
Abstract
Abstract
To address physiological and pathophysiological meanings of condensing effect of albumin in lymph through collecting lymph vessel walls, we established human lymphatic endothelial cells (LEC) and evaluated the size-dependent regulation of the permeability of such layers to hydrophilic substances. We also investigated the effects of tumor necrosis factor (TNF)-α or interleukin (IL)-1β on the permeability and on the morphology of human LEC. Significant amounts of 4 kDa dextran, but not 12 or 66 kDa dextran, passed through the layers. TNF-α or IL-1β induced significant increases in the permeability to 4 and 12 kDa dextrans. TNF-α or IL-1β also produced significant redistribution of the cytoskeletal F-actin in the LEC, which resulted in changes in their shape. Pretreatment with Y-27632, a Rho kinase inhibitor, or PD98059, an extracellular signal-regulated kinase (ERK) phosphorylation inhibitor, significantly abolished the TNF-α- or IL-1β-induced increases in the permeability of the layers to 4 and 12 kDa dextrans. Y-27632 and PD98059 significantly inhibited the changes in the F-actin distribution of the LEC produced by TNF-α or IL-1β. TNF-α or IL-1β caused significant increases in ERK 1/2 phosphorylation in the LEC, which were significantly inhibited by Y-27632 or PD98059. These findings suggest that the human LEC layer plays key roles in the transport of hydrophilic substances through collecting lymph vessel walls and that TNF-α or IL-1β significantly increases the permeability of the layers to 4 and 12 kDa dextrans via Rho kinase activation and the ERK 1/2 phosphorylation-mediated reorganization of F-actin in the LEC.
Introduction
T
In contrast to lymphatic vessels, it is well established that in blood vessels the endothelium of a semi-permeable barrier plays important roles in controlling the passage of macromolecules (more than 66,000 at molecular weight) and fluid between the blood and underlying tissues. 2 It is also known that loss of this function results in tissue inflammation. The characteristic permeability of transported macromolecules is dependent on their molecular sizes as well as the barrier properties of the particular endothelium. Recent studies have suggested that the endothelial cell barrier is regulated by contractile and tethering mechanisms, the effects of which are critically dependent on endothelial cell cytoskeletal components.2,8,9 Thus, Rho kinase families organize the actin cytoskeleton and regulate focal adhesion formation, which result in the regulation of endothelial cell barrier mechanisms.2,10
Therefore, we have attempted to address physiological and pathophysiological roles of lymphatic endothelial cell (LEC) layers in the permeability of human collecting lymph vessel walls to hydrophilic substances, and then to evaluate the effects of tumor necrosis factor (TNF)-α or interleukin (IL)-1β, which are known to be the main cytokines released from activated macrophages in the early stage of acute inflammation, 11 on the permeability of the human LEC layers. Thus, we first established (1) a lymphatic endothelial cell line from the cells that had been isolated from human prenodal collecting lymphatic vessels, and then (2) investigated the permeability of the human LEC or human umbilical vein endothelial cells (HUVEC) layers to fluorescent dye-labeled hydrophilic substances with many kinds of molecular weights. In addition, we (3) evaluated the mechanisms of the effects of TNF-α or IL-1β on the permeability of LEC layers to hydrophilic substances and the distribution and density of cytoskeletal F-actin in the cultured human LEC, and (4) the cellular mechanisms responsible for the cytokines-mediated changes in F-actin distribution and the related cell shape using Western blot analysis.
Materials and Methods
Cell culture
The isolation and culturing of human LEC were performed according to the technique of Kawai et al.4,12,13 The LEC were obtained from the afferent lymph vessels nearest the sentinel lymph node in breast cancer patients without chemotherapy or radiation treatment. The experiments involving human LEC were approved by the ethical committee for human studies of the School of Medicine at Shinshu University. All tissue donors were informed of the risks and purposes of the study before their written consent was obtained. Under a dissecting microscope, the connective and adipose tissue around the target afferent lymph vessel was dissected. The dissected lymph vessel was then cannulated centripetally with a sterile polyethylene tube (diameter: <0.5 mm). First, the lymph vessel was perfused with phosphate-buffered saline (PBS) solution to remove intraluminal lymph fluid. Next, it was intraluminally circulated for 10 min with pre-warmed (37°C) trypsin/ethylenediamine tetraacetic acid (EDTA) solution for endothelial cell cultures (500 U/mL; Catalog No. T4299, Sigma, St. Louis, MO). After the enzymatic digestion, the intraluminal fluid containing the endothelial cells was gently drained into a centrifuge tube containing endothelial growth medium (EGM)-2 (Catalog No. CC-3156, CC-4176, Clonetics, San Diego, CA) supplemented with 10% fetal bovine serum (FBS) and centrifuged at 2000 rpm for 5 min at 4°C. The supernatant was removed, and the resultant pellets were resuspended in EGM-2 culture medium and plated in 35 mm culture plates (Catalog No. 430165, Corning, NY) coated with type I collagen (Catalog No. Type I-P, Nitta Gelatin, Japan). The human LEC were maintained in EGM-2 supplemented with 10% FBS and used at the fifth to ninth passages. At passages fifth to ninth, the cultured human LEC exhibited similar biological properties (e.g., nitric oxide production and release from the LEC) to those reported in in situ studies of collecting lymphatic vessels.4,12 The LEC were incubated under hypoxic conditions of 5% O2, 5% CO2, and 90% N2 at 37°C, 12 because the LEC is known to work under the hypoxic condition (25—40 mmHg). 14
In control experiments, human umbilical vein endothelial cells (Catalog No. CC-2517, HUVEC) were purchased from Sanko Junyaku (Tokyo, Japan). The cells were also maintained in the EGM-2 together with 10% FBS. The HUVEC were incubated under normoxic conditions of 21% O2, 5% CO2, and 74% N2 at 37°C.
Cell barrier permeability assay
The permeability of the human LEC or HUVEC layers to fluorescein isothiocyanate (FITC)-labeled dextran molecules with mean molecular weights of 4,400 (4 kDa, Catalog No. FD-4), 12,000 (12 kDa, Catalog No. FD-10S), and 66,000 (66 kDa, Catalog No. FD20-1G) or FITC-labeled albumin (Catalog No. A7015-250MG; Sigma) was assessed using a 24-well cell culture plate insert and polyethylene terephthalate filters with a 1 μm pore size, which were included in the CHEMICON in vitro vascular permeability assay kit (Catalog No. ECM640, BD Biosciences, Durham, NC). Thus, the fluorescent activity of the EGM-2 culture medium solution that had filtered into the lower chamber was determined using a fluorescence plate reader 5 min after the addition of 1 mg/mL of the relevant FITC-labeled dextran or albumin into the upper chamber, which did or did not contain a cultured human LEC layer. A measurement period of 5 min was used in the present experiments because a previous study found that small molecular hydrophilic substances (≤4400) were able to permeate from the intraluminal to extraluminal spaces of isolated rat collecting lymph vessels within 1.5 min. 15 In addition, in the preliminary experiments using the vascular permeability assay kit, the permeability of the human LEC layers to hydrophilic substances with many kinds of molecular weights have been confirmed to reach the plateau level around 5 min.
In the vascular permeability assay, the permeability of the human LEC layer to hydrophilic substances was evaluated in the following manner: An EGM-2 culture medium solution containing an FITC-labeled hydrophilic substance was allowed to pass from the upper to lower chamber of the vascular permeability assay through a human LEC layer, and the level of fluorescent activity in the solution in the lower chamber was assessed using the fluorescent plate reader (CytoFluor 4000) with excitation/emission wave lengths of 485 nm/530 nm at 5 min after the addition of the FITC-labeled hydrophilic substance into the upper chamber. The fluorescent activity was normalized to that of a solution containing the same FITC-labeled hydrophilic substance that had only been passed through a polyethylene filter (pore size: 1 μm) (Normalized FITC-labeled substance flux). In addition, the ratio of the level of fluorescent activity in the solution in the lower chamber at 5 min after the addition of the FITC-labeled hydrophilic substance into the upper chamber to the fluorescent activity of the original test solution was also determined (relative flux of FITC-labeled dextran).
Using the permeability assay kit, the effects of the inflammatory cytokines, tumor necrosis factor (TNF)-α and interleukin (IL)-1β, on the permeability of human LEC layers to hydrophilic substances were also evaluated. Thus, we investigated the effects of 120 min pretreatment with TNF-α (10 or 100 ng/mL) or IL-1β (10 or 100 ng/mL) on the permeability of the LEC layers to 4, 12, and 66 kDa dextrans.
Immunocytochemical examination
Indirect immunocytochemical studies were performed using cultured human LEC seeded on glass slides that had been coated with type I collagen. After being plated on the glass slides, the cells were fixed with 4% paraformaldehyde in PBS solution for 20 min at room temperature, before being washed three times with PBS and then incubated overnight at 4°C with polyclonal human antisera to platelet/endothelial adhesion molecule (PECAM)-1 (dilution: 1:100, Catalog No. 550389, BD Bioscience), lymphatic endothelial hyaluronan receptor (LYVE)-1 (dilution: 1:50, Catalog No. 102-PA50AG, RELIATech, Germany), prospero-related homeobox (Prox) 1 (dilution 1:50, Catalog No. 11-002, AngioBio, CA), podoplanin (dilution: 1:50, Catalog No. 11-031, AngioBio), syndecan 2 (dilution: 1:50, Catalog No. AF2965, R&D Systems, Minneapolis, MN), or FITC-labeled primary polyclonal human antiserum to F-actin (concentration: 10 μg/mL; Catalog No. P5282, Sigma). Before the Prox1 staining, the cultured cells were permeabilized by 5 min-treatment with 0.1% Triton X-100. After the cells had been washed in PBS three times, they were incubated for 1 h at room temperature with 1:100 diluted Alexa Fluor 488 chicken anti-rabbit IgG (Catalog No. A21441) or Alexa Fluor 488 donkey anti-mouse IgG (Catalog No. A21202; Invitrogen, CA). The nuclei of the cultured LEC were counterstained and mounted with Pro Long Gold antifade reagent and DAPI (4′-6-diamidino-2-phenylindole) (Catalog No. P36935, Molecular Probes, Eugene, OR), before being examined with a fluorescent microscope (CTR5000, Leica Microsystems, Wetzler, Germany) and photographed.
To quantitatively evaluate the F-actin immunocytochemical expression in the human LEC, high-resolution digital photomicrographs were processed using the Scion image analysis program. 13 The constant area of each of the LEC was outlined on a gray scale image and processed for density measurement. The results are expressed in arbitrary units (mean density per pixel), and the data are shown as means±SE (n=5).
For nonspecific staining, the primary antiserum was replaced with Block-Ace (Catalog No. UK-B40, Dainippon Sumitomo Pharma, Osaka, Japan) as a negative control.
Western blot analysis
Western blot analysis was performed to quantitatively evaluate the mechanisms responsible for the effects of tumor necrosis factor (TNF)-α or interleukin (IL)-1β on the permeability of the cultured human LEC layer to hydrophilic substances in the presence or absence of the Rho kinase inhibitor Y-27632 or the ERK1/2 kinase inhibitor PD98059. The cells were dissolved in mammalian protein extraction reagent (M-PER, Catalog No. 78503, Thermo Scientific, Rockford, IL) and then centrifuged at 14,000 g for 10 min. A 15-μg sample of the total cell lysate was dissolved in sodium dodecyl sulfate (SDS) sample buffer for SDS-polyacrylamide gel electrophoresis (PAGE) and transferred to a polyvinylidene difluoride membrane (Catalog No. AE-6668, Atto, Tokyo, Japan), where it was incubated for 45 min. The membrane was then probed with rabbit anti-human phosphospecific ERK 1/2 (dilution 1:1,000, Catalog No. 9101S, Cell Signaling Tech, Beverly, MA) or anti-human ERK 1/2 (dilution 1:1,000, Catalog No. 9102, Cell Signaling Tech) and incubated with horseradish peroxidase-conjugated goat anti-rabbit antibody (dilution 1:3,000, Catalog No. 7074, Cell Signaling Tech) for 1 h. The membrane was visualized with an enhanced chemiluminescence Western blotting detection system (Catalog No. RPN2132, Amersham Biosciences, Cambridge, UK).
Drugs
All salts were obtained from Wako (Tokyo, Japan). TNF-α (Catalog No. T0157-10UG), Y-27632 (Catalog No. Y0503), and PD98059 (Catalog No. P215-1MG) were purchased from Sigma. IL-1β (Catalog No. 201-LB-005/CF) was purchased from R&D Systems. Y-27632 was diluted with ethanol. The concentration of ethanol used did not affect the biological viability of the LEC. Drug concentrations are expressed as the final concentration in the culture plate.
Statistical analysis
All results are expressed as mean±standard error of the mean (SEM) values. Statistical analyses were performed using the Student's t-test for unpaired observations or one-way ANOVA, followed by Duncan's post-hoc test, as appropriate. P<0.05 was considered statistically significant.
Results
Establishment of human collecting lymph vessel endothelial cells
We cultured human lymphatic endothelial cells (LEC) that had been isolated from prenodal collecting lymph vessels. Figure 1 shows representative photomicrographs of immunocytochemical staining of PECAM-1, LYVE-1, Prox 1, podoplanin, and syndecan 2 in the human LEC. As shown in the image depicting PECAM-1 staining, most of the human LEC demonstrated polygonal shapes, and as the culture approached confluence each cell was in contact with others. The density of human LEC at confluence was 38.4±2.1/0.1 mm2 (n=6). The doubling time of the LEC was 26.5±2.9 h (n=6).

Representative photomicrographs of immunocytochemical images of platelet/endothelial cell adhesion molecule (PECAM)-1, lymphatic endothelial hyaluronan receptor (LYVE)-1, prospero-related homeobox (Prox) 1, podoplanin, and syndecan 2 in the nonpermeabilized or permeabilized cultured human lymphatic endothelial cells (LEC). The nuclei of the human LEC were counterstained and mounted with ProLong Gold antifade reagent and DAPI (4′-6-diamidino-2-phenylindole) (middle panel). Each merged images is shown in the lower panel. The markers show 50 μm. A color version of this figure is available in the online article at www.liebertpub.com/lrb
The cultured cells were intensely immunostained with antisera against Prox1 and podoplanin, which are selective markers of lymphatic endothelial cells. In addition, the cultured cells also demonstrated a marked immunocytochemical image of syndecan 2 at the borders of the cells, which is known to be a more selective marker of lymphatic endothelial cells in collecting vessels.16,17
Molecular weight-dependent permeability of human LEC layers to hydrophilic substances
Figure 2A shows a schematic diagram of the cell barrier permeability assay used in the present study.
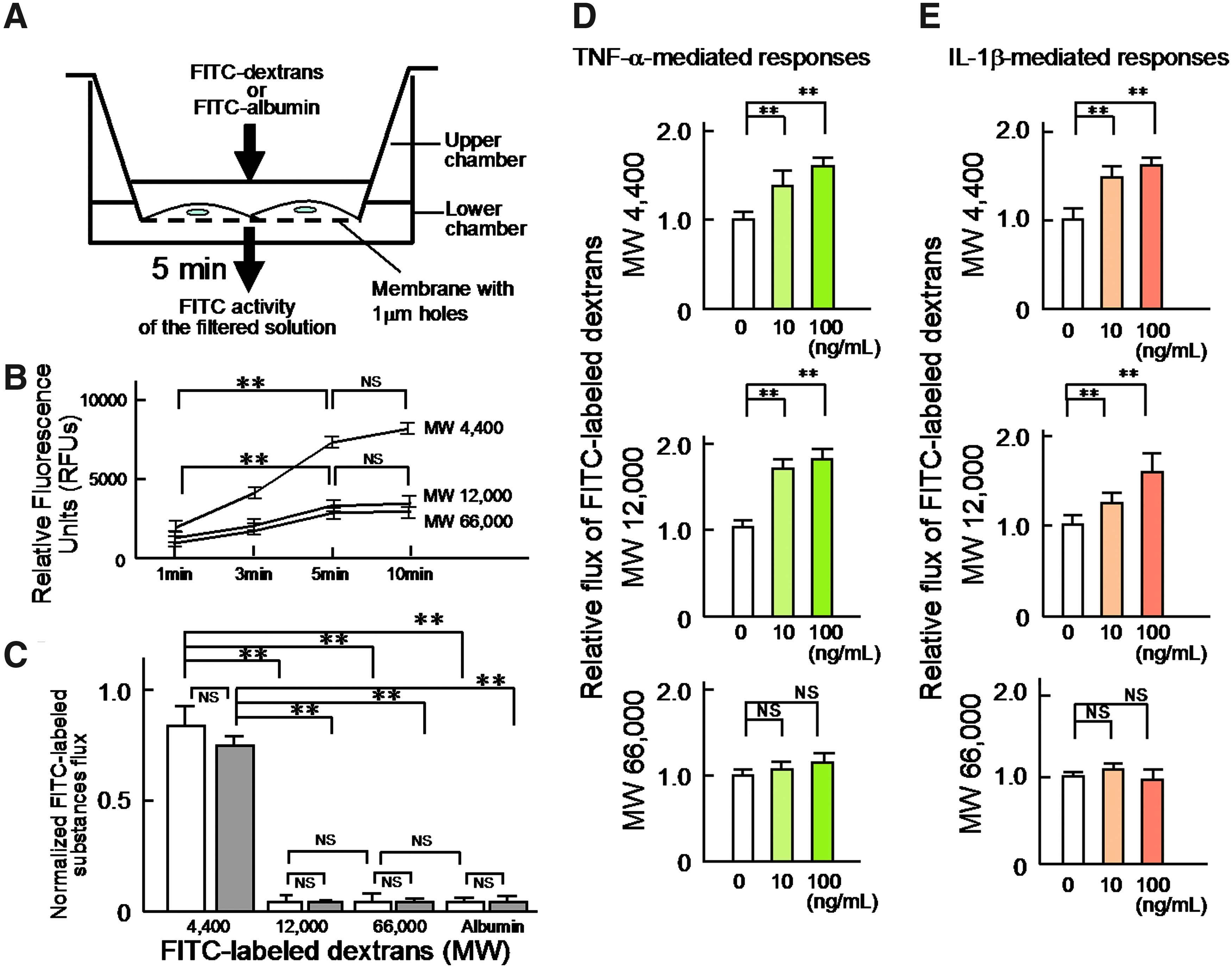
Molecular weight-dependent permeability of the human LEC layers to hydrophilic substances and effects of TNF-α or IL-1β on the permeability of the LEC layers to hydrophilic substances.
The reason why we adopted 5 min of measuring time for the evaluation of the permeability of the human LEC layer to hydrophilic substances is shown in Figure 2B. Thus, the permeability to hydrophilic substances with 4, 12, and 66 kDa evaluated by relative fluorescence units (RFUs) using excitation/emission wave lengths of 485 nm/530 nm and a gain setting of 65 tended to reach the plateau level around 5 min after each addition of the FITC-labeled substance into the upper chamber.
Figure 2C summarizes the findings regarding the permeability of human LEC or HUVEC layers to hydrophilic substances. We found that the human LEC layers (n=9) were permeable to the low molecular weight dextran (mol wt: 4 kDa), whereas significantly lower amounts of the FITC-labeled 12,000 Da (12 kDa) (0.03±0.03, n=9, p<0.01 vs. 4 kDa, 0.76±0.07) and 66,000 Da (66 kDa) (0.04±0.03, n=9, p<0.01 vs. 4 kDa) dextrans and the FITC-labeled albumin (0.03±0.02, n=9, p<0.01 vs. 4 kDa) passed through the LEC layers. Similar findings (n=9) were obtained with the HUVEC layers [4 kDa, 0.86±0.05; 12 kDa, (0.05±0.01, n=9, p<0.01 vs. 4 kDa); 66 kDa, (0.03±0.01, n=9, p<0.01 vs. 4 kDa); albumin, (0.03±0.01, n=9, p<0.01 vs. 4 kDa)].
TNF-α- or IL-1β-induced increases in the permeability of the human LEC layers to 4 kDa and 12 kDa, but not 66 kDa dextrans
Next, we evaluated the effects of TNF-α or IL-1β on the permeability of the human LEC layers to FITC-labeled dextrans (Fig. 2D and 2E). As shown in the panel of Figure 2D, the treatment with TNF-α (10 or 100 ng/mL) caused a significant concentration-dependent increase in the permeability of the layers to 4 kDa and 12 kDa dextrans (Fig. 2D; 4 kDa dextran: 10 ng/mL TNF-α, 1.27±0.11, n=9, p<0.01 vs. no treatment 1.00±0.05, n=9; 100 ng/mL TNF-α, 1.67±0.06, n=9, p<0.01 vs. no treatment) (12 kDa dextran: 10 ng/mL TNF-α, 1.69±0.15, n=9, p<0.01 vs. no treatment 1.00±0.03, n=9; 100 ng/mL TNF-α: 1.76±0.11, n=9, p<0.01 vs. no treatment). In contrast, treatments with TNF-α (10 or 100 ng/mL) caused no significant change in the permeability of the human LEC layers to 66 kDa dextran (10 ng/mL TNF-α, 1.01±0.05, n=9, NS vs. no treatment 1.00±0.03, n=9; 100 ng/mL TNF-α, 1.08±0.07, n=9, NS vs. no treatment).
Similar to the effects of TNF-α on the permeability of the layers to hydrophilic substances, the treatment with IL-1β (10 or 100 ng/mL) produced a significant increase of the permeability of the human LEC layers to 4 kDa and 12 kDa dextrans, but not to 66 kDa dextran in a concentration-dependent manner (Fig. 2E; 4 kDa dextran: 10 ng/mL IL-1β, 1.49±0.07, n=9, p<0.01 vs. no treatment 1.00±0.07, n=9; 100 ng/mL IL-1β, 1.57±0.03, n=9, p<0.01 vs. no treatment) (12 kDa dextran: 10 ng/mL IL-1β, 1.13±0.04, n=9, p<0.01 vs. no treatment 1.00±0.04, n=9; 100 ng/mL IL-1β, 1.61±0.11, n=9, p<0.01 vs. no treatment) (66 kDa dextran: 10 ng/mL IL-1β, 1.04±0.02, n=9, NS vs. no treatment 1.00±0.02, n=9; 100 ng/mL IL-1β, 0.98±0.05, n=9, NS vs. no treatment)
The Rho kinase inhibitor Y-27632 reduced the TNF-α- or IL-1β-induced increases of the permeability of the human LEC layers to 4 kDa and 12 kDa dextrans
To address the role of Rho kinase activity in the TNF-α- or IL-1β-induced increases of the permeability of the human LEC layers to 4 kDa and 12 kDa dextrans, we next investigated the effects of the selective Rho kinase inhibitor, Y-27632 (1 or 10 μM) on the TNF-α (10 ng/mL)- or IL-1β (10 ng/mL)-induced increases of the permeability to 4 kDa and 12 kDa dextrans. The results are summarized in Figure 3A. Thirty-minute pretreatment with Y-27632 significantly abolished the TNF-α- or IL-1β-induced increases in the permeability of the human LEC layers to 4 kDa and 12 kDa dextrans in a concentration-dependent manner [Fig. 3A, left panel (4 kDa dextran, 10 ng/mL TNF-α: 1 μM Y-27632, 1.00±0.06, n=9, p<0.05 vs. no pretreatment 1.27±0.11, n=9; 10 μM Y-27632, 1.02±0.11, n=9, p<0.05 vs. no pretreatment) (12 kDa dextran, 10 ng/mL TNF-α: 1 μM Y-27632, 1.08±0.05, n=9, p<0.01 vs. no pretreatment 1.67±0.06, n=9; 10 μM Y-27632, 1.00±0.04, n=9, p<0.01 vs. no pretreatment)] [Fig. 3A, right panel (4 kDa dextran, 10 ng/mL IL-1β: 1 μM Y-27632, 1.07±0.02, n=9, p<0.05 vs. no pretreatment 1.49±0.07, n=9; 10 μM Y-27632, 1.04±0.03, n=9, p<0.05 vs. no pretreatment) (12 kDa dextran, 10 ng/mL IL-1β: 1 μM Y-27632, 1.06±0.06, n=9, p<0.01 vs. no pretreatment 1.13±0.07, n=9; 10 μM Y-27632, 1.03±0.15, n=9, p<0.01 vs. no pretreatment)].
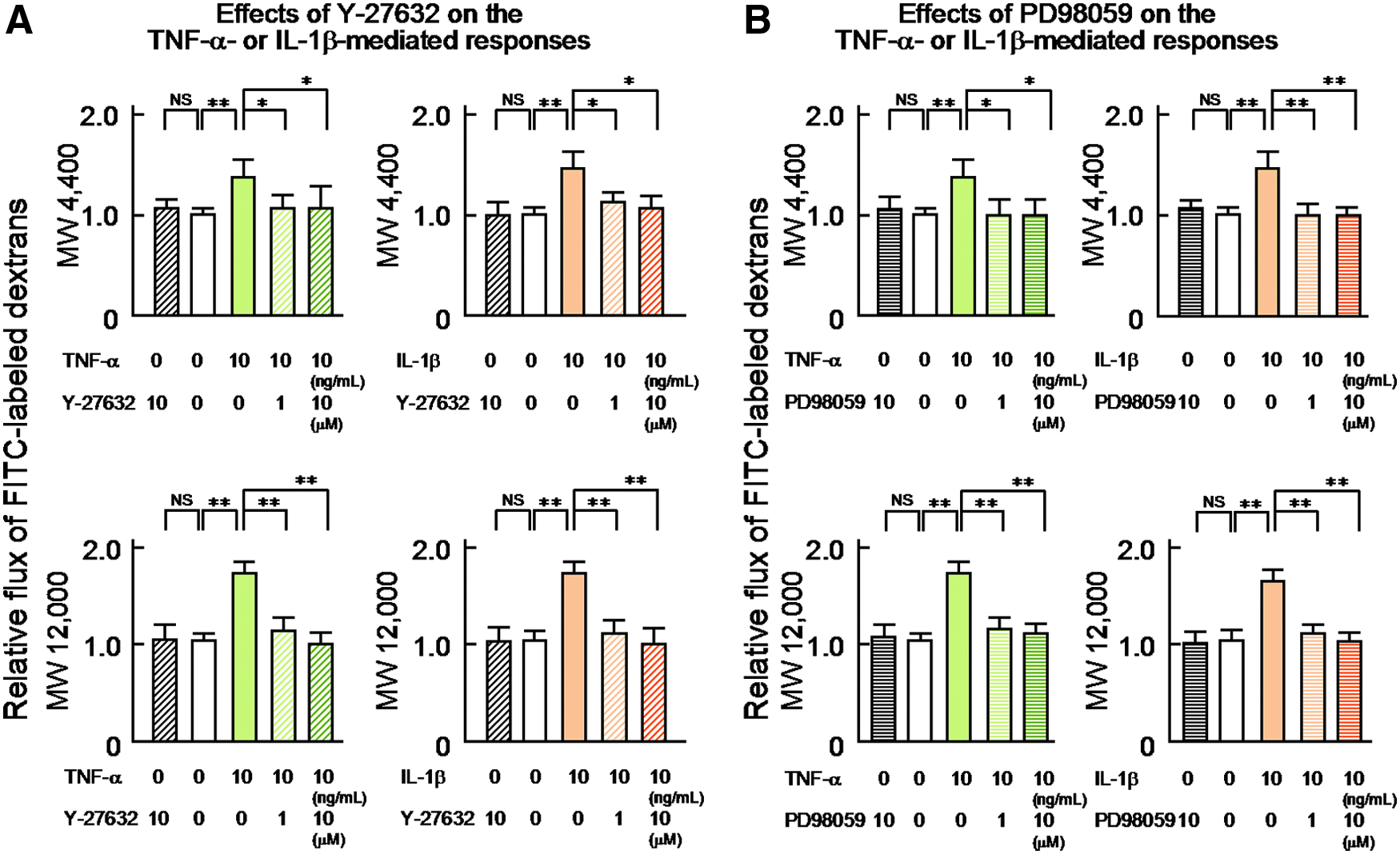
The 30-min treatment with Y-27632 (10 μM) per se produced no significant effect on the permeability of the human LEC layers to 4 kDa and 12 kDa dextrans [Fig. 3A, left panel (4 kDa dextran, 0 ng/mL TNF-α: 10 μM Y-27632, 1.00±0.06, n=9, NS vs. 0 μM Y-27632, 1.00±0.02, n=9) (12 kDa dextran, 0 ng/mL TNF-α: 10 μM Y-27632, 1.01±0.05, n=9, NS vs. 0 μM Y-27632, 1.00±0.02, n=9)], [Fig. 3A, right panel (4 kDa dextran, 0 ng/mL IL-1β: 10 μM Y-27632, 1.01±0.06, n=9, NS vs. no 0 μM Y-27632, 1.00±0.02, n=9) (12 kDa dextran, 0 ng/mL IL-1β: 10 μM Y-27632, 1.02±0.05, n=9, NS vs. 0 μM Y-27632, 1.00±0.03, n=9)].
The ERK 1/2 kinase inhibitor PD98059 reduced the TNF-α- or IL-1β-induced increases of the permeability of the human LEC layers to 4 kDa and 12 kDa dextrans
To evaluate the role of ERK 1/2 activity in the TNF-α- or IL-1β-induced increases of the permeability of the human LEC layers to 4 kDa and 12 kDa dextrans, we next examined the effects of the selective ERK 1/2 kinase inhibitor, PD98059 (1 or 10 μM) on the TNF-α (10 ng/mL)- or IL-1β (10 ng/mL)-induced changes. The results are summarized in Figure 3B. As was found for Y-27632 (1 or 10 μM), 30-min pretreatment with 1 or 10 μM PD98059 caused significant concentration-related reductions in the TNF-α- or IL-1β-induced increases in the permeability of the LEC layers to 4 kDa and 12 kDa dextrans [Fig. 3B, left panel (4 kDa dextran, 10 ng/mL TNF-α: 1 μM PD98059, 0.99±0.04, n=9, p<0.05 vs. no pretreatment 1.27±0.11, n=9; 10 μM PD98059, 0.99±0.06, n=9, p<0.05 vs. no pretreatment) (12 kDa dextran, 10 ng/mL TNF-α: 1 μM PD98059, 1.05±0.03, n=9, p<0.01 vs. no pretreatment 1.67±0.06, n=9; 10 μM PD98059, 1.03±0.04, n=9, p<0.01 vs. no pretreatment)] [Fig. 3B, right panel (4 kDa dextran, 10 ng/mL IL-1β: 1 μM PD98059, 1.01±0.02, n=9, p<0.01 vs. no pretreatment 1.49±0.07, n=9; 10 μM PD98059, 1.01±0.02, n=9, p<0.01 vs. no pretreatment) (12 kDa dextran, 10 ng/mL IL-1β: 1 μM PD98059, 1.04±0.04, n=9, p<0.01 vs. no pretreatment 1.13±0.07, n=9; 10 μM PD98059, 1.00±0.05, n=9, p<0.01 vs. no pretreatment)].
The 30-minute treatment with PD98059 (10 μM) per se caused no significant effect on the permeability of the human LEC layers to 4 kDa and 12 kDa dextrans [Fig. 3B, left panel (4 kDa dextran, 0 ng/mL TNF-α: 10 μM PD98059, 1.02±0.04, n=9, NS vs. 0 μM PD98059, 1.00±0.01, n=9) (12 kDa dextran, 0 ng/mL TNF-α: 10 μM PD98059, 1.04±0.04, n=9, NS vs. 0 μM PD98059, 1.00±0.02, n=9)], [Fig. 3A, right panel (4 kDa dextran, 0 ng/mL IL-1β: 10 μM PD98059, 1.02±0.03, n=9, NS vs. 0 μM PD98059, 1.00±0.01, n=9) (12 kDa dextran, 0 ng/mL IL-1β: 10 μM PD98059, 1.00±0.04, n=9, NS vs. 0 μM PD98059, 1.00±0.04, n=9)].
TNF-α- or IL-1β-mediated cytoskeletal F-actin-dependent changes in the morphology of human LEC
To evaluate the mechanisms responsible for the TNF-α (10 or 100 ng/mL)- or IL-1β (10 or 100 ng/mL)-induced increases of the permeability of the human LEC layers to 4 kDa and 12 kDa dextrans, we investigated the effects of TNF-α (10 or 100 ng/mL) or IL-1β (10 or 100 ng/mL) on the distribution of cytoskeletal F-actin and the related changes in the morphology of cultured human LEC. Figure 4A shows representative photomicrographs of the effects of TNF-α on the distribution of cytoskeletal F-actin and the changes in the morphology of the human LEC. As shown in the photomicrographs of Figure 4A, the treatment with TNF-α produced marked changes in the arrangement and distribution of cytoskeletal F-actin in all of the LEC, which resulted in changes in their shape (i.e., from a cobblestone-like shape to spindle-shaped). Using the ratio of the length of the maximal axis of the every cell to that of the shortest axis that ran perpendicular to the maximal axis as a morphometric parameter (the axis ratio), we quantitatively evaluated the cytoskeletal F-actin-dependent changes in cell shape induced by TNF-α. The results are summarized in Figure 4B. The treatment with TNF-α (10 or 100 ng/mL) caused significant concentration-related decreases in the axis ratio. Furthermore, the reorganization of F-actin into thick filaments was clearly observed at the border of the LEC, which may be, in part, related to change the shape of the LEC from a cobblestone-like shape to spindle shape. In similar to the responses of TNF-α (10 or 100 ng/mL), the treatment with IL-1β (10 or 100 ng/mL) also produced marked changes in the arrangement and distribution of cytoskeletal F-actin in all of the LEC, (Fig. 4C) in the similar changes in shape of the LEC from a cobblestone-like shape to spindle shape. The results are summarized in Figure 4D. Thus, IL-1β caused significant concentration-dependent decreases in the axis ratio of the LEC.

The effects of TNF-α (10 or 100 ng/mL) or IL-1β (10 or 100 ng/mL) on the distribution of cytoskeletal F-actin and the related changes in the morphology of the cultured human LEC.
Y-27632 inhibited the TNF-α- or IL-1β-induced changes in the morphology of the human LEC
To evaluate the role of Rho kinase in the TNF-α (10 or 100 ng/mL)- or IL-1β (10 or 100 ng/mL)-induced morphological changes in the human LEC, we investigated the effects of Y-27632 (1 or 10 μM) on these changes. Figure 5A shows representative photomicrographs of the effects of 1 or 10 μM Y-27632 on the changes in the morphology of the human LEC induced by 10 or 100 ng/mL TNF-α. As shown in the high magnification image of Figure 5A, the treatment with TNF-α produced marked concentration-dependent changes in the shape of the LEC (left-most panel of Fig. 5A). Thirty-minute pretreatment with Y-27632 (1 or 10 μM) significantly inhibited the TNF-α-induced changes in the shape of the human LEC. In addition, the pretreatment with Y-27632 markedly inhibited the organization of cytoskeletal F-actin at the borders of the human LEC, which resulted in maintaining their cobblestone appearance. Consequently, the pretreatment with Y-27632 (1 or 10 μM) abolished significantly the TNF-α (10 or 100 ng/mL)-induced changes in the axis ratio of the LEC. The results are summarized in Figure 5B.
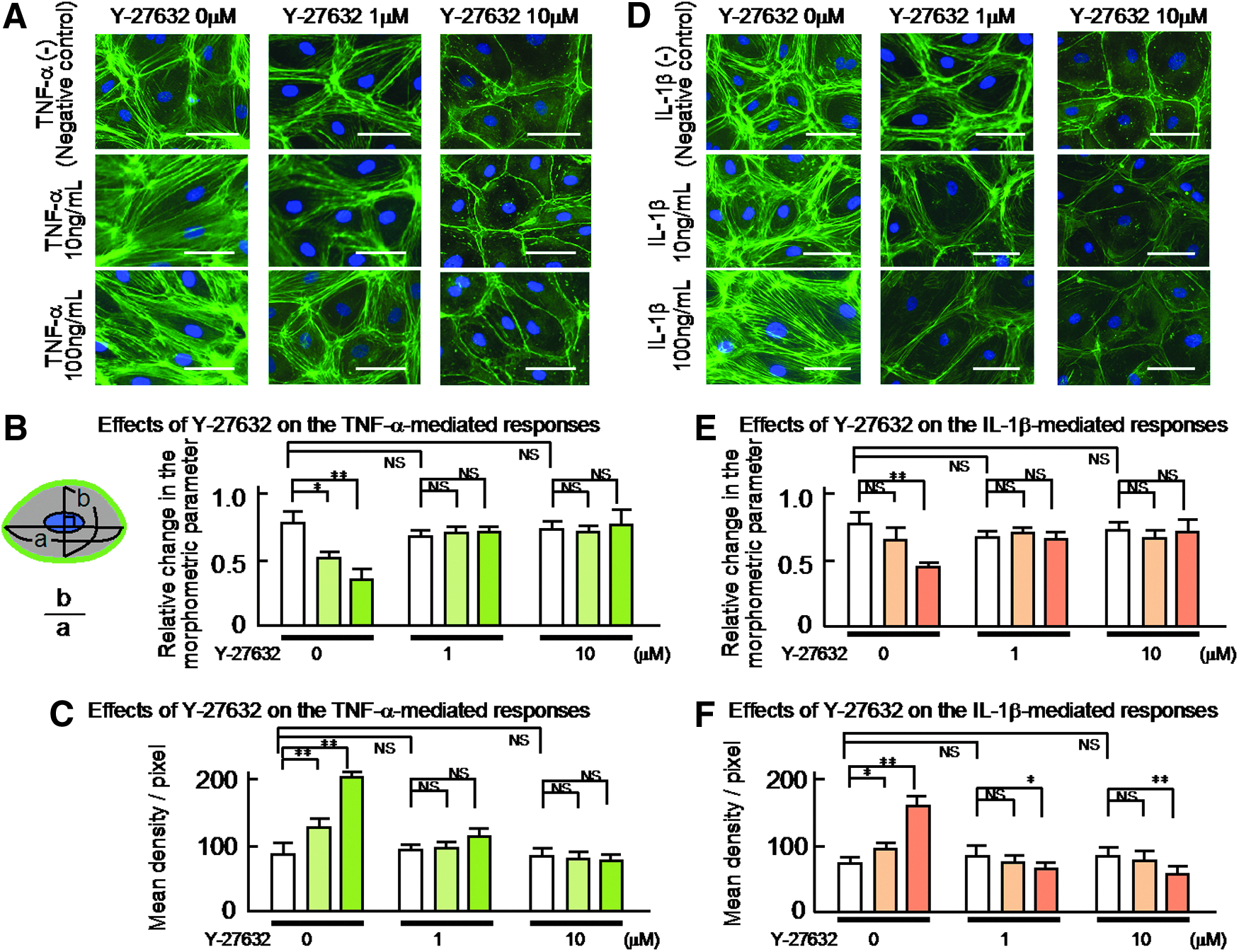
The effects of Y-27632 (1 or 10 μM) on the TNF-α (10 or 100 ng/mL)- or IL-1β (10 or 100 ng/mL) induced the distribution of cytoskeletal F-actin and the related changes in the morphology of the cultured human LEC.
The treatment with Y-27632 (1 or 10 μM) per se produced no significant effect on the morphology of the human LEC (Fig. 5A and D, negative control).
In addition, to examine the relationship between the reorganization of florescence-labeled F-actin and the changes in shape of the LEC, we evaluated again the effects of Y-27632 (1 or 10 μM) on the fluorescent density changes in the human LEC using the density measurements of each photomicrograph obtained by a Scion image analysis program, which may be, in part, related to one of parameters for evaluating the polymerization level of cytoskeletal F-actin in the LEC. The results are summarized in Figure 5C.
In similar to the responses of TNF-α (10 or 100 ng/mL), 30-min pretreatment with Y-27632 (1 or 10 μM) produced marked concentration-related inhibition of the IL-1β (10 or 100 ng/mL)-induced morphological changes of the LEC (Fig. 5D). Thus, the pretreatment with Y-27632 markedly inhibited the reorganization of the cytoskeletal F-actin at the borders of the LEC, which resulted in maintaining their cobblestone appearance. The results are summarized in Figure 5E. Thus, the pretreatment with Y-27632 (1 or 10 μM) abolished significantly the IL-1β-induced changes in the axis ratio of the LEC. The results are also compatible with the data obtained with the density measurements of the LEC (Fig. 5F).
PD98059 inhibited the TNF-α- or IL-1β-induced changes in the morphology of the human LEC
To evaluate the roles of the ERK 1/2 in the TNF-α (10 or 100 ng/mL)- or IL-1β (10 or 100 ng/mL)-induced morphological changes in the human LEC, we next studied the effects of PD98059 (1 or 10 μM) on these changes (Fig. 6A and D). As was found for Y-27632 (1 or 10 μM), the 30-min pretreatment with PD98059 (1 or 10 μM) significantly inhibited the TNF-α- or IL-1β-induced changes in the shape of the human LEC (Fig. 6A and D). The results are summarized in Figure 6B and E. In addition, the pretreatment with PD98059 markedly inhibited the TNF-α- or IL-1β-induced redistribution of cytoskeletal F-actin at the cell border of each human LEC, which resulted in maintaining their cobblestone appearance. The results obtained with the density measurement of the human LEC are also summarized in Figure 6C and F.
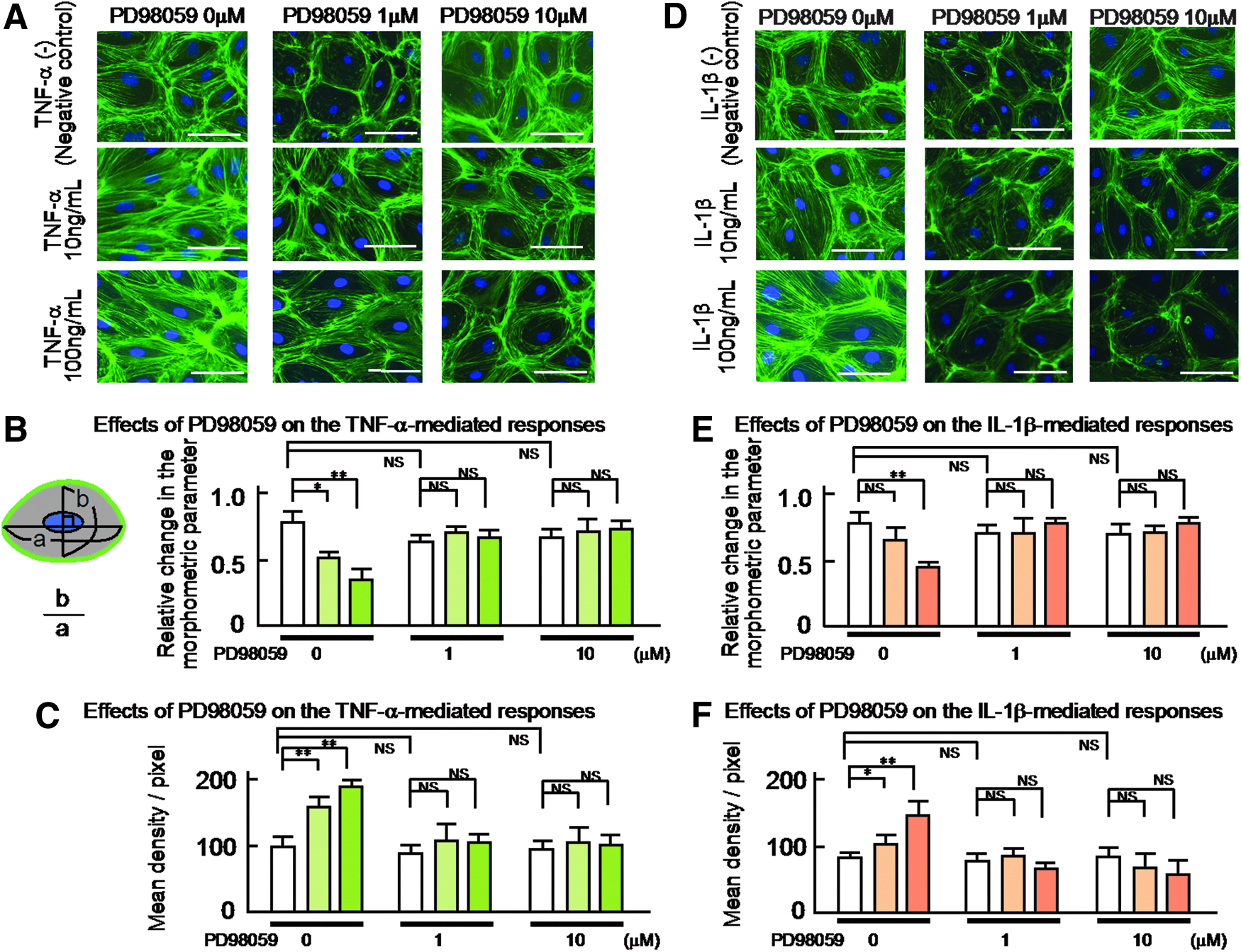
The effects of PD98059 (1 or 10 μM) on the TNF-α (10 or 100 ng/mL) or IL-1β (10 or 100 ng/mL)-induced the distribution of cytoskeletal F-actin and the related changes in the morphology of the cultured human LEC.
TNF-α- or IL-1β-induced phosphorylation of ERK 1/2 in the human LEC
Finally, to evaluate the mechanisms responsible for the TNF-α (10 ng/mL)- or IL-1β (10 ng/mL)-induced Rho-dependent reorganization of cytoskeletal F-actin in the human LEC and the subsequent increases in the permeability of human LEC layers to 4 kDa and 12 kDa dextrans, but not 66 kDa dextran, we studied the phosphorylation of the cell signaling proteins ERK 1/2 in cultured human LEC using Western blot analysis. The results are summarized in Figure 7. As shown in panel A, treatment with 10 ng/mL TNF-α markedly increased the phosphorylation of ERK 1/2 in the LEC, which was partially abolished by additional treatment with 10 μM Y-27632 or completely inhibited by 10 μM PD98059 (Fig. 7A). As shown in the same panel A, the treatment with Y-27632 (10 μM) or PD98059 (10 μM) per se produced no significant effect on the phosphorylation of ERK 1/2 (Fig. 7A). These results are quantitatively summarized in Figure 7B.
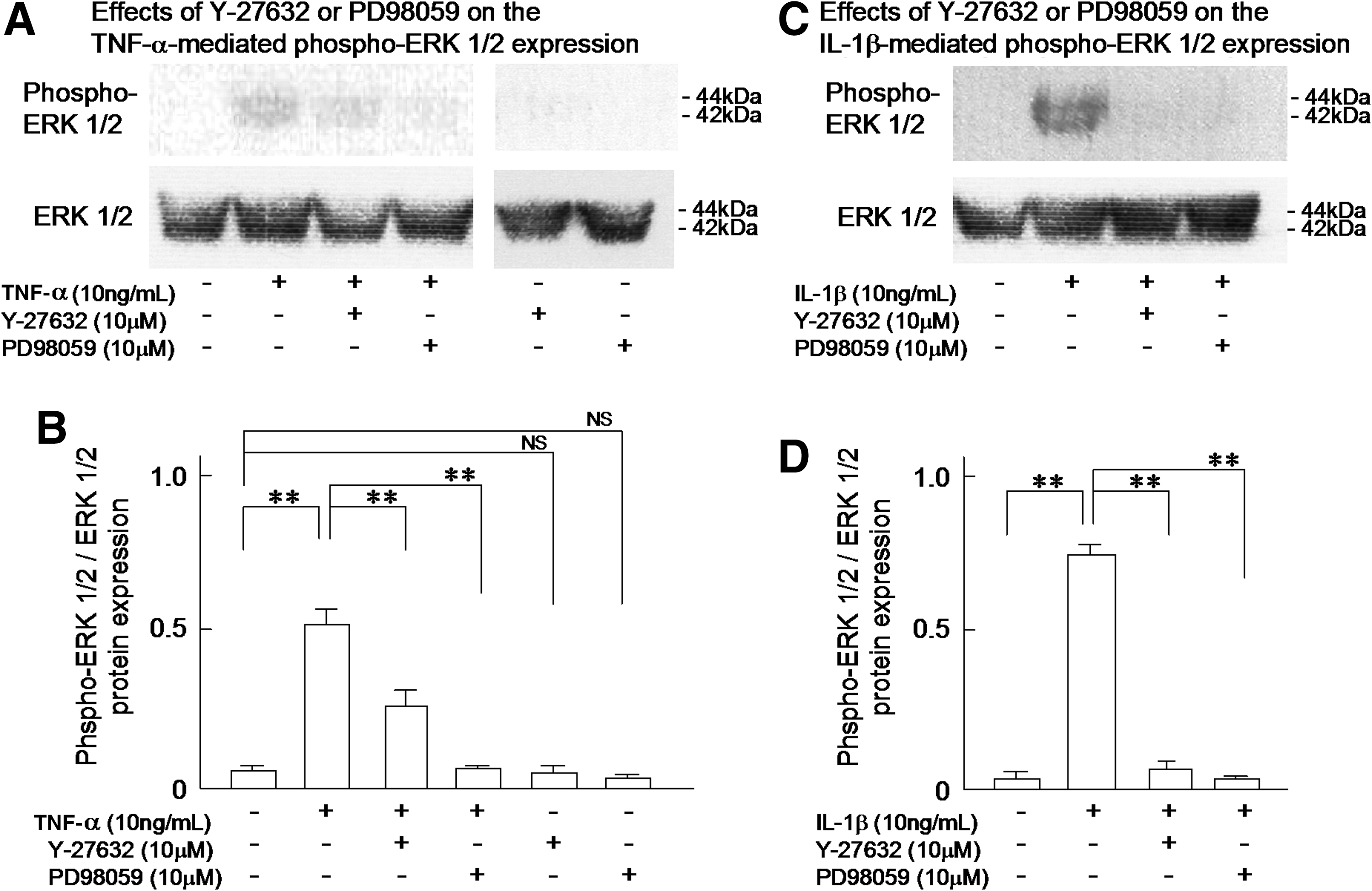
The effects of Y-27632 (10 μM) or PD98059 (10 μM) on the TNF-α (10 ng/mL) or IL-1β (10 ng/mL)-induced phosphorylation of ERK 1/2 in the cultured human LEC using Western blot analysis.
As was found for TNF-α (10 ng/mL), IL-1β (10 ng/mL) also increased the phosphorylation of ERK 1/2 in the LEC. Additional treatment with 10 μM Y-27632 or 10 μM PD98059 inhibited the completely IL-1β-induced phosphorylation of ERK 1/2 in the LEC (Fig. 7C). The findings are quantitatively summarized in Figure 7D.
Discussion
Human LEC layers play a pivotal role in protein concentration in lymph of collecting lymph vessels
First, we established human lymphatic endothelial cells from the cannulated afferent collecting lymph vessels of sentinel lymph node in breast cancer patients without chemotherapy or radiation treatment using trypsin/EDTA digestion. The cultured cells showed markedly lymphatic markers, such as transmembrane mucoprotein podoplanin 18 and transcripition factor Prox-1. 19 The hyaluronidase receptor LYVE-120 was also observed heterogenouly within each cell. In contrast, a selective marker of collecting lymph vessels, syndecan 216 was clearly found around the cultured cells. These immunocytochemical findings are compatible with the quantitative RT-PCR analysis data of the cultured cells. 16 The evidence suggests that the cultured cells seem to be selective human LEC of collecting lymph vessels. In fact, the cultured human LEC exhibited similar biological properties such as nitric oxide production and release from the LEC, to those reported in in situ collecting lymph vessels.12,21,22
In the present study, we found that significant amounts of 4 kDa dextran were able to pass through the cultured human LEC layer, whereas little or no 12 kDa or 66 kDa dextran or albumin permeated through the LEC layers. These findings were compatible with the Mayerson's classical study. 23 In experiments involving small isolated and cannulated rat collecting lymphatic vessels, we also demonstrated that while 4 kDa dextran passed through the lymphatic vessel wall easily, 70 kDa dextran was unable to pass through it. 5 The characteristics of the permeability to hydrophilic substances through collecting lymphatic vessel walls is quite similar to those obtained with the HUVEC (Fig. 2) or venular endothelial layers.8, 24 Taking the results of our present and previous studies into consideration, we have concluded that the human lymphatic endothelial cell layer plays a pivotal role in the permeability of collecting lymph vessel walls to hydrophilic substances.
Another important aspect of the present study is that 66 kDa dextran or albumin was unable to pass through the human LEC layers. We previously confirmed similar findings that the FITC-labled 70 kDa dextran did not pass through the walls of pressurized rat collecting lymphatic vessels. 5 Thus, macromolecules with molecular weights of 66 kDa or higher, such as albumin, remain in human collecting lymphatic vessels and are returned to the systemic circulation via regional lymph nodes and the thoracic duct. In conclusion, the walls of human collecting lymphatic vessels might play a crucial role in increasing the intraluminal concentrations of high molecular weight proteins. In agreement with this conclusion, it is known that the protein concentration of lymph increases as it moves from the peripheral lymph vessels to the thoracic duct.25–28 Thus, Takahashi et al. 28 also concluded that the filtration of macromolecule-free fluid across the lymphatic wall plays an important role in the lymph concentration mechanism in the collecting lymph vessels.
The detailed physiological meaning of such increases in the protein concentration of lymph remains unknown. One possibility is that the protein concentration of lymph may be an important regulator of innate immunity. For example, the excretion of nonselective T- and B-cell lymphocytes and natural killer (NK) cells into the efferent lymph vessels of lymph nodes was found to be positively correlated with the protein concentration of lymph in the afferent collecting lymph vessels.4,6,7,29,30 In agreement with the possibility, in preliminary experiments we have confirmed that the massage for lymph draining in the lower extremities of human beings produced a marked increase of the number of lymphocytes including NK cells in blood at 1–2 hours after the procedure. Further investigations using in vivo human experiments will be needed to clarify the innate immunological meaning of the condensing effect of lymph protein through the lymph vessel walls and lymph nodes.
TNF-α or IL-1β induced Rho kinase- and ERK 1/2 phosphorylation-dependent increases in the permeability of the human LEC layers to hydrophilic substances with low molecular weights
To address pathophysiological roles of the LEC layers in collecting lymph vessels in inflammation, we investigated the effects of TNF-α and IL-1β, major cytokines released from activated macrophages in the early stage of inflammation, on the permeability of human LEC layers to hydrophobic substances using fluorescent dextrans with many kinds of molecular weights.
TNF-α or IL-1β produced significant increases in the permeability of human LEC layers to 4 kDa and 12 kDa dextrans. The findings suggest that the LEC layer is one of targets of inflammatory cytokine-mediated changes to hydrophilic substances through the lymph vessel walls. In addition, treatment with the selective Rho kinase inhibitor Y-27632 or the ERK 1/2 kinase inhibitor PD98059 significantly inhibited the TNF-α- or IL-1β-induced increases in the permeability of the human LEC layers to 4 kDa and 12 kDa dextrans. In agreement with these findings, the administration of TNF-α or IL-1β produced significant changes in the distribution of cytoskeletal F-actin in the cultured human LEC, which resulted in a marked change in their shape (from a cobblestone-like shape to a spindle shape), and these morphological changes were significantly abrogated by pretreatment with Y-27632 or PD98059. In order to evaluate the mechanisms responsible for the TNF-α- or IL-1β-induced reorganization of cytoskeletal F-actin in the human LEC, we investigated the role of cytokine-mediated cell signaling (i.e., ERK 1/2 phosphorylation15,31,32) in these processes using Western blot analysis. Accordingly, we found clear evidence that TNF-α or IL-1β accelerated the phosphorylation of ERK 1/2 in human LEC. In conclusion, both TNF-α and IL-1β significantly increased the permeability of human LEC layers to 4 kDa and 12 kDa dextrans via the activation of Rho kinase and the phosphorylation of ERK 1/2, which resulted in the reorganization of cytoskeletal F-actin and related changes in the morphology of the LEC.
F-actin, a cytoskeleton protein that is found in blood vessel endothelial cells, makes an important contribution to the permeability of the blood vessel endothelium, and it has been found that inflammatory mediators increase vascular permeability by inducing the redistribution of F-actin. 33 The activation of Rho kinase in endothelial cells plays a pivotal role in the redistribution of cytoskeletal F-actin. 33 Recently, Rho kinase activity and ERK-dependent cytoskeletal tension in cells were demonstrated to promote cell growth30,34,35 and focal adhesion assembly. 36 This suggests that cell membranes, matrix stiffness, Rho kinase activity, and ERK phosphorylation are functionally linked. Therefore, in the present study, we addressed the relationships between TNF-α- or IL-1β-induced increases of the permeability of human LEC layers to hydrophilic substances and Rho kinase activity or ERK phosphorylation in human LEC. As a result, we confirmed that the TNF-α- or IL-1β-induced activation of Rho kinase and ERK phosphorylation in collecting LEC resulted in the redistribution of cytoskeletal F-actin in these cells, which produced marked changes in their shape and increased the permeability of human LEC layers to hydrophilic substances such as 4 kDa and 12 kDa dextrans. Thus, the TNF-α- or IL-1β-induced Rho kinase and ERK phosphorylation-dependent redistribution of cytoskeletal F-actin in the human LEC may be similar to the responses in blood vessel endothelial cells.
However, the validity of this assumption that the changes in endothelial cell shape is a surrogate for cytoskeletal F-actin redistribution remains unsolved. The reason why the treatment with 10 μM Y-27632 produced a partial inhibition of the TNF-α-induced, but not IL-1β, phosphorylation of ERK 1/2 also remains unknown. Further investigation will be needed in future to clarify these unsolved problems.
In conclusion, in the state of inflammation TNF-α and IL-1β released from activated macrophages accelerates the permeability of hydrophilic substances with low molecular weights less or equal to 12 kDa through the collecting lymph vessel walls and then results in the increasing of concentration of lymph protein, which may be related to pathophysiological meanings of activation of innate immunity via augmentation of nonselective release of lymphocytes from the regional lymph nodes.
Footnotes
Acknowledgments
Author Contributions: The idea to study transport regulation in human LEC layers was conceived by Y. K. and T. O., who designed the experiments and analyzed and interpreted the data; Y. K., M. K., and Y. Y. performed most of the experiments and collected the data. T. O. provided financial assistance and guidance throughout the project. Y. K. and T. O. wrote the original manuscript and approved the final version of the manuscript.
Author Disclosure Statement
The authors declare that no competing financial interests exist. The study was financially supported, in part, by Grants-in-Aid for Scientific Research (22249052, 24659098, 24590272) from the Japanese Ministry of Education, Science, Sports, and Culture and by the Intelligent Surgical Instruments Project of the METI (Japan) (2007–2012).