
Abstract
Abstract
Introduction:
Lymph nodes are often the target of radiotherapy procedures. Unfortunately, the impact of nodal irradiation on lymphatic function is uncertain. In this study, our aim was to quantify the impact of lymph node irradiation on lymph flow.
Methods and Results:
The popliteal node or the nodal excision site of rabbits was treated with four daily 8 Gy doses of radiation. A FITC-dextran tracer was infused into a prenodal popliteal lymphatic. The area under the tracer blood recovery curve (AUC) indicated lymphatic functionality and the inflow pressure versus flow rate relationship inferred resistance through the system. Fluoroscopic and histological examination provided supporting data. Radiation of intact nodes decreased lymph transport significantly at 1 week, 1 month, and 6 months post-treatment (AUCs of 207.9±79.87, 191.6±62.95, and 250.44±46.45) in comparison to controls (667.32±104.18). Surprisingly, this functional decline was similar to that detected with a combination of node removal and irradiation of the excision site. The pressure–flow relationships in all treatment groups were significantly different from controls. This may be due in part to fibrosis and the thickening of the nodal capsules and trabeculae observed at 1 and 6 months. Fluoroscopy and Evans blue dye studies revealed vigorous new lymphatic vessel growth and occasionally, vessels anastomosed with local veins.
Conclusions:
Irradiation of the popliteal lymph node impaired lymph transport and increased the pressure required to maintain flow through the system. New vessel formation and the growth of lymph-venous anastomoses indicated the development of alternative drainage pathways as a compensatory response.
Introduction
L
While the clinical data suggests that there is a significant relationship between radiation therapy and lymphedema,6–8 it is difficult to determine the impact of irradiation because it is almost always used in conjunction with other treatments. To compound the problem, radiation has not commonly been incorporated into animal models of lymphedema and consequently our understanding of its impact on lymphatic function is limited. It is generally acknowledged that radiotherapy applied to the nodes decreases cancer recurrence.5,9–11 However, apart from its immunological duties, the lymph node has a complex and generally underappreciated physiological relationship with its attendant pre- and post-nodal lymphatic vessels. How targeted radiotherapy impacts lymph transport through the nodes or affects the absorption of interstitial fluid from the upstream drainage basin is not entirely clear. In this study, our objective was to irradiate a single popliteal lymph node and quantify and compare several lymphatic functional parameters in a rabbit model that permits direct access to the lymphatics of interest. We compared four groups of animals; non-treated, single lymph node irradiation, node resection alone, and node resection with irradiation. These studies revealed that any decrease in lymph transport could be attributed largely to the effect of radiation itself.
Methods
A total of 170 male and female New Zealand white rabbits (2.5–4.5 kg) were used in this study. Animals were given free access to food and water for an observation period of 1 week preceding surgery. All experiments outlined in this article were approved by the ethics committee at Sunnybrook Health Research Institute and conformed to the guidelines set by the Canadian Council on Animal Care and the Animals for Research Act of Ontario.
Radiation therapy
Animals were anesthetized with an intramuscular injection of ketamine (50 mg/mL) and xylazine (5 mg/mL) and radiotherapy was applied to the popliteal fossa region in the hind limb using a Faxitron X-ray device (model #43855F, Faxitron Bioptics, LLC, Tusan, USA). Breast cancer patients typically receive a dose of approximately 50 Gy in 25 fractions of 2 Gy over a period of 6 weeks. Using the linear-quadratic model, we can provide a biological equivalent dose in rabbits by applying fractionated 8 Gy doses (160 kv, 6.3 mA, 33 cm from source, 4 cm depth) on 4 consecutive days. Biological equivalent dose is an established measure for quantifying the expected biological effect of different radiation dose fractionation schedules.12,13 In patients, radiotherapy is applied 4–6 weeks following surgery to permit tissue healing. We found that 3 weeks was sufficient healing time in rabbits. Outcomes were measured at 1 week, 1 month, and 6 months post radiation.
Surgery for popliteal lymph node removal
Rabbits were fasted 12 hours prior to anesthesia. The animals were anesthetized initially with an intramuscular injection of ketamine (50 mg/mL) and xylazine (5 mg/mL). Subsequently, 1.5%–3.0% isofluorane was delivered through an endotracheal tube. The surgical site was shaved and prepped with alcohol and betadine. The popliteal node lies in the hind limb posterior to the biceps femoris and can be located with palpitation. A vertical skin incision (approximately 2–3 cm long) was made over the lateral aspect of the popliteal region. The pre- and post-nodal lymphatic vessels were tied off with a 2-0 polysorb ties and the node excised. Hemostasis was ensured prior to closure of the surgical site. Subcutaneous buprenorphine (0.035 mg/kg) was given for pain management postoperatively and every 12 h for 2 days. Duplocillin (0.1 mL/kg; procaine penicillin 150,000 IU/mL, benzathine penicillin 150,000 IU/mL) was given intramuscularly on the day of surgery.
Assessment of lymphatic function; integrity of lymphatic network
The main role of the lymphatic vessel is to absorb extravasated vascular derived protein from the interstitial spaces and return it to the venous circulation. Therefore, the ability to transport a macromolecule to plasma provides a quantitative measure of the lymph transport effectiveness of a given lymphatic network.14–16 A schematic illustrating the features of the experimental design is provided in Figure 1A.

Schematic illustrating essential features of experimental design. Evan's blue dye was injected subcutaneous into the foot to highlight lymphatics. A 25-gauge cannula was inserted into an afferent lymphatic in the direction of flow.
To measure lymph transport and the integrity of the lymphatic network, Evans blue dye (0.5% in saline) was injected into multiple sites on the dorsal surface of the hock to permit visualization of the popliteal prenodal ducts. One of these vessels was dissected free of connective tissue and cannulated in the direction of flow using a 26-gauge angiocatheter. A fluorescent lymph flow tracer (FITC-dextran 70 kDa, 50 mg/mL, Sigma-Aldrich, Oakville, Canada) was infused via a syringe pump (2 mL/h, 400 μL, model #260, Kd Scientific Inc, Holliston, MA) and the recovery of the tracer monitored over time in plasma. Blood samples were obtained via a 22-gauge angiocatheter placed in the central auricular artery. Prior to infusion, a blood sample was taken for baseline and then once infusion had begun, a sample was taken every minute to 10 min, and then at 15, 20, 25, 30, 60, 90, and 120 min. Accumulation of the dextran tracer in plasma was determined using a plate reader at 520 nm wavelength. A concentration (as a percent injected) versus time plot was then generated and the area under the each curve (AUC) was calculated using the trapezoidal rule. As we have demonstrated in our previous work, this dimensionless number is an effective way to quantify lymphatic functionality over time.14,15
Assessment of lymphatic function; Resistance through lymphatic network
Hydraulic resistance is an alternate way to assess the functionality through the popliteal lymphatic system. By measuring the inflow pressures during the infusions into the prenodal ducts, quantification of the systems resistance can be deduced. To achieve this, a prenodal lymphatic was cannulated (in the direction of flow) with a 26-gauge angiocatheter connected to a stopcock. One arm of the stopcock was attached to a syringe pump and the second arm was attached to a pressure transducer (Custom CDX3, Cobe, Richmond Hill, Canada; or 042982100AT, Argon Medical Devices, Texas, USA) (Fig. 1B). Inflow pressures were recorded at a rate of 10/sec on a data acquisition system (Daqview Software, A-tech Instruments, Toronto, Canada). The infusate consisted of ‘artificial lymph
Assessment of limb water
Tissue water content of effected limbs was monitored using a commercial bioelectrical impedance device (ImpediMed, SFB7, Carlsbad, CA). This device passes a small current through the tissue and the impedance or the resistance to the flow of this current is directly relatable to the amount of extracellular fluid in the limb. The greater the water content, the lower the resistance. To take each measurement, 25-gauge needles were placed subcutaneously at the four locations outlined in Figure 2. Each animal had a baseline reading before experimental procedures, as well as one additional reading in the same limb before sacrifice. The difference between baseline and the final reading was expressed as a percent change. A positive percent change equated to greater tissue water. A negative percent change indicated a loss of water in the limb in comparison to baseline readings. Bioimpedance was also monitored in the contralateral non-treated limbs.

Bioelectrical impedance spectroscopy. 25G needles were bent (1.5 cm x 1.5 cm) and placed subcutaneously. Leads were then attached in the pattern illustrated in the figure. The red lead (1) was placed at the region where toes meet foot on dorsum. The yellow lead (2) was placed 2 cm cranially from the red. Blue (3) was placed on the contralateral patella, and black (4) was placed on the external occipital protuberance of the head. Bioimpedance was measured in the area between the yellow and blue leads (red hatching).
Visualizing lymphatics
To visualize the lymphatics in all experiments, Evan's blue dye was injected subcutaneously into the dorsum of the hind foot. The dye binds to interstitial proteins and is subsequently taken up by initial lymphatics. Additionally, an X-ray contrast agent (1–3 mL of Lipiodol, EZ-EM Canada, Anjou, Canada or 1–3 mL of Visipaque (iodixanol) 320 mg/mL, GE HealthCare, Mississauga, Canada) was injected into a prenodal lymphatic vessel using a 26-gauge cannula and a mobile fluoroscopy system (BV Pulsera, Philips, Markham, Canada) was used to visualize the lymphatic vessels and nodes of the hind limb.
Histology
Following sacrifice, nodes and surrounding fatty tissues were placed in 4% paraformaldehyde. Tissue was then sectioned and stained using hematoxylin and eosin. To visualize fibrosis, Masson's Trichrome stain was used.
Lymph–venous anastomoses
The visualization of limbs that received Evans blue dye revealed an unexpected finding. In some preparations, lymph–venous connections were observed upstream of the irradiated/excised lymph nodes. For further confirmation of these anastomoses, in a limited number of animals, FITC-dextran tracer was infused into a popliteal pre-nodal lymphatic as described earlier and blood samples were taken from both femoral veins simultaneously at 0, 2.5, 5, 10, 15, 20, 25, 30, 60, 90, and 120 min. Limbs containing a lymph–venous connection should display enriched tracer concentrations in the ipsilateral side.
Experimental groups and data analysis
Four groups were compared in this study. (1) Nontreated animals provided baseline measurements for all experiments. (2) Irradiation of a single popliteal lymph node. Outcome measures were performed at 1 week, 1 month, and 6 months post-surgery. (3) Node resection alone. Outcome measures were performed 1 and 6 months post-surgery. (4) Node resection with irradiation. Outcome measures were performed 1 and 6 months post-surgery.
Transport and resistance data were analyzed with one-way or repeated measures ANOVA with Dunnets post-hoc test as appropriate. Bioimpedance data were analyzed with paired t-test. Data are presented at mean±SE. We interpreted p<0.05 as significant.
Results
Radiation effects: General observations
Radiation was well tolerated by all animals. Slight to moderate erythema developed immediately following the completion of treatment but generally subsided by 3 months post-irradiation. Desquamation also occurred in some cases. The extent of radiation-induced scarring varied greatly from animal to animal. In some cases, no visible scarring was noted, while in others extensive scarring was noted at both the 1 and 6-month time points. In two animals, tissue hardening developed directly upstream of the irradiated area.
Lymphatic function
Lymphatic function data is illustrated in Figure 3. In intact limbs, the plasma accumulation of the tracer increased over time, peaking at approximately 20 min after the start of tracer infusion. This group generated a mean AUC of 667.32±104.18 (Fig. 3). There was a significant decrease in functionality observed in limbs subjected to radiation treatment alone (Fig. 3A). AUCs for 1 week and 1 month were 207.9±79.87 and 191.9±62.95, respectively (p=0.018 for both). Lymphatic functionality was only slightly increased at the 6-month time point (250.44±46.45, p=0.034), indicating a relatively long-term deficit in lymph transport in this group. It should be noted there were a significant number of animals in each of the early radiation groups that lacked suitable lymphatics to cannulate (15 animals). In these cases, the vessels present were simply too small to carry out the procedure.

Transport of FITC-Dextran. The transport results are displayed as area under the curve of the transport vs. time graph. Data were analyzed with a one-way ANOVA with Dunnets post hoc. Significance from control (p<0.05) is denoted by star.
On average, node removal caused only a modest decline in function at 1 month (590.28±139.88, p=0.741). However, this appeared to be an anomaly as two animals had very high AUCs (938 and 839), both of which were greater than the average for intact (control limbs). If we omit these two animals, the average AUC would be 341.97±142.38. Lymph transport at 6 months was less than controls but did not reach significance (AUC 274.03±142.58, p=0.183) (Fig. 3B).
As expected, limbs that underwent both node removal and radiation also saw a significant decrease in accumulation at 1 month (AUC of 227.18±105.92, p=0.019) (Fig. 3C). This deficit was somewhat resolved over the next 6 months (AUC 311.79±146.96, p=0.092) to the extent that these data were not significantly different from controls. In this regard, there were four animals subjected to node removal and radiation (two at 1 month and two at 6 month time points) who had functionality levels that were similar to the intact limbs. As will be discussed below, we observed some unusual patterns of new lymphatic vessel growth upstream of the site of injury in these animals and lymph-to-venous anastomosis may have occurred.
Resistance to flow
These studies presented a considerable experimental challenge. In the radiation treatment groups, the pre-nodal lymphatics were often much smaller than controls and we had difficulty cannulating a suitable duct. This was especially an issue in the node excision and combination treatment groups, in which 3/12 and 6/15, respectively, were unsuccessful. Additionally, we had multiple experiments in which the treated lymphatic networks began to leak midway through the experiment and we were unable to obtain any useful pressure recordings. This was a problem in 2/15 in nodectomy limbs and 3/12 in the combination groups. Ultimately, we were able to obtain data for 24 limbs (Fig. 4). As the 6-month data for the three treatment groups did not differ significantly from 1-month data, the results for the 2 times were combined to facilitate statistical analysis.
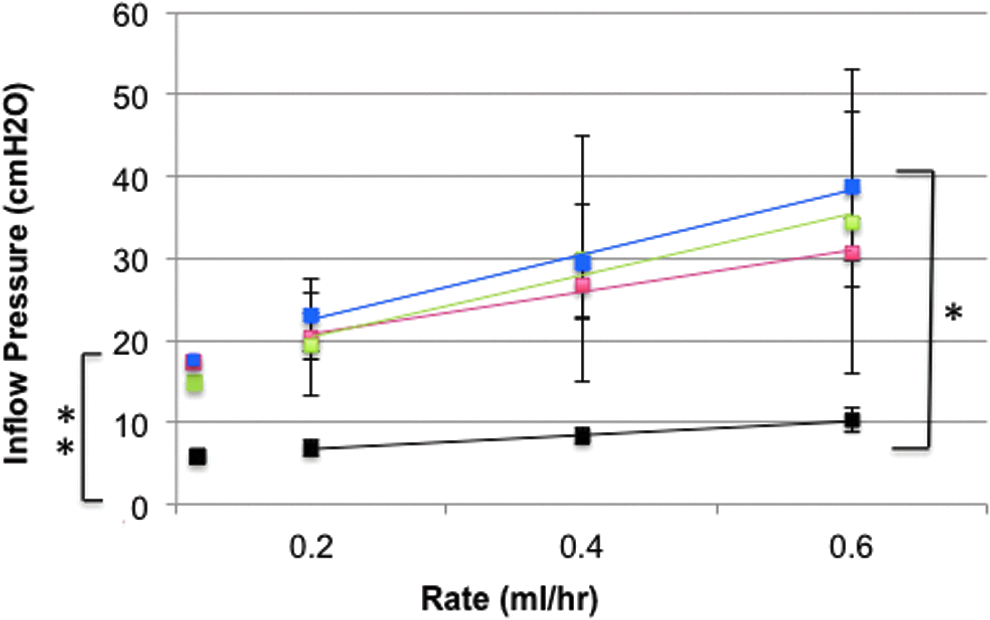
Resistance to flow. The pressure–flow relationships in all treatment groups were significantly different from controls (radiation, p=0.049; nodectomy, p=0.045, and combination treatment, p=0.005). Additionally, the average slope of the pressure–flow relationship was greater for the irradiated limbs (3.84±0.53 vs. 1.70±0.24 in nontreated animals). The y-intercepts (16.55±2.03 for radiation treated, 16.34±5.02 for nodectomy group, and 17.18±2.02 for combination treatment group) were significantly higher than those of controls (5.04±0.85). Control limbs (black, n=6). Radiation (red, 1-month, n=5, and 6-months, n=3, were combined). Nodectomy (green, 1-month, n=4). Combination Treatment (blue, 1-month, n=5, and 6-months, n=1 were combined). Pressure–flow relationships were analyzed with repeated measures ANOVA and Dunnets post hoc. Significance from control is denoted by a star. Slope and Y-intercept data was analyzed with one-way ANOVA with Dunnet's host hoc. Significance from the control (p<0.05) is denoted by multiple stars (y-intercept).
The pressure–flow relationships in all treatment groups were significantly different from controls (radiation, p=0.049; nodectomy, p=0.045; and combination treatment, p=0.005) (Fig. 4). The slopes for all treated groups were greater then that of the control (1.70±0.24), although only the combination treatment group reached significance (combination treatment 4.70±2.83, p=0.027; radiated 3.84±0.53, p=0.0.84; nodectomy 3.27±1.54, p=0.266).
The most striking observations in the resistance studies were that 1) the pressure–flow relationships were shifted upwards in all of the treated groups, and 2) that the three treatment groups were quite similar to one another. In the first instance, the y-intercepts for radiation alone (16.55±2.03 cm H2O, p=0.002), nodectomy alone (17.18±5.02 cm H2O, p=0.009), and radiation plus nodectomy (17.18±2.02 cm H2O, p=0.002) were all significantly higher than controls (5.04±0.85 cm H2O). The y-intercepts can be used to denote a theoretical ‘opening pressure’ for the lymphatic system. Therefore, it would appear that a greater lymphatic pressure is required to establish flow in limbs subjected to radiation therapy. Second, as is indicated above, the y-intercepts for the treatment groups are all very similar. This suggests that the major insult to the lymphatic system may due largely to radiation injury and not to the removal of the lymph node.
Tissue water
In limbs subjected to irradiation, nodal excision or a combination treatment, visible edema was only rarely observed. This is generally supported by the bioimpedance data as typically, there was not more than a 250 ohm difference between pre- and post-treatment measurements regardless of treatment. To put this into perspective, baseline bioimpedance measurements in untreated limbs ranged from 1300 to 1900 ohms. Untreated limbs underwent slight changes (less than ±2%) over the course of the study. This is most likely due to slight variations in hydration level of the animals at the time of measurement.
Following node removal, there was a significant increase in tissue water found at 1 month (p=0.028, Fig. 5), however this edema resolved over the next 6 months. While no other treatment group demonstrated a significant change in water content (most likely due to the large variability within each group), there was an unexpected trend. Following radiation treatment, tissue water declined especially at 6-month time point (p=0.094). This suggested that these limbs had less water than the pre-treatment level. Also, when the nodal excision site was irradiated, the trend was to a decrease in tissue water in comparison with nodectomy alone.

Assessment of tissue water with bioelectrical impedance. Following nodal excision (1 month), a significant increase in tissue water was observed (indicated by star, p=0.028). This increase appeared to resolve over 6 months. No other treatment had a significant effect, although a trend to reduced tissue water was observed in all radiation treated limbs. Radiation 1 week (n=10), 1 month (n=4), 6 months (n=8). Nodectomy 1 month (n=6) and 6 months (n=8). Combined treatment 1 month (n=9) and 6 months (n=8). Paired t-test, p<0.05.
Visualization of the lymphatic system: Evans blue studies
Observations made during study using Evans's blue dye have been summarized in Figure 6. In intact nontreated controls, it is typical to see two large afferent lymphatic vessels (∼0.5 mm) that run parallel to the saphenous vein on both the medial and lateral aspects of the lower limb leading to the popliteal node (Fig. 6A). There are no connections between the medial and lateral vessels until they coalesce at the popliteal node.
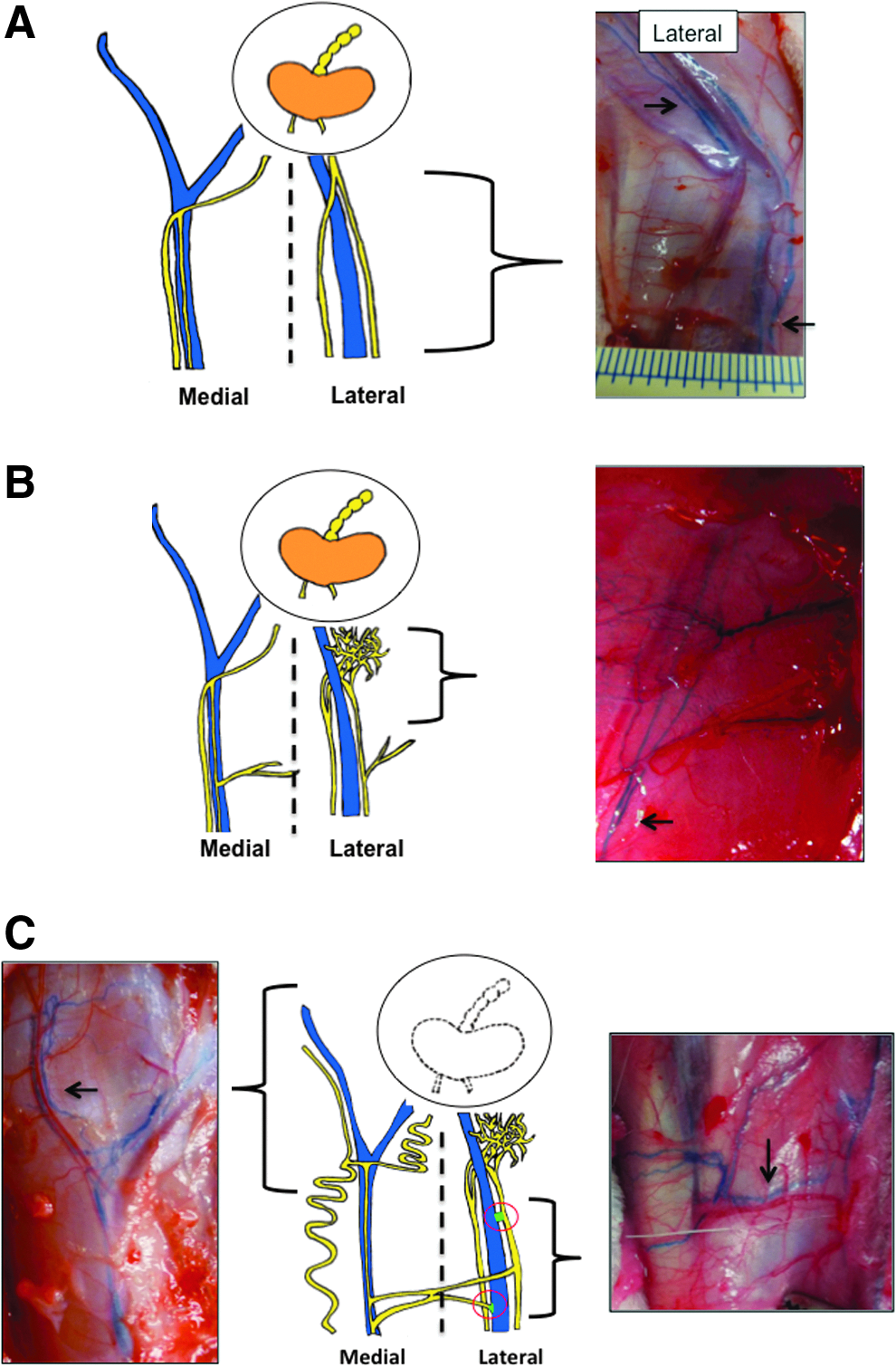
Schematic illustrating patterns of lymphatic regeneration following irradiation and lymph node excision with irradiation. Afferent lymphatic vessels are indicated by arrows.
In limbs subjected to radiation (Fig. 6B), lymphatic vessels tended to be very delicate and prone to leakage (at all times analyzed). Afferent vessels leading into the node had a lattice appearance (<0.5 mm, 1 month and 6 month time points). The lymphatics were typically shrunken in appearance in comparison to control limb vessels.
Following removal of the popliteal node, it was typical to see dilation of the afferent lymphatics in comparison to controls, no doubt due to the impediment to flow caused by the removal of the node (not illustrated). Some fluid continuity had been re-established by the 1-month time point and to support this, the regenerating lymphatic network showed a lattice appearance.
Lastly, a combination of both node removal and radiation appeared to induce the most radical compensatory response (Fig. 6C). A large increase in number and/or branching of lymphatic vessels was observed. In some cases, underlying vessels were found to branch into the dermis. Instead of branching into the popliteal region, lymphatics tended to do one of two things. In some animals, collateral pathways appeared to form around the popliteal region. In others, vessels upstream of the popliteal region (both on the lateral and media aspects of the limb) would frequently branch at right angles towards the saphenous vein on the lateral side of the limb. In some cases, medial and lateral vessels would coalesce and in others it appeared that there were lymph-to-venous connections formed between an afferent vessel and a vein.
Visualization of the lymphatic system: Fluoroscopy
Intact nontreated preparations revealed the cannulated afferent vessel emptying into the popliteal node and a single large diameter efferent vessel draining it (Fig. 7A). Limbs subjected to radiation were prone to leakage from both the afferent vessel as well as from the node. At the 1-week time point, leakages were noted, and by 1 month, channels were visible within the node (Fig. 7B). Six months following irradiation, additional lymphatic vessels branching around the periphery of the popliteal node could be seen (Fig. 7C). At 1 month following node removal, continuity was been re-established and leakage into the node excision site was often seen (Fig. 7D). In rare cases collateral pathways had developed at 6 months following nodectomy (Fig. 7E). Combined treatment limbs also displayed fluid continuity by 1 month post-treatment; however abundant branching and the formation of multiple collateral pathways were often present (Fig. 7F).
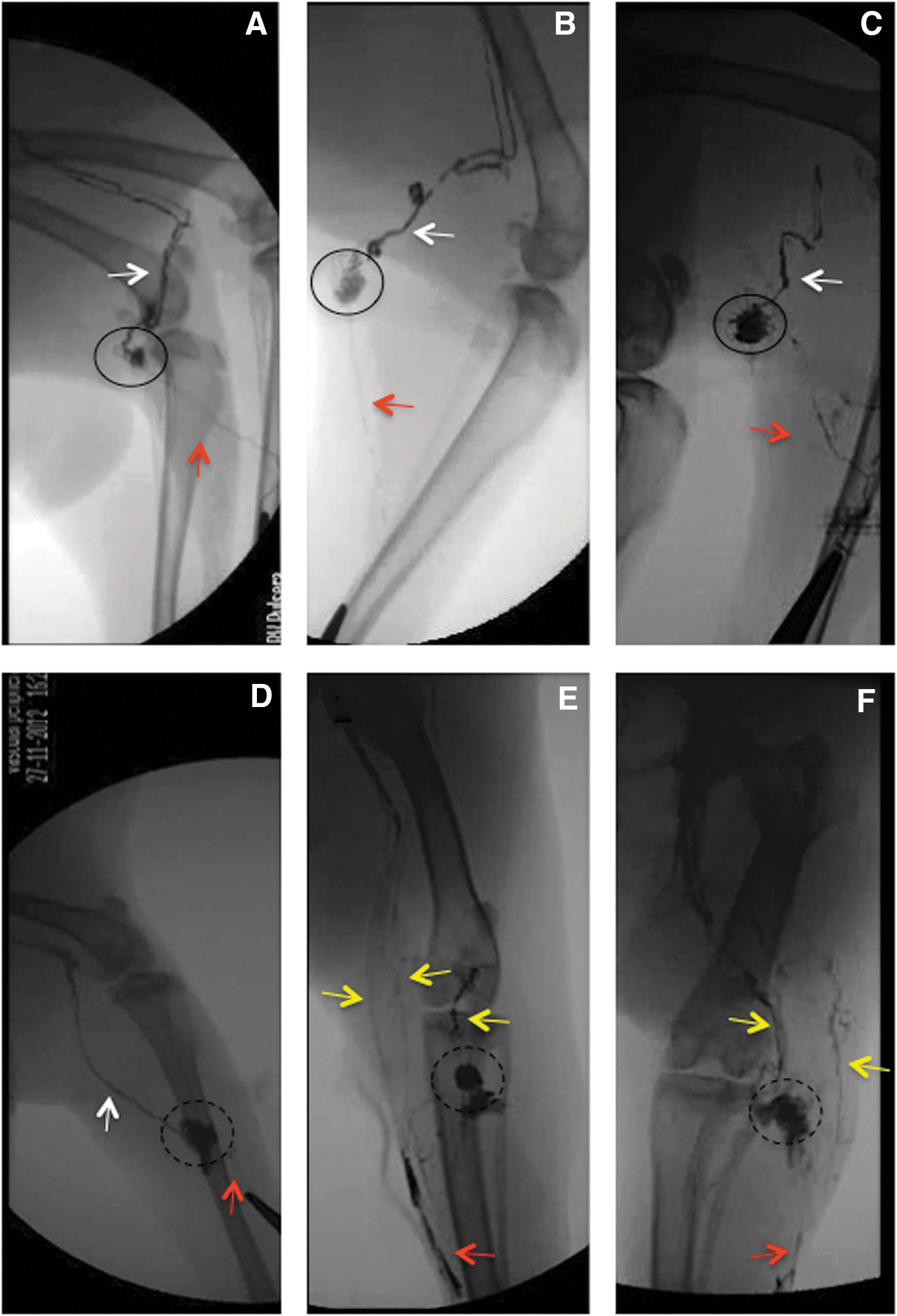
Fluoroscopy.
Histology
In the node intact animals, the most obvious change in the lymph nodes following radiation was an increase in collagen distribution at the 1 and 6-month time points (Fig. 8). Most evident was a thickening of the node capsule and trabeculae.

Histology of lymph node following irradiation. Popliteal nodes were sectioned and stained with Masson's Trichrome. Blue color indicates presence of collagen, which increases following radiation exposure. Thickening of capsule and trabeculae is especially evident (arrows). (A) Control;
Lymph–venous anastomoses
The development of potential lymph–venous anastomoses was unanticipated and we did not turn our attention to this phenomenon until midway through the study. Our first indication that a lymph–venous connection could be occurring was the observation of Evan's blue dye in veins running parallel to afferent lymphatics of treated limbs (Fig. 9A). When Evan's blue dye is injected subcutaneously, it binds to interstitial protein and is then absorbed into the lymphatic system. Normally, no dye enters the local vasculature but we identified 10 limbs in which the dye was observed in a lower limb vein connecting with a nearby lymphatic vessel. Eight potential anastomoses were identified in the combination treatment group and 2 in the node excision animals.
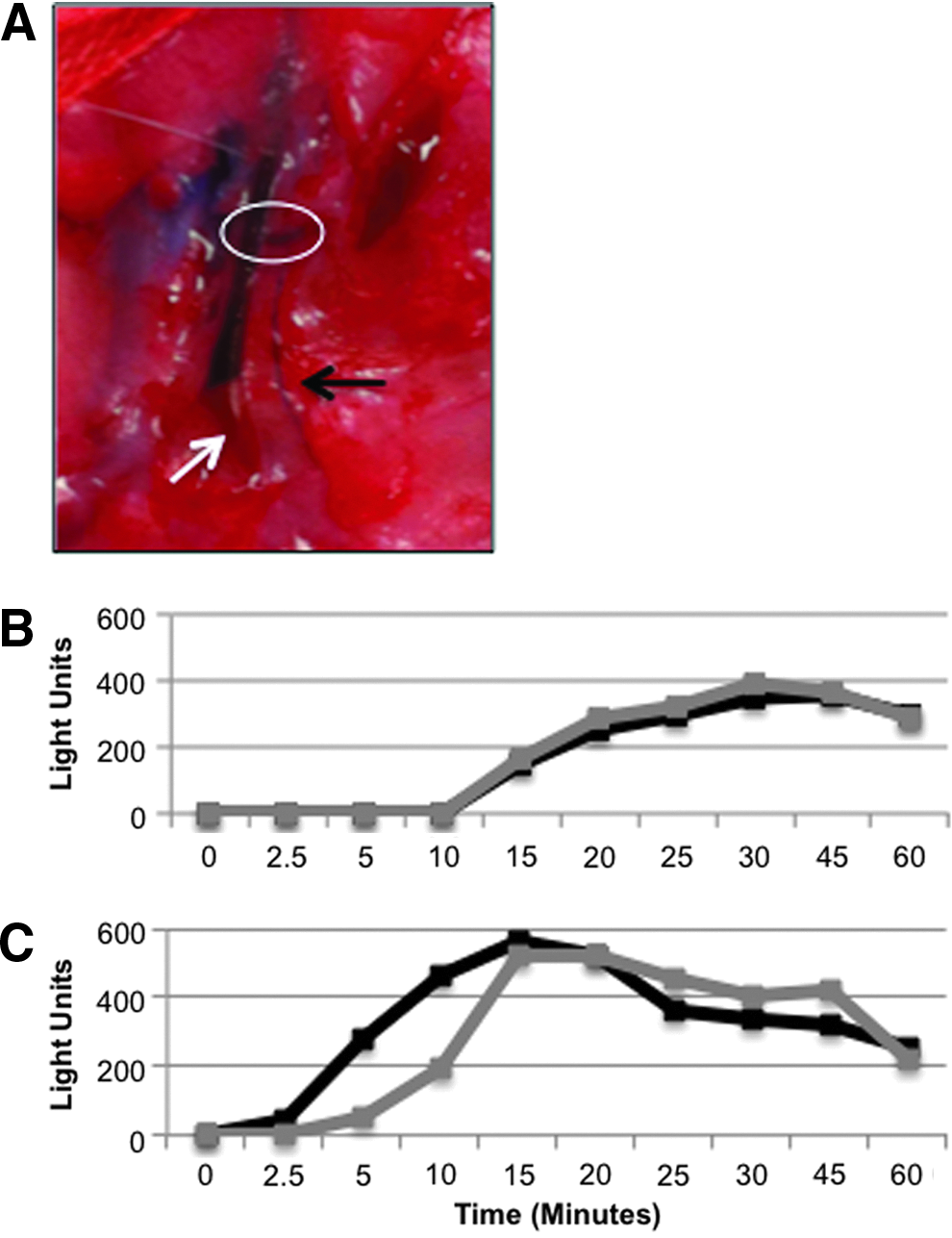
Evidence of lymphatic–venous anastomosis.
We attempted to obtain physiological evidence to support the presence of anastomoses but conditions were not favorable for routine analysis. An upstream afferent lymphatic that was large enough for cannulation in combination with the ability to sample blood from both ipsilateral and contralateral femoral veins was only achievable in two treated animals. While it is difficult to draw any conclusion, the results are intriguing nonetheless. In one of the two of these animals (combination treatment), the tracer appeared in the ipsilateral blood (with suspected anastomosis) before appearance in the contralateral side (Fig. 9C). In addition, the tracer began to accumulate in the blood earlier (5 vs. 15 min) and also peaked earlier (15 vs. 30 min) than in control limbs. In contrast, in the second animal with a potential anastomosis, as well as in untreated limbs with no suspected anastomosis, we observed simultaneous accumulation of tracer into blood from both the infusion limb and the contralateral limb (Fig. 9B).
Discussion
Radiation of the lymph node induced a deficit in lymphatic function
Damage to the lymphatic system occurs frequently during the innumerable surgical procedures that occur in hospitals around the world. However, while acute edema is a natural response to tissue injury, chronic tissue swelling (lymphedema) is not a characteristic feature of most of these surgeries. However, the removal of one or more lymph nodes appears to be a significant causative factor in lymphedema development. For this reason, our studies have focused on defining the lymph transport deficit caused by nodal excision, and developing methods to restore function after node removal including autologous node transplantation and the introduction of lymphangiogenic factors into the nodal excision site.14–16 In this report, we have investigated the impact of single lymph node radiation on lymphatic functional parameters.
Irradiation of a single popliteal lymph node reduced the lymphatic-to-blood transport of the lymph flow tracer to about a third of that observed in non-irradiated controls. In addition, when inflow pressures were measured, the irradiated nodes caused a significant upward shift in the inflow pressure–flow relationship. This indicated that much higher pressures were needed to move lymph in the irradiated limbs. For example, at 0.4 mL hour, a pressure of 8.3±0.92 cm H2O was required to move lymph in nontreated controls, whereas 26.64±3.9 cm H2O pressure was necessary in the irradiated preparations. The latter result suggested a greater resistance to flow following radiation treatment. Perhaps surprisingly, the inflow pressure–flow relationships in the nodectomy alone or combined treatment groups were very similar to those in the radiation-treated limbs. We expected that the removal of a lymph node or the combination of nodal excision plus irradiation would lead to a greater functional deficit, but this did not appear to be the case. These data suggested that the radiation itself was a main factor in reducing lymph function in this model.
Choice of animal model
Radiation with or without surgical intervention has been applied to dogs, 20 rats, 21 and mice 22 with varying results. A notable difference between this study and previous radiation models is the extent/or location of radiation application. Radiation therapy is generally applied to tumor excision sites and the downstream drain nodal beds. In previous animal models, entire limbs were often subjected to radiation, while our experiments were limited to radiation of a single popliteal lymph node or nodal excision site. In these models, the major focus was on inducing lymphedema, usually requiring significant lasting damage to lymphatic system, while the goal of this study was to determine the effect of radiation therapy on lymphatic functionality.
A problem with the smaller species is that it is difficult to quantify the physiological parameters that are most relevant to lymphatic drainage. We have devoted considerable effort in developing larger animal models that permit direct access to lymphatic vessels, which can then be manipulated as appropriate. Much of our work has centered on studies in sheep.14–16 However, we do not have the ability to irradiate an animal of this size and consequently, have adapted our methods to a more suitable species, the rabbit. As can be seen in Figure 6, the popliteal system in this species is easy to identify with afferent (prenodal) ducts emptying into a single lymph node and one (post-nodal vessel) draining the node. For the most part, the large vessel size in this species allows us to use the methods we developed in sheep to assess lymphatic function (Fig. 3). Additionally, this species can be irradiated in devices designed for small animal studies.
Choice of radiation protocol
In the study reported here, we used a fractionation schedule that mimics to some extent, the protocols that are used clinically. In breast cancer, the standard radiation dosage is∼2 Gy delivered in twenty-five individual fractions. However, due to logistical and practical reasons, we reduced the number of fractions given. The use of high doses, such as 8 Gy, is limited mostly to boost treatments (radiation delivered to tumor beds at the end of radiation schedule in patients undergoing breast conservation surgeries) and palliative care. Utilizing this higher dose would most likely cause increased damage to healthy tissues, especially the skin and vascular systems. To what extent the lymphatic system would be affected by this radiation dose is unknown.
We realize that in cancer patients, some radiation-induced effects may take a long time (even years) to develop. However, we know very little of the chronology of pathophysiological events that occur in animal models exposed to ionizing radiation. Rather than commit to longer-term studies at this time, we thought it prudent to focus our initial efforts on a 6-month period, as it seems likely that radiation-induced injury to the lymphatic system, sets in motion a number of relatively early events that facilitate the development of chronic lymphatic failure. As an example, an overexpression of genes involved in collagen deposition (most notably TGF-β1), has been observed as early as 3 weeks following irradiation. 23 Once the most relevant injuries to the lymphatic system have been identified, selected parameters can be examined over a longer time frame if appropriate in future studies.
How does radiation affect lymph transport through a lymph node?
One normally does not consider a function for the lymph node in the fluid balance of a limb. However, in addition to its immunological duties, the lymph node has a complex physiological association with its attendant pre- and post-nodal lymphatic vessels. Lymph contains less protein than that in the nodal capillaries and on passage through the nodes, oncotic forces draw water from lymph into the blood. As such, lymph is concentrated and volume is lost on passage through the node.24–26 In doing so, lymph nodes anatomically separate the afferent and the efferent lymph vessels into higher and lower pressure systems, respectively. 27 This is important because lymphatic vessels contract and this pumping action provides a major portion of the force required to move lymph through the lymphatic circulation. Pre- and post-nodal lymphatics have different contractile properties and reach peak pumping performance over different pressure ranges. 28 Therefore, if the ability of the node to concentrate lymph were compromised by radiation, the downstream vessels may be forced to function over a nonoptimal range of pressures with the result that lymph drainage of the limb could be reduced. Indeed, based on studies using lymphatic congestion lymphoscintigraphy, it is evident that lymph pump failure occurs in breast cancer-related lymphedema. 29 Surprisingly, the lymph node itself is capable of contracting and radiation may negatively impact this function as well. 30
While irradiation of the node may affect several parameters related to the aforementioned physiological parameters (for example the permeability of the nodal vasculature), excess collagen deposition in the node is a factor to consider. Fibrosis is a well-documented consequence of ionizing radiation and in the study reported here, one of the most obvious changes we observed following radiation of the node was the deposition of collagen with a thickened node capsule and interior trabeculae. Presumably, transforming growth factor beta 1 (TGFβ1) 31 plays a role in this process by promoting myofibroblast proliferation and extracellular matrix deposition. TGFβ1 has also been shown to cause endothelial cells to undergo a phenotypic conversion into fibroblast-like cells (endothelial-mesenchymal transitional (EndMT) event, 32 which would exacerbate the ongoing fibrotic process in irradiated tissues. There is some evidence that the EndMT phenomenon occurs in lymphatic endothelium following radiation exposure,22,33 but whether such an event occurs within the node parenchyma is unknown.
New lymphatic vessel growth and relationship with local veins
The pre- and post-nodal popliteal lymphatic vessels are normally well delineated in the rabbit hind limb and follow a relatively linear course. With lymph node excision, there is vigorous new vessel growth in an attempt to restore fluid continuity. In past studies in sheep, vessel regeneration was capable of restoring lymphatic drainage to about 80% of control values over a 4-month period. 16 From the literature, we learn that the number of lymphatic vessels is altered by radiation, but results have been inconsistent. For example, in one study, the number of lymphatic vessels decreased one week following a large dose of radiation (15 or 30 Gy) in a mouse-tail model. 22 This decrease continued over the course of the study, reaching a 4-fold loss of vessels in comparison to controls by 6 months. Alternatively, in humans, patients receiving conventional delivery (small dose 5 days a week) of 30–40 Gy had an increase in the total number of lymphatics (mainly those of diameter <10 μm) at 1-year time point. 34
In the study reported here, both Evans blue and fluoroscopy demonstrated that the number/density of lymphatic vessels increased following irradiation, even though fibrosis has been shown to inhibit lymphangiogenesis. 33 The pattern of new vessel growth was however, extremely variable. Both densely and sparsely packed vessels were observed upstream of the irradiated popliteal node or its excision site. Heavily branched vessels were also commonly noted. The upstream existing lymphatics (up to 15 cm away) were often observed to branch laterally, apparently using the vasculature for guidance. This is expected as lymphatic sprouting and growth has been found to be dependent on vascular derived VEGFC or VEGFD. 35 While some function could be attributed to these vessels as indicated by the tracer transport to plasma in all treatment groups at all times, the irradiated lymphatics were delicate and much more prone to leakage in comparison to controls.
An unexpected finding was the presence of connections between afferent lymphatic vessels and veins. Under normal conditions, almost all lymph is returned to the bloodstream at the major lymph–venous junctions in the neck. However, lymph–venous anastomoses in unconventional locations have been reported in various species (rats, cat, dogs, monkeys) as well as in humans in response to lymph obstruction or edema.36–40 In a study conducted on 552 patients with primary and secondary lymphedema, as well as lymph node malignancies, 16 anastomoses were identified by lymphography. 41 Iodinated (125I) human serum albumin was injected intra-lymphatically to detect lymph–venous connections physiologically in 40 post-mastectomy patients. 42 Blood was taken from both basilic veins simultaneously and an increased quantity of the labeled protein transferred locally to the ipsilateral venous system in the non-edematous group. The presence of a lymph–venous connection was confirmed radiographically in two of the non-edematous patients. Little local transfer of the tracer was noted in the edematous patients or volunteers. Similar connections have been suggested in more recent studies involving subcutaneous injection of radiolabeled human IgG in patients with established breast cancer related lymphedema.43–45 These findings imply that lymphovenous connections may compensate for reduced lymph drainage in arms following breast cancer treatment.
Considering that the formation of these junctions is a relatively rare event, we were surprised to observe lymphatics filled with Evans blue dye apparently joining with local veins that also contained some of the dye. Additionally, the more rapid appearance of the dextran tracer in ipsilateral venous blood after intra-lymphatic injection in a treated limb suggested a functional connection. We recognize that these data are very limited numbers, but if supported by additional studies, these connections could have important implications. First, these lymph–venous connections could, in part, help to explain the lack of correlation between the lymphatic functional state and the bioimpedance data as is discussed below. Second, it may be possible to take advantage of this model to enhance this process with lymphangiogenic or other therapeutic approaches. This would offer an invaluable opportunity to study the physiological and molecular factors that facilitate the formation of these new junctions, which presumably work to enhance fluid clearance from the limb in response to the stress of downstream lymph flow obstruction.
Paradoxical findings related to bioimpedance measures of tissue water
Once the lymphatic system's capacity to remove protein and water from the interstitium is compromised, edema develops.46,47 In this study, radiation impaired lymphatic function as evidenced by reduced transport of the intra-lymphatic tracer to plasma and as altered inflow pressure–flow relationships. Nonetheless, the association between lymphatic dysfunction and tissue edema was less clear. While not statistically significant, our data demonstrated a counter-intuitive trend to reduced tissue water after radiation was applied to the tissues in concert with impaired lymphatic function as noted above. A number of factors may have contributed to this effect.
A major issue was the variability of the bioimpedance data. While measurements of tissue water with bioimpedance is well accepted,48,49 the method in our hands was quite sensitive to needle location and it was difficult to ensure the identical placement of the needles between the pre- and post-treatment measurements. However, it should be noted that we rarely observed visible edema in the rabbit limbs of the treated animals (all groups). This is in contrast to our experience with sheep where the removal of the popliteal lymph node produced a visible edema consistently. We must also reflect on the biological variability and the individual compensatory mechanisms marshaled in response to the radiation or surgical insult.
Second, in some dog and rat hind limb studies, the authors concluded that radiation alone was not sufficient to induce any permanent edema.20,21,50,51 Therefore, in our study it is possible that any elevation in tissue water in the irradiated group was largely subclinical. Additionally, the reduced tissue water observed at 6 months may be due to the presence of large scar in the treatment area as collagen binds less water). 52 Tissue fibrosis in one part of the limb might mask or cause an underestimation of elevated water (possibly subclinical) in other areas.
A final point to consider is the possible disconnect between our measurements of lymphatic function and global limb drainage. Our lymph function measurements were based on the intra-lymphatic infusion of tracer or artificial lymph. We observed a significant reduction in transport capacity and changes in the pressure–flow relationships that signaled increased resistance to flow. Nonetheless, at the same time, we found evidence of vigorous new vessel formation in the limb and new lymph–venous connections, both of which would work to bypass the obstruction in the popliteal fossa.
Therefore, it is possible that the capacity of the lymphatic system to regenerate new drainage pathways and find new upstream venous connections helped to maintain tissue fluid removal from the limb in the face of compromised flow through the vessels from which we were taking measurements. This may be especially true in the combination treatment groups (radiation plus node removal) in which one would expect to see the greatest degree of lymphatic dysfunction. This group exhibited the greatest evidence for new lymphatic vessel growth.
Global impact of radiation on lymphedema
Guyton and colleagues have described the concept of a ‘safety factor’ that works to prevent increases in interstitial fluid volume. 53 The normal negative limb interstitial fluid pressure provides some hydrostatic buffering effect as interstitial fluid pressure must rise before edema develops. Negative interstitial pressures exist as long as transcapillary fluid flux and lymph transport are at appropriate levels to maintain the interstitium in a relatively dehydrated state. As interstitial fluid increases, lymph flow rates rise to remove fluid from the interstitial compartment. If lymph transport is impaired, the magnitude of this ‘edema safety factor’ is reduced. The probability that edema may occur is increased because the threshold interstitial fluid pressure reflective of clinical edema would be more easily achieved if interstitial fluid pressures were allowed to rise chronically. The wide range of baseline transport rates of the control limbs in this study suggest normal lymphatic functionality levels differ from animal to animal. This phenomenon would most likely be recapitulated in human populations and is a probable explanation as to why only some patients develop lymphedema, while others do not.
Radiation undoubtedly contributes to the erosion of this ‘safety factor’ and may occasionally produce lymphedema by itself. For example, in a clinical study on a group of breast cancer patients unable to undergo surgical treatments (inoperable tumors, unable to undergo general anesthetics etc.), 11% of women developed arm edema in the following 3 and half years, 54 suggesting that ionizing radiation alone has the ability to impact the lymphatic system negatively. In any event, radiation is likely to have a complex effect on the interstitial–lymphatic system. In the study reported here, we observed that much higher pressures were required to establish a given flow rate through lymph nodes after the application of radiation. This would imply that the initial insult to the node (possibly radiation-induced fibrosis) impaired the velocity of fluid flow from the interstitium into the initial lymphatic vessels, a phenomenon that is a central tenet of a novel interstitial lymphedema hypothesis. 55 Consequently, elevated hydrostatic pressure differences between the interstitium and lymphatics would be necessary to maintain lymph flow at a given level through the system. Additionally, the functional deficit caused by radiation may introduce hemodynamic factors, which could exacerbate the accumulation of fluid in the interstitium. Increased capillary filtration due to vasodilation or the formation of new blood vessels has been suggested to be a significant contributor to edema in cancer-related lymphedema.55,56
Whatever the mechanism responsible for radiation's effect, it seems likely that multiple ‘hits’ to the system are necessary for edema formation. The cumulative damage induced by radiation, lymph node removal and possibly chemotherapy as well may push the interstitial pressure beyond the edema threshold. In addition, some patients with unilateral lymphedema have abnormal lymph scintigraphy in the clinically normal limb, suggesting some underlying lymphatic structural anomaly that may predispose the individual to clinical edema (possibly genetic susceptibility). 57 On the other side of the ledger, the ability to form new lymphatics and generate lymphatic–venous anastomoses appears to compensate to some extent for the pathological events that facilitate edema formation.
Conclusions
The data in this study demonstrate that radiation of a lymph node inhibits lymph transport significantly. The fact that the magnitude of functional suppression following lymph node excision plus irradiation is similar to irradiation alone, suggests that a major portion of the injury to the system was caused by the radiation itself. Collagen deposition in the node resulting in a thickened nodal capsule may have contributed to this functional deficit. The edema associated with lymph node removal or irradiation in the rabbit model appears to be largely subclinical. However, there was a tendency for tissue water to decrease in the irradiated limbs. This was likely due to fibrosis in the affected tissues. The development of new lymphatic vessels and lymph–venous anastomoses indicated a vigorous attempt by the host to compensate for the lymph flow impediment by establishing collateral pathways for drainage.
Footnotes
Author Disclosure Statement
The author(s) declare that they have no competing financial interests.
This project was funded by the Canadian Breast Cancer Foundation–Ontario Chapter. Student funding was provided by through a Frederick Banting and Charles Best Canada Graduate Scholarship from the Canadian Institute of Health Research. The Budd Sugarman Foundation generously provided the bioimpedance device. JLS is supported by the Canadian Breast Cancer Foundation (Ontario) as the Chair in Surgical Research (for CBCF).