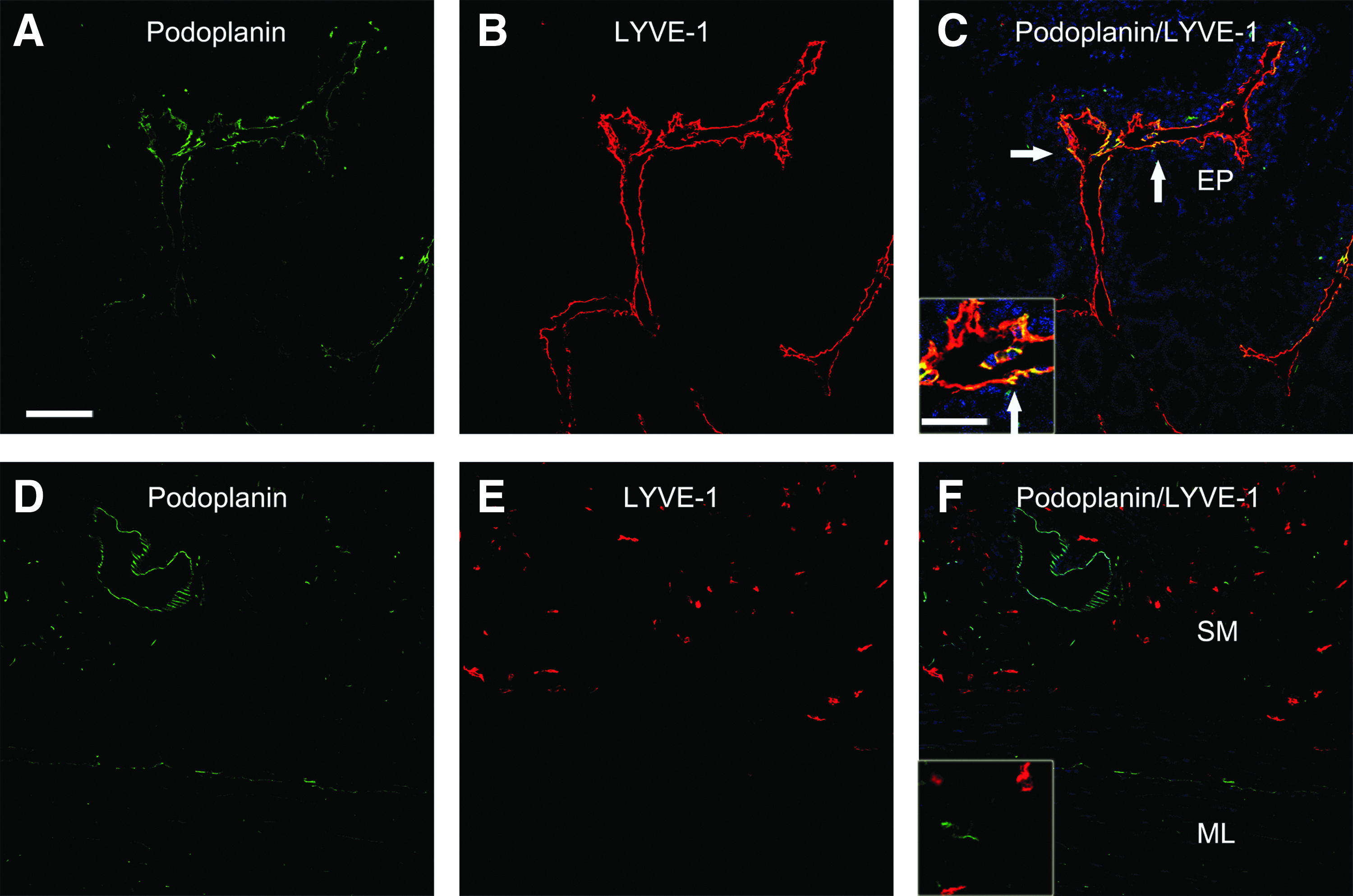
Simultaneous immunostaining for LYVE-1 and podoplanin in macaque intestine. Co-immunostaining for LYVE-1 and podoplanin was performed on macaque intestinal tissue sections as described in the legend to Figure 2 and representative images are shown. The epithelium/lamina propria (EP), submucosa (SM), and muscle layer (ML) are noted. Tissue sections were obtained from the following animals and disease states:
Discussion/Conclusions
In the present study we have investigated changes in the expression levels and patterns of LYVE-1 in cynomolgus macaque colon during SIV infection and have examined the phenotypic identities of vascular and nonvascular LYVE-1+ cells. LYVE-1 has been recognized as a reliable marker for LECs in the mouse,6,31 human, 6 and macaque. 15 Using ISH, we found the levels of expression of LYVE-1 mRNA in the macaque colon were decreased in macaques with AIDS compared with acutely infected or uninfected macaques (Fig. 1). These data indicate that the lymphatic vascular network and/or nonvascular LYVE-1+ cells in colon are affected by SIV infection. Possible explanations for the loss of LYVE-1+ cells as SIV infection progresses include changes in the recruitment, proliferation, life-span/turnover, egress, or differentiation of these cells in the local environment. Recently, LYVE-1+ sinusoidal endothelial cells were shown to be decreased in liver in patients with chronic hepatitis or cirrhosis, 32 possibly associated with local inflammation. In addition, the density of LYVE-1+ lymphatic vessels in human skeletal muscle decreases with age. 33 These reports and our results here indicate that LYVE-1+ cell populations are affected by persistent local and/or systemic inflammation and infection. Therefore, clinical interventions focused on reducing systemic inflammation and reducing microbial burdens would be expected to maintain or rebalance the LYVE-1+ cell populations in the colon and potentially elsewhere in the body.
Throughout the different layers of tissue in colon, we observed a large population of nonvascular LYVE-1+ cells that co-stained for DC-SIGN, and to a lesser extent CD68 and CD206. There are reports of LYVE-1 expression by cells other than LECs. LYVE-1 is expressed by Mac-1+ macrophages in mouse tongue, 34 by macrophages near inflamed pancreatic islets, 35 by bone marrow-derived macrophages 36 such as in the tip portion of epididymal adipose tissue in adult mice, 37 and by CD45+ conjuctival cells. 38 In addition, LYVE-1 is expressed by a population of CD68+ and DC-SIGN+ cells in placenta. 39 Whether there are lineage, trans-differentiation, and/or precursor/product relationships amongst these different LYVE-1+ populations is not clear and will require challenging marking and time course studies as recently performed in mice to study intestinal macrophage lineages. 40 Therefore, as with DCs and macrophages, multiple markers are needed to definitively identify LECs amongst a mixture of cells types and caution should be used when LYVE-1-specific antibodies are used to purify LECs from disaggregated tissues.
Our finding that nonvascular LYVE-1+ cells co-express DC-SIGN in the submucosa of macaque colon and that the LYVE-1+ vasculature does not, indicates that the LECs comprising the lymphatic vasculature draining the colonic mucosa do not express an important adhesion molecule and pattern recognition receptor used by a wide variety of microbes to enter migratory antigen presenting cells (APCs) such as macrophages and DCs. These microbes include HIV-1, SIV, HCV, Ebola virus, and Mycobacterium tuberculosis.27,41 Therefore, if the lymphatic vasculature in the colon is a target for infection by these microbes, it will not be DC-SIGN-dependent. In contrast, our findings reveal that individual, nonvascular LYVE-1+ cells strongly co-express DC-SIGN and are likely migratory APCs, although it is not clear what function LYVE-1 is performing in these cells. These cells could contribute to dissemination of microbes that are able to breach the mucosal barrier of the large intestine and vaccination or therapeutic approaches that target these cells could reduce susceptibility to infection by mutliple microbes. In contrast to the colonic mucosa, the medullary network in LNs, including the efferent lymphatic vasculature, co-expresses DC-SIGN. It is possible that DC-SIGN, as well as LYVE-1, are performing different functions in different cellular (APC versus vascular) and microanatomic (mucosal versus secondary lymphoid tissue) contexts. For DC-SIGN these might include antigen or microbe uptake, broader environmental sensing and signaling, or cell adhesion. For LYVE-1, which binds hyaluronan and aids in its turnover, these functions are not clear and will depend upon whether LYVE-1 is sialylated. 12 Nonetheless, the loss of the LYVE-1+ nonvascular APCs in the colon during the progression of SIV infection could potentially contribute to loss of immune function due to loss in local innate immune function, mucosal immune surveillance, and antigen presentation in draining LNs. Current antiretroviral therapies that potently suppress HIV-1 levels, and additional anti-inflammatory therapies, could help to reverse this loss of LYVE-1+ cells in the colon and possibly aid in restoration of local immune function.
Footnotes
Acknowledgments
The authors thank Mr. Jason Devlin and the Center for Biologic Imaging at the University of Pittsburgh for assistance with and use of the confocal microscope. We also thank Dr. Michael Murphey-Corb and laboratory for the care of the groups of animals that gave rise to the archival tissues examined here.
Disclosure Statement
This work was supported by the National Research Foundation of Korea Grant funded by the Korean Government (NRF-2010-013-E00026), NIH R01 AI90825, and the Center for Lymphatic Immunobiology at the University of Pittsburgh. Drs. Choi and Reinhart and Ms. Junecko and Ms. Klamar have no conflicts of interest or financial ties to disclose.