
Abstract
Abstract
Background:
The lymphatic system is commonly injured during cancer treatment. However, despite the morbidity of these injuries, there are currently no options for replacing damaged lymphatics. The purpose of this study was to optimize methods for decellularization of murine lymph nodes (LN) and to determine if these scaffolds can be used to tissue engineer lymph node-like structures.
Methods and Results:
LNs were harvested from adult mice and subjected to various decellularization protocols. The degree of decellularization and removal of nuclear material was analyzed histologically and quantitatively using DNA isolation. In addition, we analyzed histological architecture by staining for matrix proteins. After the optimal method of decellularization was identified, decellularized constructs were implanted in the renal capsule of syngeneic or allogeneic recipient mice and analyzed for antigenicity. Finally, to determine if decellularized constructs could deliver lymphocytes to recipient animals, the matrices were repopulated with splenocytes, implanted in submuscular pockets, and harvested 14 days later. Decellularization was best accomplished with the detergent sodium dodecyl sulfate (SDS), resulting in negligible residual cellular material but maintenance of LN architecture. Implantation of decellularized LNs into syngeneic or allogeneic mice did not elicit a significant antigenic response. In addition, repopulation of decellularized LNs with splenocytes resulted in successful in vivo cellular delivery.
Conclusions:
We show, for the first time, that LNs can be successfully decellularized and that these matrices have preserved extracellular matrix architecture and the potential to deliver leukocytes in vivo. Future studies are needed to determine if tissue engineered lymph nodes maintain immunologic function.
Introduction
T
Decellularized allografts have gained significant favor as a means of developing three-dimensional biologic scaffolds that can be repopulated to restore complex organ function. These scaffolds are efficacious because they preserve cellular binding sites and tissue-specific growth factors that are necessary for rapid cellular migration, orientation, and differentiation. For example, recent studies have reported successful development of digestive organs, heart valves, nerves, liver, bladder, and trachea.1–3 Interestingly, however, this approach has not been applied to tissue engineering of immune organs.
The purpose of the current study was to optimize protocols to decellularize murine lymph nodes, to assess the potential of these scaffolds to elicit immune responses after allogeneic transplantation, and to determine if this technique can be used to deliver immune cells after in vivo implantation.
Methods
Lymph node harvest and decellularization
Axillary and brachial lymph nodes were harvested from adult female (12–16-week old) C57B6/J mice (Jackson Laboratories, Bar Harbor, Maine) under a protocol approved by the IACUC committee at Memorial Sloan-Kettering Cancer Center. The explanted lymph nodes were washed in sterile, ice-cold PBS to remove blood and debris followed by various decellularization protocols (outlined below).
A review of the literature provided a variety of protocols for tissue decellularization including the use of detergents such as sodium dodecyl sulfate (SDS), 3-cholamidopropyldimethylammonio-1-propanesulfonate (CHAPS), and Triton X (4-11). In general, each of these protocols have advantages and disadvantages including preservation of tissue architecture, efficacy of cellular removal, and denaturation of endogenous DNA. Therefore, to define the optimal method for lymph node decellularization, we subjected freshly explanted lymph nodes to these various protocols, and within each protocol, we optimized concentrations of the active ingredients to obtain the best balance between decellularization and maintenance of tissue architecture (Table 1).
Decellularization with SDS was performed using a modification of the methods reported by Schaner et al. 12 SDS (Fischer Scientific, Fair Lawn, NJ) was tested in a variety of concentrations ranging from 0.075 to 1.0% (prepared in phosphate-buffered saline (PBS)). Lymph nodes were continuously agitated at 37°C for 16 h, washed with PBS, and then further digested with or without DNase I (0.2 mg/mL; Roche Diagnostics, Mannheim, Germany) for 20 min.
Decellularization with CHAPS was performed using a modification of the methods reported by Petersen et al. 10 Briefly, 8 mM CHAPS solution was prepared in an alkalinized PBS solution and the lymph nodes were incubated at 37°C with continuous agitation for 12 h. Following this, the tissues were washed in ice cold PBS and then further treated with or without DNase I (0.2 mg/mL).
Decellularization with 0.25% Triton X-100 (Fischer Scientific, Pittsburgh, PA) and 0.25% deoxycholic acid (DOC; Alfa Aesar Ward Hill, MA) was performed using a modification of the methods reported by Rieder et al. 13 Briefly, lymph nodes were digested in a solution of 0.25% Triton X/0.25% DOC for 24 h at 37°C. Tissues were subsequently washed in sterile PBS at 4°C for 24 h with serial changes of PBS. A subset of these nodes were then subjected to a 20 min incubation in a 0.2 mg/mL DNase I in PBS solution. These lymph nodes were then washed for 24 h in PBS at 4°C.
Histology
Normal (i.e., nondecellularized) and decellularized lymph nodes were fixed in 4% paraformaldehyde (PFA; Affymetrix, Cleveland, OH), embedded in paraffin, sectioned at a thickness of 5 μm, and stained with Hematoxylin and Eosin using standard protocols. Additionally, in order to analyze tissue architecture, tissue sections were stained for reticulum using the Gordon and Sweets Method. 14 Briefly, tissue sections were deparaffinized, hydrated, and oxidized in potassium permanganate, followed by bleaching in oxalic acid. Next, the sections were sensitized in ferric ammonium sulfate, stained with ammoniacal silver nitrate solution, reduced in 10% formalin, then toned with gold chloride and sodium thiosulfate. In addition, we performed immunohistochemical localization of type I collagen to assess for preservation of collagen scaffold structures in decellularized lymph nodes using a rabbit polyclonal anti-mouse type I collagen antibody (Abcam, catalog #AB292; Cambridge, England). Images were captured (Mirax Scanner, Carl Zeiss, Munich, Germany) and collagen density was analyzed using Metamorph Offline software (Molecular Devices; Sunnyvale, CA). Finally, in order to analyze tissues for DNA contamination, frozen sections of decellularized lymph nodes were stained with 4′,6-diamindino-2-pheylindole (DAPI), and visualized with an Axioscope (Carl Zeiss; Oberkochen, Germany). These methods enabled us to identify the most efficacious method of lymph node decellularization. Once this was done, the remainder of the experiments were performed only on this method in an effort to limit extraneous information.
DNA quantification
The DNA content of decellularized lymph nodes was assessed using the Qiagen DNeasy Blood and Tissue kit (Qiagen, Valencia, CA). Briefly, normal and decellularized lymph nodes were enzymatically lysed using proteinase K. The resulting lysate was then passed through a selective DNA binding membrane. DNA was eluted and spectrophotometric analysis was performed on the purified DNA.
Subcapsular renal implantation
In an effort to determine the antigenic potential of decellularized lymph nodes, tissues were decellularized, mechanically lysed, and then injected in the renal subcapsular space of syngeneic (C57B6) or allogeneic (BALB-C) recipient mice using a 31 gauge Hamilton Syringe (n=4 in each group). The location of tissue injection was marked with India Ink. Recipient animals were sacrificed 14 days after transplantation and the kidneys were harvested. Tissues were then fixed with 4% paraformaldehyde, paraffin embedded, and sectioned. Inflammation was analyzed using H&E staining as well as immunohistochemistry for CD45 (Ab114; R&D Systems, Minneapolis, MN) a pan-leukocyte marker. Cell counts of CD45+ cells/high powered field (HPF) were calculated by two blinded reviewers.
Intramuscular decellularized lymph node implantation
In order to determine if decellularized lymph nodes could be repopulated with immune cells and if these cells survive implantation, we harvested lymph nodes from donor mice, decellularized them using SDS, and repopulated these lymph nodes with leukocytes harvested from the spleen of a C57B6-GFP transgenic donor mouse (C57BL/6-Tg(UBC-GFP)30Scha/J; Jackson labs). Briefly, the spleens of GFP mice were fractured, filtered through a 40 μm cell strainer, and treated with red blood cell lysis buffer (Ebioscience; San Diego, CA). The cells were then resuspended in PBS and counted. Decellularized lymph nodes were repopulated with 106 GFP+ leukocytes using a 31-gauge Hamilton syringe and then implanted in an intramuscular pocket in the adductor magnus muscle. The implantation site was marked and the skin was closed over the implantation site. Animals were subsequently sacrificed 14 days later and the muscle with the lymph node were excised en bloc. The tissue was embedded in optimal cutting temperature compound (O.C.T. Compound, Tissue Tek, Sakura, Torrance, CA), sectioned at a thickness of 5 microns using a cryostat (Leica, Buffalo Grove, IL) and mounted on Fisher Plus slides (Fisher, Pittsburgh, PA). GFP-positive cells were identified using a fluorescent microscope, and cellular nuclei were stained with Hematoxylin. Images were captured using an Axioscope (Carl Zeiss).
Statistical analysis
Statistical analysis was performed using the Student's t-test for comparisons between two groups. All data are presented as mean±standard deviation unless otherwise noted. All experiments were performed using a minimum of 5 animals per group.
Results
SDS is the most effective method of decellularizing murine lymph nodes
Comparison of histologic sections of decellularized lymph nodes with normal lymph nodes demonstrated that 0.075% SDS was the most effective method (Fig. 1). This process yielded virtually complete absence of cellular material and maintenance of lymph node architecture based on H&E staining. In contrast, decellularization with various concentrations of CHAPS or Triton-X primarily resulted in decellularization of the cortical regions but not medullary areas of the lymph node. Increasing concentration or time of digestion with CHAPS or Triton-X resulted in significant tissue disorganization and loss of architectural structure (not shown). The addition of DNase to the digestion process did not further improve decellularization with SDS, CHAPS, or Triton-X. In fact, DNase digestion (even when used at low concentrations or performed over relatively brief periods of time) resulted in varying degrees of structural distortion and loss of extracellular matrix proteins (not shown).
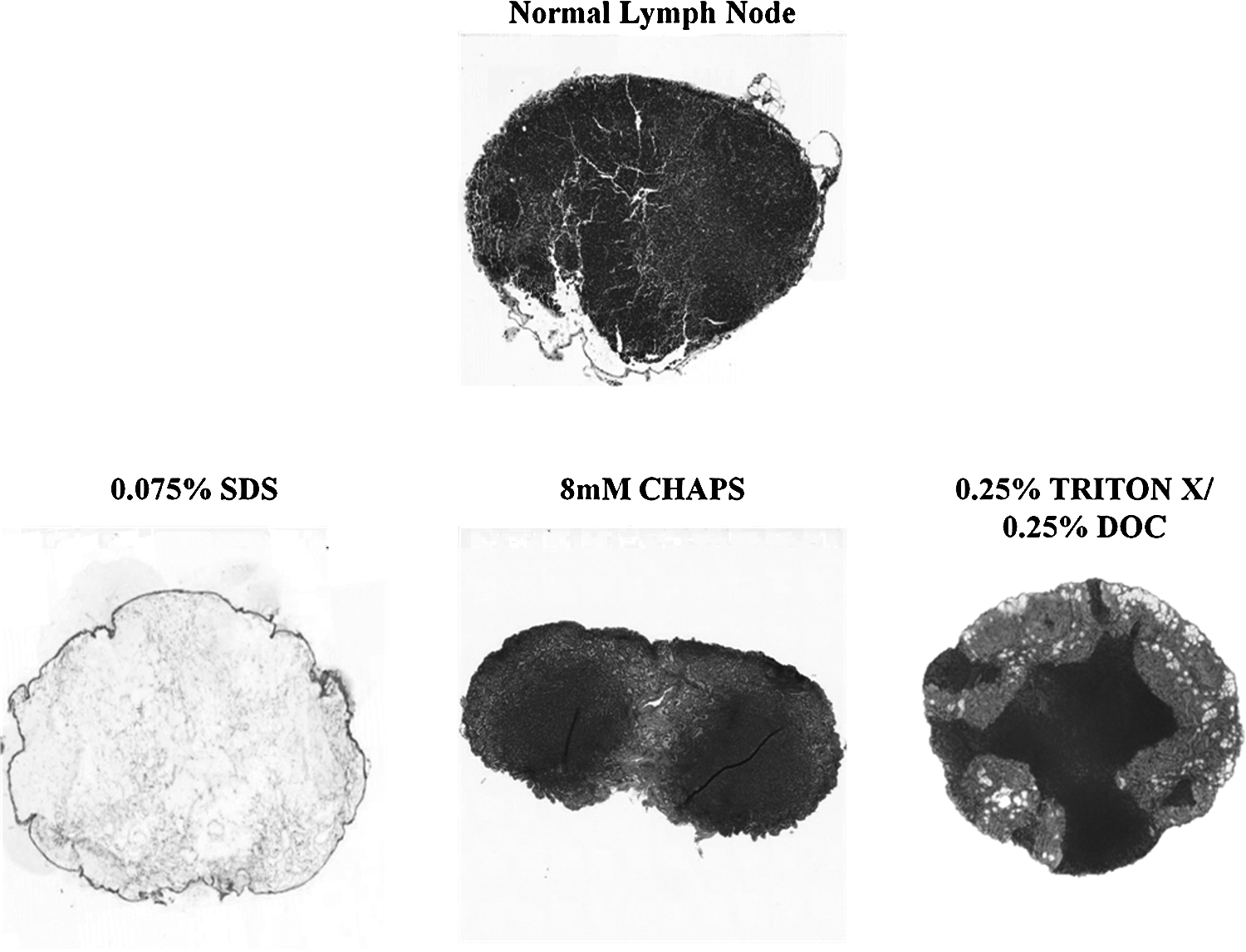
SDS is the most effective method of decellularizing murine lymph nodes. Representative histological sections of normal (i.e., nondecellularized) lymph node (top) and lymph nodes decellularized with 0.075% SDS (lower left panel), 8 mM CHAPS (lower middle panel), or 0.25% Triton-X (lower right panel). Note complete decellularization only in SDS-treated nodes. Sections are shown at 5X magnification and are representative of at least 5 lymph nodes per group.
Treatment with SDS removes nuclear material in murine lymph nodes
To determine how effective various decellularization protocols were in removing nuclear material, decellularized tissue sections were stained with DAPI a fluorescent marker that binds DNA (Fig. 2a). Removal of DNA and nuclear material is a critical step in decellularization since these components can elicit significant immune responses by activating innate immunity via toll like receptor (TLR-9) activation. 15 As expected, analysis of normal lymph nodes stained with DAPI demonstrated densely concentrated cells. In contrast, analysis of lymph nodes decellularized with SDS demonstrated virtually no DAPI staining. As expected from our histological analysis, tissues decellularized with CHAPS or Triton-X had varying amounts of DAPI staining, although cellular localization was lost, suggesting that these treatments were successful in disrupting cellular membranes but not removing nuclear material.
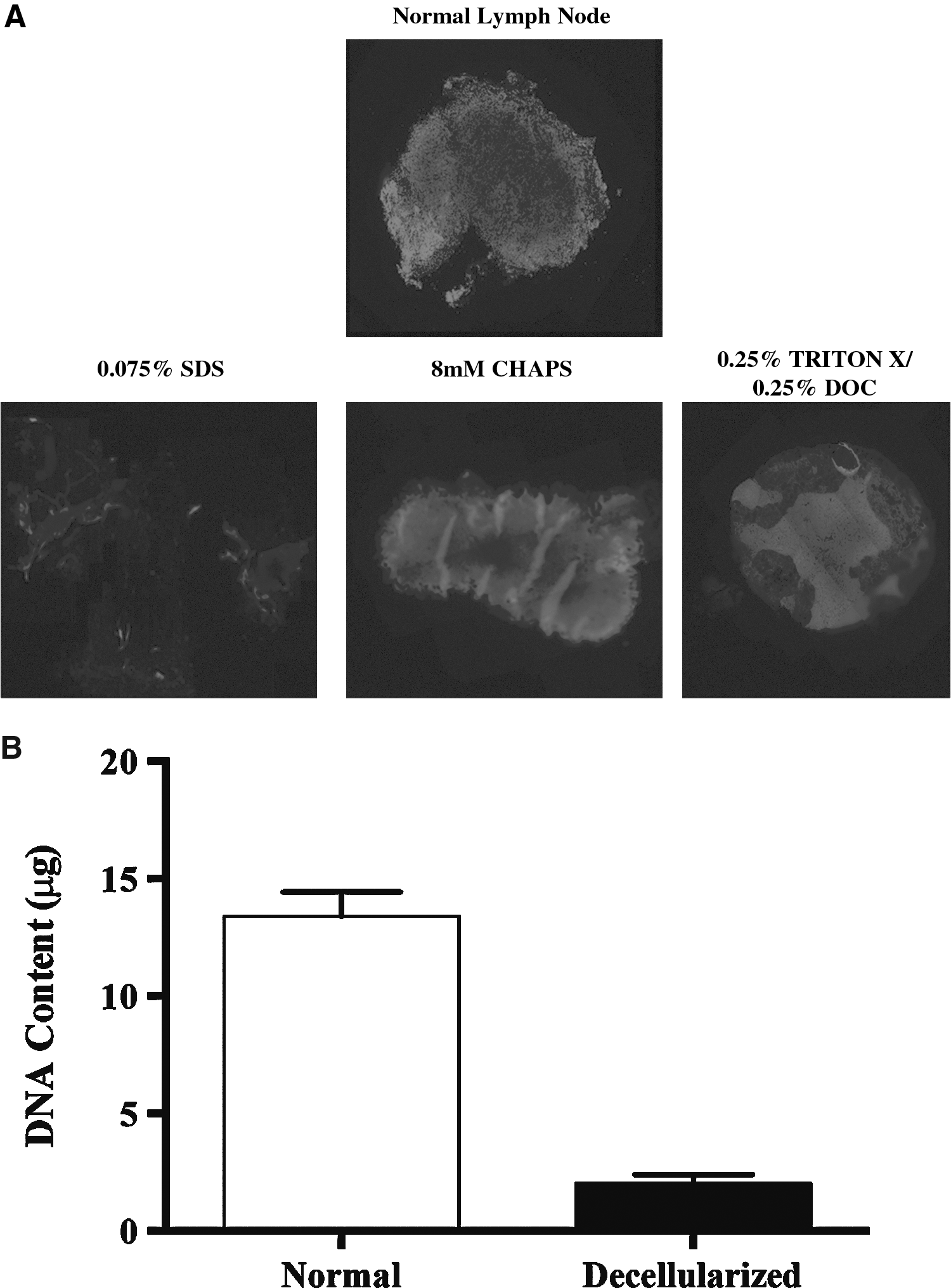
Treatment with SDS removes nuclear material in murine lymph nodes.
We confirmed our DAPI staining by quantifying the amount of DNA present in tissues decellularized with SDS (Fig. 2b). This analysis demonstrated a significant reduction in DNA content of SDS decellularized lymph nodes as compared with normal lymph nodes (p<0.0001).
Lymph nodes decellularized with SDS have preserved tissue architecture
While effective tissue decellularization and removal of nuclear material is a critical first step in developing a potentially viable tissue matrix, this process has to be balanced with preservation of tissue architecture to enable cellular binding and maintenance of three-dimensional structure. Reticulin is a connective tissue network made up of Type III collagen and other extracellular matrix proteins that acts as a supportive structure for cellular components of the lymph node. 16 Therefore, normal and decellularized lymph nodes were stained for reticulin to determine how these treatments preserved the normal tissue architecture of the lymph node. As expected, analysis of normal lymph nodes demonstrated prominent staining for reticulin fibers forming a structural matrix for cellular material in the lymph node (Fig. 3A). This staining was present (though reduced) in lymph nodes decellularized with SDS or Triton-X. In contrast, reticulin staining was almost completely lost in tissues treated with CHAPS. These findings were confirmed with immunohistochemical localization of type I collagen demonstrating near normal collagen architecture of the lymph node. Given our histologic and structural findings demonstrating success of SDS in decellularization, we limited the remainder of our studies to tissues decellularized with SDS only.
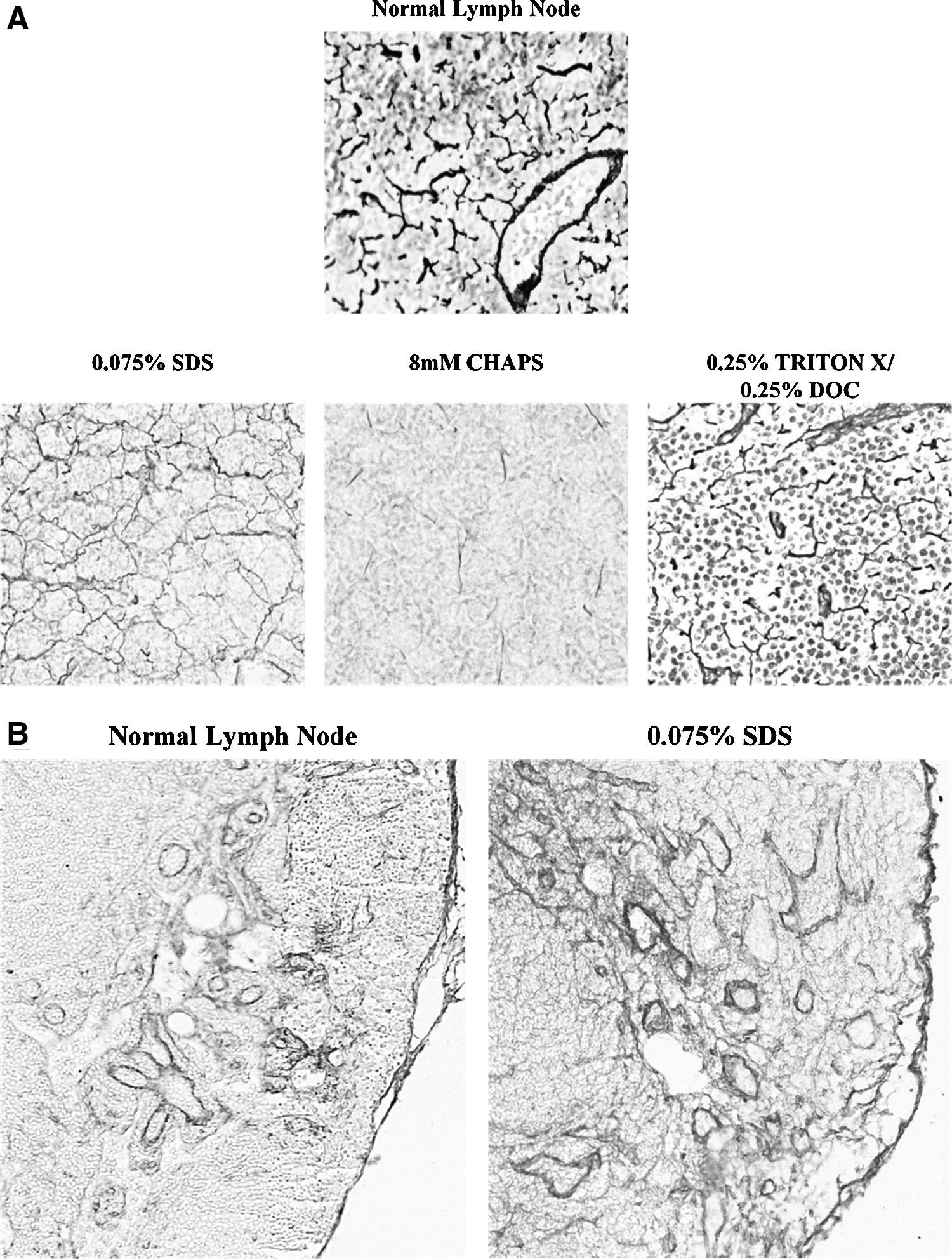
Lymph nodes decellularized with SDS have preserved tissue architecture.
Decellularized lymph nodes do not cause significant tissue inflammation
In order to determine if decellularized lymph nodes elicit an antigenic response, we implanted constructs treated with SDS under the renal capsule of syngeneic (C57B6) or allogeneic (Balb-C) recipient mice. Analysis of histologic sections 14 days after implantation revealed a mild tissue reaction in both groups (Fig. 4a). Further analysis with immunohistochemical localization of CD-45, a pan-leukocyte marker, revealed a similar inflammatory reaction in both syngeneic and allogeneic implants consistent with surgical manipulation but not rejection (Fig. 4b). This finding was confirmed with leukocyte cell counts of high power fields in multiple sections.
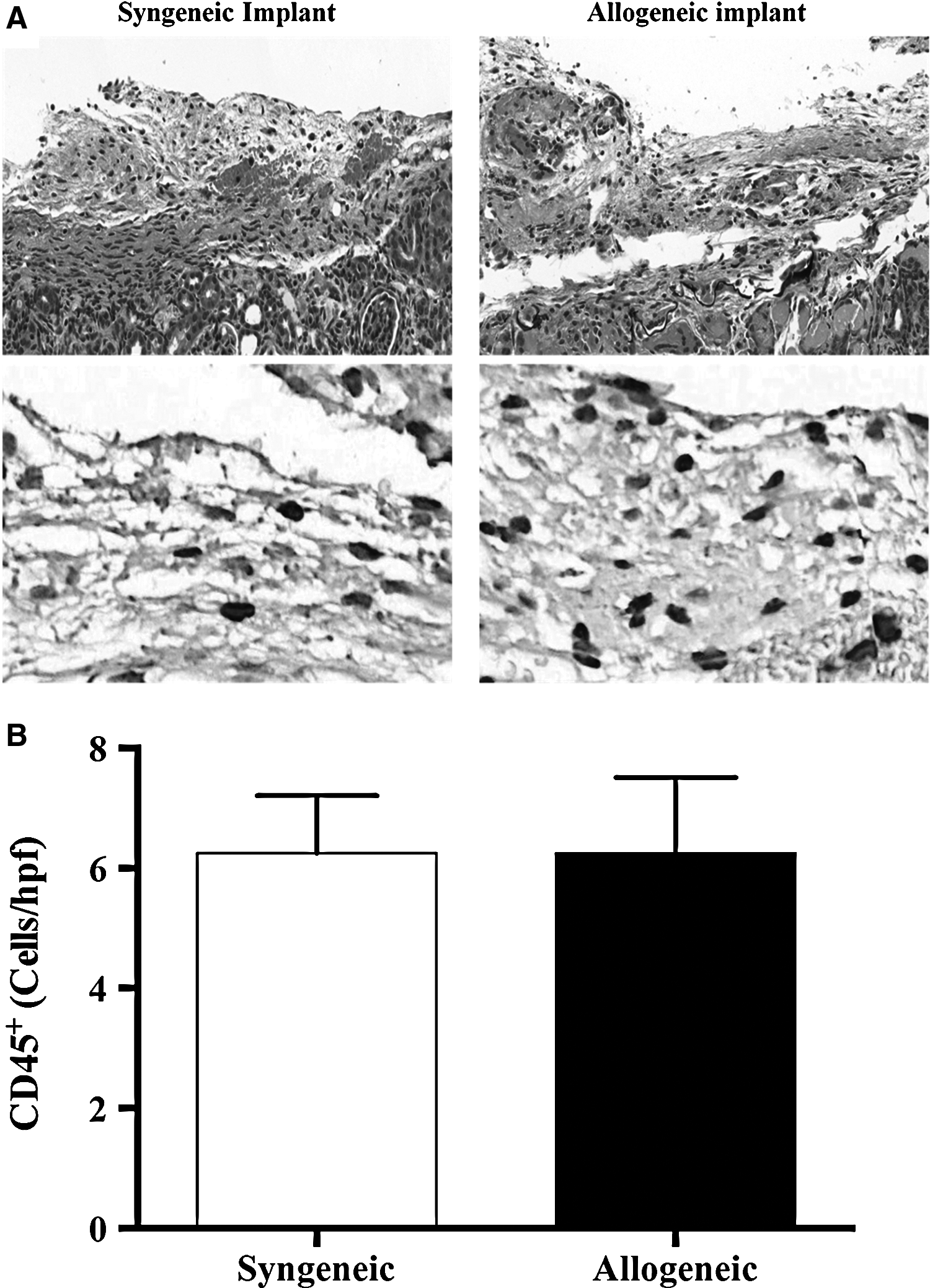
Decellularized lymph nodes do not cause significant tissue inflammation.
Decellularized lymph nodes can be used to deliver immune cells in vivo
A first step in the use of decellularized constructs for production of tissue engineered lymph nodes is to determine if these scaffolds can deliver immune cells in vivo Therefore, to test the hypothesis that lymph nodes decellularized with SDS can deliver immune cells, we injected GFP+ splenocytes into decellularized nodes and implanted them in a muscular pocket for 2 weeks. Interestingly, histological analysis of implanted lymph nodes demonstrated that these scaffolds were completely filled with cells and that the highest concentration of cells was localized to the cortical regions (Fig. 5a). The cells occupying the tissue engineered construct had large nuclei with relatively sparse cytoplasm consistent with lymphocytes. Interestingly, some of these cells appeared to occupy vacuoles in the peripheral regions of the scaffold consistent with the fact that we only injected leukocytes rather than delivering both leukocytes and lymph node fibroblastic reticular cells (FRCs), since the latter are known to regulate lymph node homeostasis and tissue remodeling. 17 This deficiency, as well as the lack of an intact blood supply after transfer, may also explain the fact that the tissue engineered lymph nodes were not completely normal in histologic appearance and lacked the typical follicular pattern.
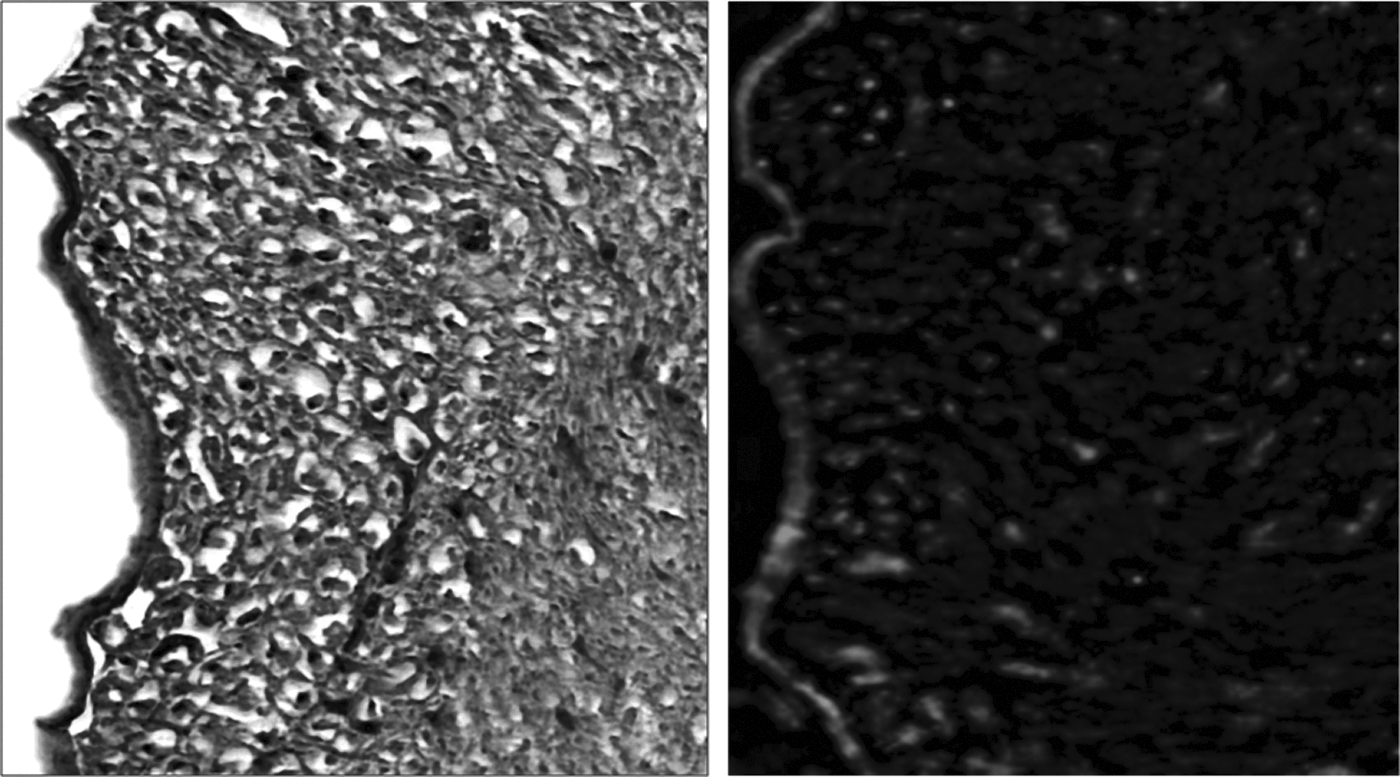
Decellularized lymph nodes can be used to deliver immune cells in vivo. Representative H&E (left panel; 10X magnification) and fluorescence microscopy for GFP (green stain in right panel; 10X magnification) of decellularized lymph nodes reconstituted with GFP+ splenocytes and implanted in a submuscular pocket of syngeneic recipient mice for 14 days. Boxed regions represent a 20X magnification view.
Analysis of tissue engineered lymph nodes with fluorescent microscopy demonstrated distinct cellular staining primarily within the cortical regions of the node confirming that the cells that occupied these regions were of donor origin (i.e., GFP+; Fig. 5b). This finding is important since it suggests that the decellularized construct provides signals that enable cellular survival. Importantly, control sections without cells as well as tissues analyzed with non-GFP fluorescence channels demonstrated no fluorescent labeled cells, suggesting that the cells present in the lymph node are not simply auto-fluorescence (not shown).
Discussion
In the current study we show that lymph nodes can be effectively decellularized and stripped of nuclear content while preserving tissue architecture using SDS. Previous studies have also shown that SDS is effective in decellularization in a number of applications, including tendon, ligaments, bladder, lung, and blood vessels.18–23 Interestingly, when comparing a variety of published techniques for decellularization, we found that other commonly reported methods (e.g., CHAPS or Triton-X) were not very effective in removing cellular material in lymph nodes. Although it is beyond the scope of the current study to determine why these detergents failed to clear cellular material as well as SDS in lymph nodes, we hypothesize that this failure was a consequence of the unique structure of the lymph node (i.e., densely populated organ with delicate extracellular matrix proteins surrounded by a dense fibrous capsule). Thus, it is possible that the dense fibrous capsule of the lymph node prevented adequate diffusion of CHAPs or Triton-X when these detergents were used in low concentrations whereas higher concentrations resulted in loss of architectural proteins. This hypothesis is supported by the pattern of cellular depletion we noted in lymph nodes treated with CHAPS or Triton-X (i.e., mostly depleted in the cortex) as well as previous studies demonstrating that SDS is more effective than Triton X-100 for removing nuclei from dense tissues and organs.8,24,25 An alternative method to achieve decellularization may be in vivo perfusion with detergents thus enabling bypass of the lymph node capsule.9,20 However, this method is likely less clinically applicable due to potential difficulties in perfusion-fixation of human donor tissues.
An important aspect of all tissue engineering is to develop tissues with little or no antigenicity. A key step in this process is to develop methods to remove not only cellular material but also nucleic acids. This is important since the presence of extracellular DNA has been shown to activate innate immune responses. In the current study, we show that digestion of murine lymph nodes with SDS effectively removes both cellular material and DNA as evidenced by DAPI staining and quantification of DNA content. As a consequence, we found that lymph nodes decellularized with SDS elicited only a minimal inflammatory reaction when implanted in either syngeneic or allogeneic recipients.
Although it is important to demonstrate that complex organ systems such as lymph nodes can be effectively decellularized, it is far more important to show that these matrices can deliver cells in vivo. In our study, as a first step towards this goal, we show that donor splenocytes survive in decellularized lymph nodes as long as 14 days after implantation. Although the architecture of these engineered lymph nodes is not yet normal, future studies will determine if lymphangiogenesis can be stimulated in these constructs. This is important since previous studies have shown that organization and differentiation of cells in lymph nodes is highly dependent on the flow of interstitial fluid. 26 We have recently developed a mouse model of lymph node transplantation 27 and plan to use this model in future studies to show that tissue engineered lymph nodes can help restore lymphatic drainage and immune function.
The concept of lymph node tissue engineering has been reported in several other manuscripts in the literature. For example, Suematsu et al. 28 reported on the development of an “artificial lymph node”-like tissue in which bovine collagen scaffolds were seeded with thymic stromal cells and bone marrow-derived dendritic cells and implanted in the renal capsule. This study was exciting as the authors were able to demonstrate that these constructs became populated with lymphoid cells and were able to restore immune function in immunocompromised mice. However, a shortcoming of this approach was the fact that the scaffolds were fragile and due to their xenogeneic nature prone to degradation over time. The use of scaffolds that can retain immunocompetent cells, such as decellularized lymph nodes, is therefore novel and may circumvent these difficulties.
In conclusion, we have shown for the first time that lymph nodes can be effectively decellularized and that these constructs retain minimal amounts of DNA with near normal architecture. Further, we have shown that these constructs elicit only a mild immune reaction in either syngeneic or allogeneic recipients, and that decellularized lymph nodes can be used to deliver immune cells in vivo. Future studies will determine if lymphatic drainage and immune function can be restored by these tissue engineered lymph nodes in animals that have been treated with lymphadenectomy.
Footnotes
Acknowledgments
The authors are grateful to Mesruh Turkekul for expert technical and histologic assistance. We would like to thank the Molecular Cytology Core at Memorial Sloan Kettering Cancer Center for assistance with both histology and tissue imaging.
Author Disclosure Statement
Drs. Cuzzone, Ghanta, and Mehrara and Mr. Albano, and Mr. Aschen have no conflicts of interest or financial ties to disclose. Funding: NIH R01 HL111130-02 awarded to BJM, PSEF PRG 274231 (SKI 10819) awarded to DAC.