
Abstract
Abstract
Background:
Abundant Larval Transcript (ALT) is one of the major groups of immune-dominant proteins produced by filarial worms during their larval stage. The major B-cell and T-cell epitopic domains of the ALT-2 antigen were mapped to develop a multiple antigenic peptide (MAP) prophylactic antigen against lymphatic filariasis.
Methods and Results:
ALT MAP was constructed by solid phase peptide synthesis. The reactivity of whole ALT protein and ALT MAP against clinical sera described a high reactivity of endemic normal sera against ALT MAP compared to WbALT-2 protein. The antibody isotype pattern revealed elevated levels of IgG1 and IgG2 against ALT MAP, followed by IgG3 and IgG4. In this study we also analyzed the immune response pattern elicited by ALT MAP, ALT in mice models, which revealed similar pattern of humoral response, while low T cell proliferation in ALT MAP groups. The low proliferation could be attributed to T/B epitope arrangement on the construct, MHC restriction, and incomplete signal delivery by T cell receptor.
Conclusion:
The immunodominant epitopes in ALT MAP were found to play a crucial role in inducing high antigen specific proliferation. This revealed the significance of ALT MAP in stimulating innate immunity in offering protective immune response probably through the activation of complement cascade along with stimulation of cellular response. An improved understanding, including the construction of ALT MAP and parasite challenge study in jirds to determine the worm clearance would give a better insight in the characterization ALT MAP construct as a prophylactic vaccine candidate.
Introduction
L
Abundant Larval Transcript (ALT) is an antigen highly expressed by the infective L3 stage larvae. The ALT produced from cDNAs from the EST dataset 4 is a putative vaccine candidate conferring 76% protection in animal models. 5 Epitope regions of ALT have been analyzed, and the dominant B and T cell epitopes have been identified both by in silico and in vivo models. 6 Wuchereria bancrofti ALT-2 (Wb ALT-2) peptides have shown differential reactivity among clinical samples, which was implicated to the immunomodulatory role of the various segments of the ALT peptide (55–68, 73–91). 7
Screening of Brugia malayi ALT-2 (BmALT-2) T epitopes also reiterated the finding that regions 55–68, 73–91 carried the immunomodulatory domains of ALT protein, which was shown by the elicitation of high levels of IL-10 in animal models. 6
Conventional ways of developing vaccines against infections, either on pragmatic grounds or by identifying protective antigens and attempting to mimic natural immune responses, have largely been unsuccessful for parasitic infections, mainly because of the complexity of the immunological processes involved. Hence in this study, we constructed a multiple antigenic peptide (MAP) by arranging regions of identified immunodominant epitopes by solid phase peptide synthesis and analyzed their reactivity pattern with human clinical samples, along with the cellular response to filarial antigens. Construction of ALT MAP proved to exceed the immune response elicited by whole protein ALT, as they carry only the dominant epitope regions of ALT excluding the immunomodulatory regions of the same. Comparative study was performed to analyze the immune response pattern elicited by ALT MAP and ALT in mice models.
Materials and Methods
ALT MAP synthesis
The immunodominant domains of WbALT-2 comprising of both B and T epitopes were incorporated in the construction of ALT MAP.6,7 The MAP construct consisted of three ALT epitopic domains (AE): AE1 (2–21), AE2 (21–50), AE3 (104–128) (Table 1). The synthesis was done using standard Fmoc chemistry. 8
Briefly, the Fmoc-Gly-HMP-TentaGel resin, substitution factor 0.16 mmole/g, was pre-swelled in dichloromethane (DCM) for 10 min as per the manufacturer's instructions and left for swelling in dimethylformamide (DMF) overnight. The F-moc group was removed using 20% piperidine in DMF. To this, F-moc-Lys-(ivDde) was attached. The N-terminal F-moc was removed and the peptide was synthesized. At the end of the sequence, the last amino acid used was with a different protecting group, t-Boc, in order to prevent its removal while deprotecting F-moc groups during attachment of different sequences and to prevent unwanted chain elongation thus end capping the synthesized peptide sequences. In the next step, the ξ-NH2 protecting group, ivDe was removed using 2% hydrazine in DMF and another Fmoc-Lys-(ivDde) was attached. The cycle was repeated till the completion of all the sequences. ALT MAP was purified by high performance liquid chromatography.
Recombinant WbALT-2 production
WbALT-2 gene in T7 expression system (pRSETB) was transformed in BL21pLysS E.coli strain for expression studies. It was induced at 0.6 OD with 1 mM isopropyl β-D-thiogalactoside for 4 h at 37°C. The expressed recombinant protein WbALT-2 was analyzed on 12% SDS PAGE and purified by electro elution of gel using reversible KCl staining. 9
Study population
Blood samples were collected from different clinical groups of lymphatic filariasis patients residing in the endemic regions (Chennai and Vellore districts of Tamil Nadu, India). The samples were collected after obtaining the individual's consent, as per the guidelines of the human ethical committee of Anna University, Chennai. Sera were grouped into endemic normal (EN), microfilariae positive (MF), chronic pathology (CP) based on the parasitological examination, and clinical signs and symptoms. Non-infected individuals living in non-endemic regions were used as control (non-endemic normals: NEN). Thus a total of 21 individuals from each clinical sera group and 10 NEN individuals were sampled.
Immunoreactivity with clinical sera and isotype determination
Human clinical sera were analyzed for their reactivity against both ALT MAP and WbALT in natural infection by performing ELISA. Briefly, ELISA plates were coated with ALT (100 ng) or ALT MAP (100 ng), left overnight at 4°C, further incubated with 1:100 dilution of human serum from different clinical populations, namely EN, MF, CP, and NEN. Plates were incubated at 37°C for 1 h, following which they were washed and incubated with goat anti-human IgG-ALP conjugate for 1 h at 37°C and then washed again. Plates were developed with substrate containing p-nitrophenyl phosphate (PNPP) (1 mg/mL) in substrate buffer (100 mM Tris-Cl, pH 9.5, 100 mM NaCl, 5 mM MgCl2) and absorbance was measured at 405 nm. The mean absorbance of the negative control NEN sera+3SD was taken as the cutoff value.
The protocol for IgG isotype ELISA was similar to that described above. Briefly, ALT /ALT MAP antigens were coated (500 ng) in ELISA plate and incubated overnight. After washing, sera samples (1:50 dilution) were incubated at 37°C for 1 h at 4°C. Monoclonal antibodies against human IgG1, IgG2, IgG3, and IgG4 (Sigma Aldrich) were incubated at 37°C for 1 h. Following incubation, ALP-labeled goat antimouse IgG was added as a secondary antibody and PNPP substrate was sequentially added to detect the concentration of the respective isotype of antibodies in the sera.
PBMC proliferation assay
PBMC were obtained from heparinized blood purified by sedimentation on a Ficoll gradient from endemic normal individuals (n=13). The cells were washed and the viability was determined by Trypan blue dye exclusion method. PBMCs were placed in flat-bottomed wells of microtiter plates at a concentration of 2×106 cells/mL in RPMI-1640 supplemented with gentamicin (80 μg/mL), 25 mM HEPES, 2 mM glutamine, and 10% fetal bovine serum. The cells were stimulated with ALT protein (1–20 μg/mL), ALT MAP (10–50 μg/mL), Con A (10 μg/mL) as positive control. Wells with medium alone were used as unstimulated controls. The cultures were pulsed with [H3] thymidine (USB, Amersham Pharmacia, UK) at 0.5 μCi/well. The cells were incubated for 18 h and harvested onto glass fiber disks; thymidine incorporation was measured by a liquid scintillation counter. Proliferative responses were expressed as stimulation index (S.I.) calculated as counts per minute (cpm) of cells stimulated with antigen divided by cpm of unstimulated cells.
Immunization of mice with ALT MAP, ALT
Six–eight-week-old female Balb/c mice were procured from Kings Institute, Chennai, and housed in animal facility at Anna University, Chennai. A group of six mice were immunized with 50 μg of ALT MAP or 25 μg of ALT in 100 μL of PBS and mixed with alum at 1:1 ratio. 10 For vaccination of the animal groups, four doses of the corresponding antigens were injected at weekly intervals.
Splenocyte proliferation assays
The ALT MAP and ALT protein immunized mice were sacrificed on day 42 and the spleens were removed aseptically. Splenocytes were separated and washed twice with fresh culture medium (RPMI 1640). Lysis buffer (0.1% ammonium chloride) was added to the pellet to remove the RBCs and the cells were counted. The single cell suspension was cultured in triplicate in 96-well plates at 2×105 cells/mL in RPMI 1640 medium (100 μL/well) (Sigma, St. Louis, USA) supplemented with gentamycin (80 μg/mL) (Ranbaxy Laboratories, India), 25 mM HEPES (USB, Amersham Pharmacia, UK), 2mM glutamine (USB, Amersham Pharmacia), and 10% fetal bovine serum. The cells were then stimulated in vitro with ALT antigen (0.1, 1, 5, and 10μg/well), ALT MAP (1, 5, 10, 25 μg/well), or positive control Con A (1 μg/well). 10 Wells with medium alone were used as unstimulated controls. The cultures were pulsed with [H3] thymidine (USB, Amersham Pharmacia) at 0.5 μCi/well. The cells were incubated for 18 h and harvested onto glass fiber disks; thymidine incorporation was measured by a liquid scintillation counter. Proliferative responses were expressed as stimulation index (S.I.) calculated as counts per minute (cpm) of cells stimulated with antigen divided by cpm of unstimulated cells. All cultures were taken in triplicates and the results expressed as mean S.I.±SEM.
Statistical analysis
All statistical analysis was done using graph pad prism software version 5. For clinical sera reactivity and T cell proliferation studies, two-tailed T test was performed.
Results and Discussion
Clinically uninfected individuals residing in endemic regions (Endemic Normal) exhibit acquired protective immunity (resistance against filarial parasites), which paved the way in identifying the novel protective antigens in lymphatic filariasis. 11 Chronic pathology patients exhibit high immunopathology, for which care should be taken that the identification of new vaccine targets should not be expressed in adult stage, which otherwise would exacerbate the disease due to vaccine-induced immune response. 12 Many potential immune evasion genes of filarial nematodes have been identified, which encompasses Bm-CPI-1 and -2 (cysteine protease inhibitor), Bm-SPN-2 (microfilarial serpin), Bm-MIF-1 and -2 (migration inhibitory factor), Bm-TGH-1 and -2 (transforming growth factor homologue), Bm-ALT-1 and -2 and Bm-VAL-1 (venom allergen Ancylostoma secreted protein-like family). 13 Among these genes, the ALT-2 gene has high efficacy in clearing filarial parasites in an animal model. Since ALT genes share no homology with host, the high larval specific expression of this gene4,14 indicates that heightened reactivity to ALT is unlikely to exacerbate pathogenic responses to adult worms that might initiate lymphatic damage. Many homologues of ALT have been identified.
Clinical sera reactivity
In this study we constructed ALT MAP with immunodominant epitopes excluding the immunomodulatory domains of ALT to ensure if these putative epitopes were able to mount adequate protective immune response to EN in order to develop a prophylactic vaccine candidate. The immunoreactivity with clinical sera (Fig. 1) showed a significant (p=0.002) number of EN populations to be highly reactive against ALT MAP with optical density (O.D) ranging from 0.312–1.444, compared to ALT (O.D: 0.230–1.128). The higher reactivity of ALT MAP against EN sera confirms the significance of the immunodominant epitopes of ALT and the prospects of eliciting a protective immune response. The MF sera reactivity pattern showed significantly high reactivity against WbALT (p=0.003) in comparison to ALT MAP, which showed many samples lying below the cut off value. These results may indicate that the immunomodulatory domains of WbALT may have paved way in higher reactivity of the MF sera, while the exclusion of these domains in ALT MAP have elicited a lower reactivity against MF and higher reactivity against EN sera. The overall reactivity pattern of high total IgG elicited by EN sera led us to speculate the isotype levels of IgG elicited by both the antigens against EN sera.
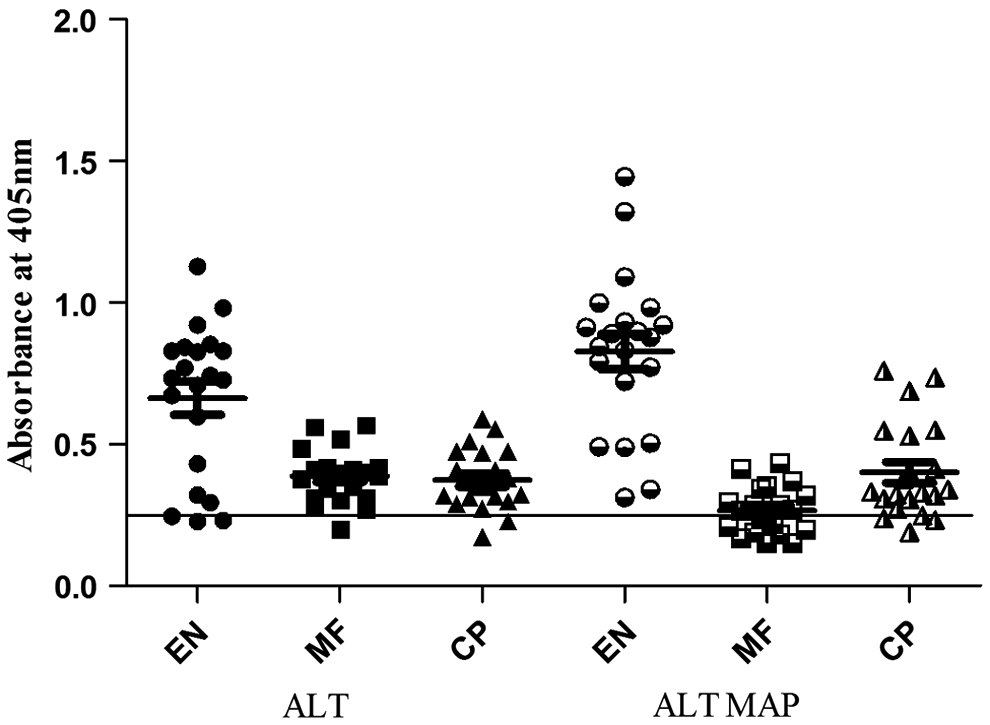
Reactivity of ALT, ALT MAP with human sera samples from different clinical groups: endemic normals (EN), microfilaraemics (MF), and chronic pathology (CP) (n=21). Non-endemic normal (NEN) sera was used as negative control (n=10). Data are represented as scatter plot where each dot represents absorbance of individual sera. The line drawn parallel to the X-axis represents the cut off value (mean O.D. of NEN±3 SD).
IgG isotype profile
The isotype pattern revealed high levels of IgG1 and IgG2 against ALT MAP, followed by IgG3 and IgG4 (p<0.05) (Fig. 2). However, WbALT elicited low levels of IgG2 and IgG3. The major immunoglobulins involved in antifilarial immunity are IgG, IgM, and IgE.15–17 Among this IgG isotype, profiles can be measured in all categories of patient groups and the IgG isotypes vary with different clinical groups of filariasis that can be related to the severity of infection. 18 IgG1 and IgG3 are strong activators of complement C1, whereas IgG2 are weak activators and IgG4 does not activate complement.19–21 The high levels of IgG1, followed by IgG2 and IgG3, elicited by ALT MAP may hypothesize the role of innate immunity in offering protective immune response through activation of complement cascade. Though ALT elicited a similar pattern of isotype profile, the optical density (OD) values were significantly lower than ALT MAP, along with low profiles of ALT specific IgG2, IgG3, and IgG4.
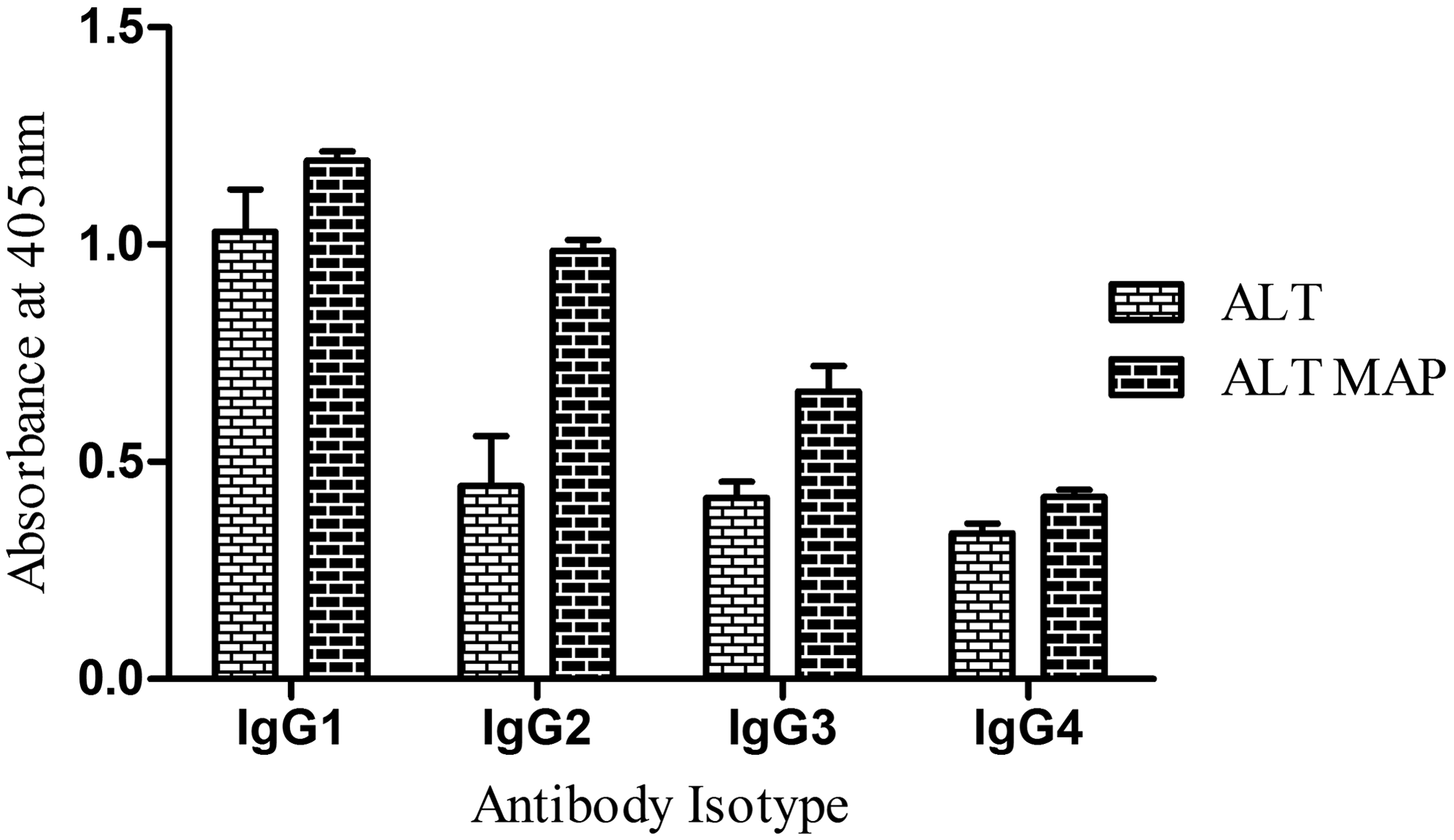
Antibody isotype profile of ALT, ALT MAP in endemic normal population. Data represents mean absorbance of ten samples±SD.
Cellular immune response
To analyze the ALT MAP specific T cell response, human PBMCs were separated from endemic normal individuals and stimulated in vitro with WbALT and ALT MAP. The PBMCs showed significantly high proliferation (p<0.002) when stimulated with ALT MAP compared to WbALT (Fig. 3). The elevated levels of ALT MAP specific proliferation can be attributed to the dominant T cell epitopes used in the ALT MAP construct. The immunomodulatory regions in ALT play a crucial role in diminishing the ALT specific proliferation.
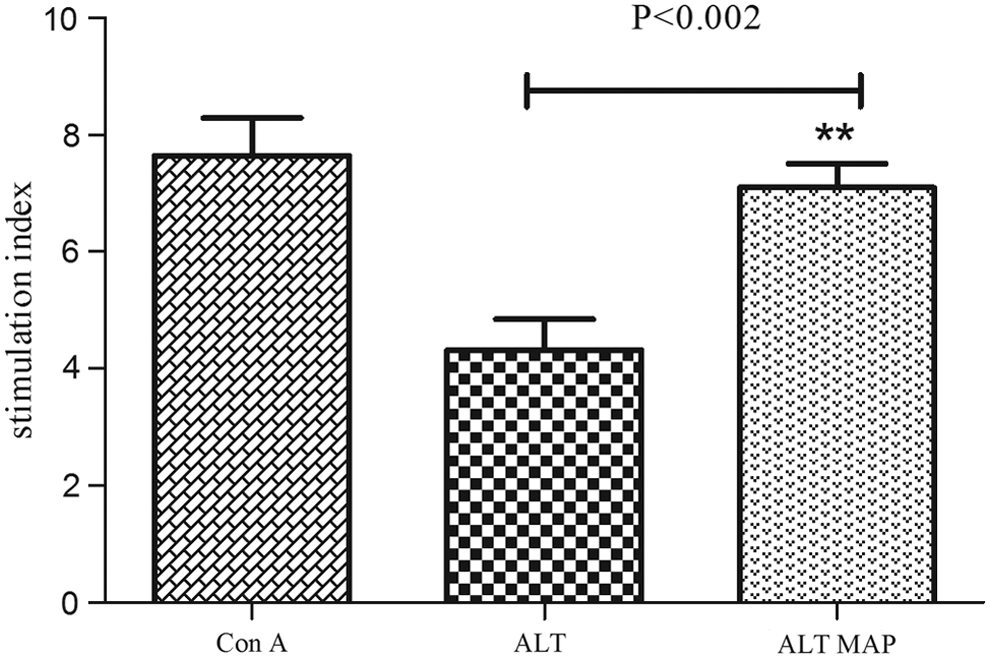
Proliferation of human PBMCs stimulated with ALT, ALT MAP. Con A was used as a positive control. The data is represented as mean stimulation index (S.I.) of 13 endemic normal individuals±SEM.
Antibody titer and isotype analysis
The protein and MAP immunized groups induced high titers of antibody levels in Balb/c mice. The mean peak titer of 32,000 was induced on day 21 by ALT, while the same titer was induced by ALT MAP on day 28 (Fig. 4). The antibody isotype elicited by ALT MAP, ALT showed similar patterns of antibody isotype profile (Fig. 5). High levels of IgG2b and IgG2a was seen in ALT MAP groups. However, ALT groups elicited comparable levels of IgG2b and IgM that paralleled with ALT MAP. The IgG1 level was also found to be high in ALT MAP group compared to ALT immunized group, which is a marker of Th2 immune response. The type 2 response induced by infective L3 stage and adult parasites are known to be protective against helminth infection. 22 Both IgG1 and IgM23,24 have been implicated in protection in experimental filariasis.
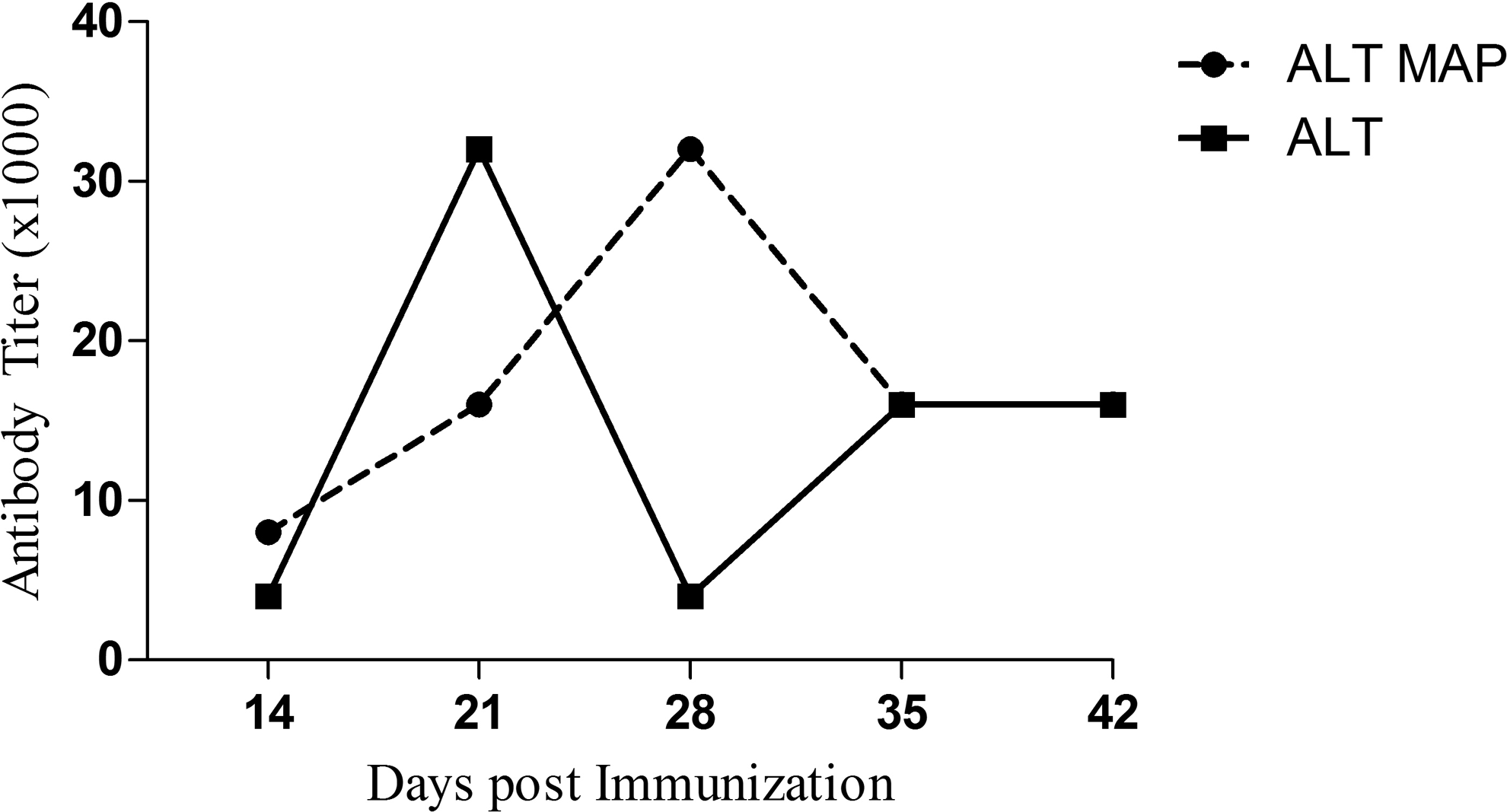
ALT and ALT MAP specific antibody titers (in thousands) in mice at different intervals post-immunization.
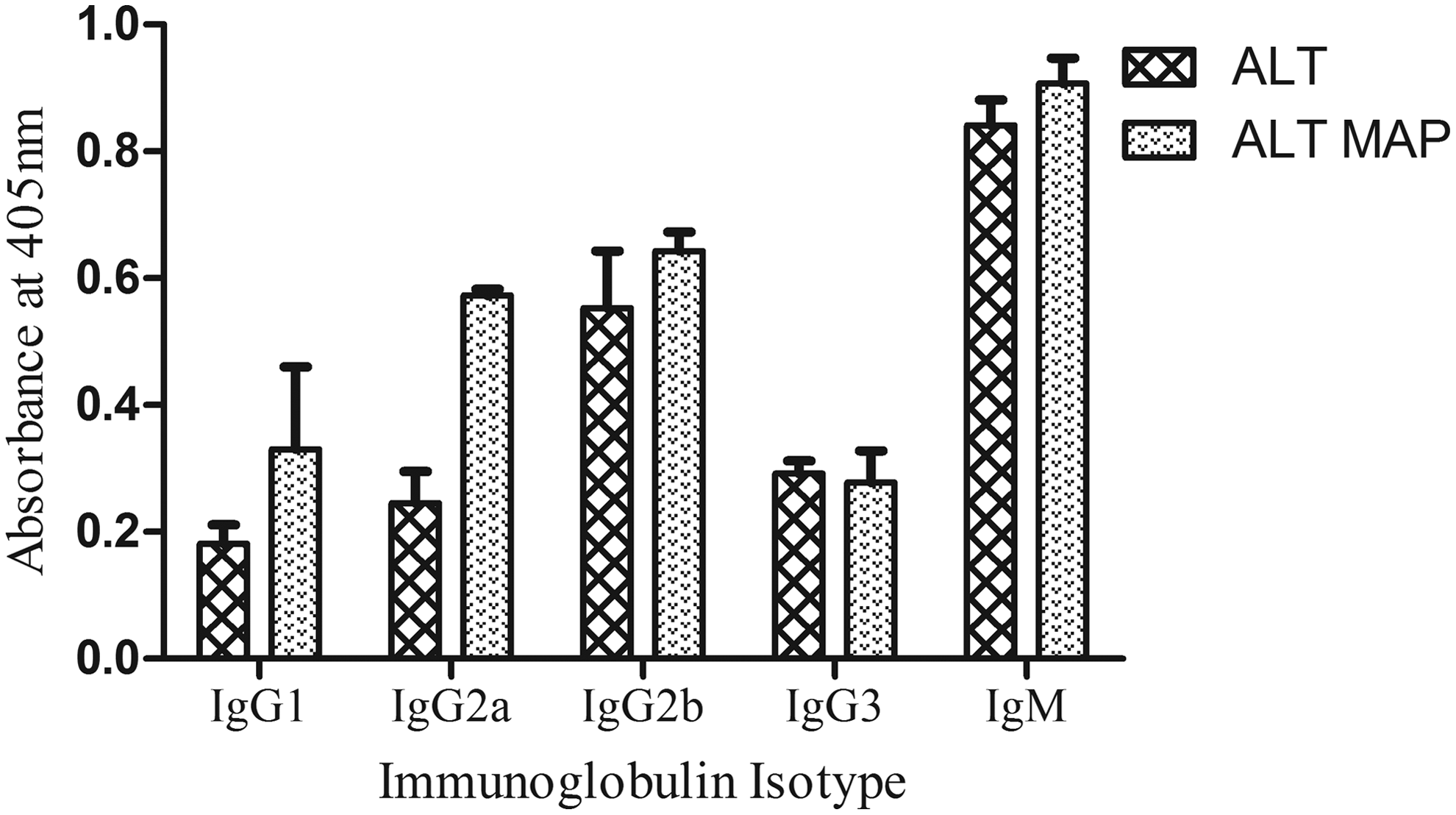
The antibody isotypes measured in 1:100 diluted sera of ALT, ALT MAP are given as bars. Each bar represents mean absorbance of 6 mice±SD.
T cell proliferation
The stimulation index indicating the lymphocyte proliferation of effector T cells in ALT MAP groups shows a significantly low proliferation to both antigens ALT (p=0.01) and ALT MAP (p=0.009) compared to ALT group (Fig. 6). Though the humoral response elicited by the ALT MAP group was similar to that of ALT, this was not observed in the cellular response. This could be due to the arrangement of B/T epitopes in ALT MAP construct, which also could pave way for this diminished cellular response. Studies have reported that the peptides that consist of two tandemly repeated epitopes joined by a flexible linker have an increased affinity for class II molecules and are more potent at inducing proliferation of T cell clones than monomeric epitopes. 25
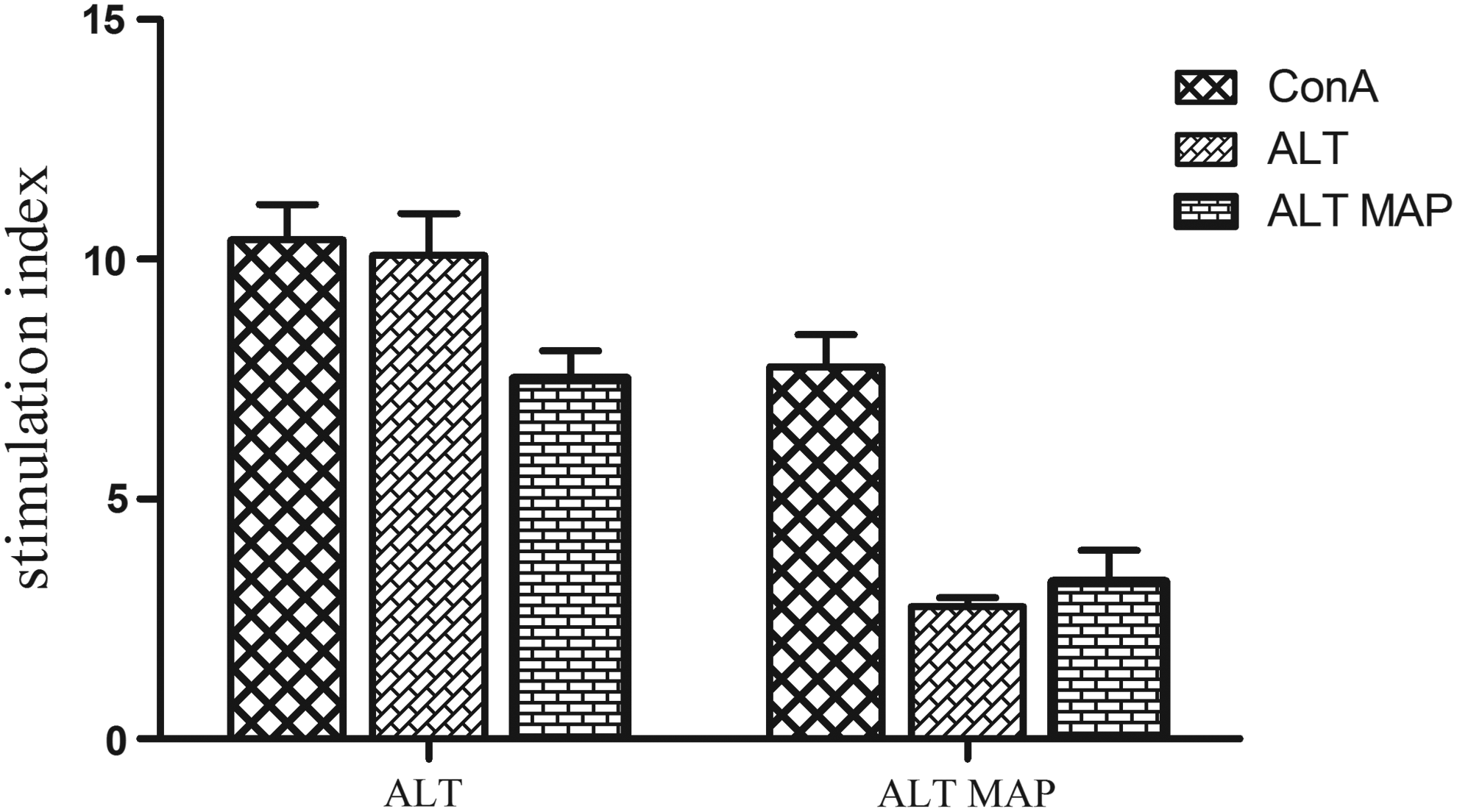
Splenocyte proliferation of ALT and ALT MAP immunized mice stimulated with ConA ALT and ALT MAP. The data are represented as mean stimulation index (S.I.) of six mice±SEM.
The low proliferation elicited by ALT MAP could be due to any of the following reasons : (1) blockade of MHC presentation by competitor peptides restricted to same class II molecule, 26 (2) TCR antagonism or inhibition of T cell activation, 27 (3) partial TCR antagonism which leads to altered or incomplete signal delivery. 28 The C-terminal region (104–128) of ALT has been cited as a dominant T epitope which induced a Th2 response, and the N-terminal region (2–21) has shown induction of IFN-γ that enhanced cellular response. 6 Since the ALT MAP construct was designed to carry both T/B immunodominant epitope regions of ALT, the low proliferation could be attributed to the arrangement of the epitope regions.
Conclusion
The human filarial infection is characterized by a profound bias in the Th cell response of infected individuals, with a dominating Th2 response and impaired parasite specific Th1 response.29–31 For a prophylactic vaccine to be effective, a balanced Th1/Th2 response is required. Multiple epitopes covering regions of immunological hot spots when delivered simultaneously as a single chimeric construct are known for induction of a broad spectrum of immune responses. 32 Thus put together, ALT MAP construct excelled ALT in human clinical samples, since it induced protective immune response with the mixed B and T cell epitopes of ALT protein. The immune response elicited by ALT could be due to the presence of immunomodulatory regions that are absent in ALT MAP, making it an effective peptide vaccine, the first of its kind against lymphatic filariasis. However, the ALT MAP construct elicited protective humoral response in mice models similar to ALT, while the low proliferation of T cells could be attributed to the arrangement of T epitopes on the construct. This can be addressed by including universal T epitope in the construct as an additional epitope or tandemly arranging T epitopes. Further insight is required in concluding the immune response pattern of ALT MAP, which includes the parasite challenge studies in Jirds to know the parasite clearance ability of this construct.
Footnotes
Acknowledgment
This work was supported by Department of Biotechnology (DBT) and University Grants Commission (UGC), Gov't of India. The senior research fellowship to Aparnaa Ramanathan from CSIR, Gov't of India is gratefully acknowledged.
Author Disclosure Statement
No competing financial interests exist.