
Abstract
Abstract
Background:
The lymphatic system contributes to fluid homeostasis in various tissues. Recent evidence suggests that lymphangiogenesis induced by a high-salt diet (HSD) is associated with blood pressure regulation. Lymph nodes, located along lymphatic pathways, are not only important secondary lymphoid tissues for cancer metastasis, inflammation, and immune responses, but are also important for fluid homeostasis. Afferent lymphatics collect lymph from the pre-nodal area and efferent lymphatics drain lymph out of the lymph nodes. However, the difference in mechanical activity between afferent and efferent lymphatics and the effect of a HSD on these vessels have not been shown.
Methods and Results:
Changes in mechanical activity of isolated afferent and efferent lymphatics in normal salt diet (NSD) and 4-week HSD mice in response to increases in intraluminal pressures from 3 to 7 cmH2O were measured using video-microscopy. The higher intramural pressure equivalently decreased pumping activity of afferent and efferent lymphatics in NSD mice. A HSD suppressed the amplitude, ejection fraction, and stroke volume of afferent lymphatics, leading to marked reductions in pumping activity. In contrast, the pumping activities of efferent lymphatics were resistant to a HSD and were preserved by enhancing the contraction frequency.
Conclusions:
A HSD differentially modulated the mechanical activity of afferent and efferent collecting lymphatics in murine iliac lymph nodes.
Introduction
T
Recently, specific adhesion molecules involved in the interaction between HEV and lymphocytes have been detected. 4 Cells floating in interstitial spaces and lymphatic capillaries, known as the initial sites for cancer metastasis, are reported to interact through cytokines or chemokines. 5 Furthermore, clinical examination of sentinel lymph nodes is advantageous for cancer patients to improve quality of life by preventing secondary lymph edema and reducing surgical trauma.6–8 Thus, in the pathophysiology of the lymphatic system, the lymph node is a critical factor for lymph dynamics.
Lymph nodes have a unique structure involving blood and lymph circulation. In general, several afferent lymphatic vessels connect to lymph nodes and one or more relatively large efferent lymphatic vessel drains lymph from the nodes. These afferent and efferent lymph vessels that penetrate lymph nodes work as a series of lymphatic pumps to propel the intra-nodal lymph fluid by oscillatory constriction and dilation, as well as draining lymph from more peripheral parts. Smooth muscle cells in the lymph vessels of humans and animals exhibit intrinsic spontaneous activity, and serve as the main machinery of the pump in vivo and ex vivo. 1 Intraluminal pressure of lymphatics is one of the most important factors for regulating spontaneous contraction of lymphatic smooth muscle. An elevation of intraluminal pressure increases the frequency and decreases the amplitude of oscillatory changes in the diameter of lymphatic vessels.9–11 Despite extensive research on the lymphatic system, functional analyses of afferent and efferent lymphatics have not been fully shown.
Hypertension is a major risk factor for diseases such as stroke, heart failure, and kidney disease. High salt intake is closely associated with the pathogenesis and development of salt-sensitive hypertension. 12 Salt loading, via inefficient renal excretion of sodium, often results in the expansion of circulating body fluid volumes. As an effect of salt loading on the lymphatic system, Machnik et al. reported vascular endothelial growth factor (VEGF)-C mediates hyperplasia of lymph capillaries in the rodent ear and proposed that capillaries aid in maintaining fluid homeostasis and blood pressure. 13 However, salt-induced functional changes of the lymph transport system remain uncharacterized in collecting lymphatics, especially in vessels perfusing through lymph nodes. Therefore, in the present study we examined the effect of intraluminal pressure on the mechanical activity of afferent and efferent lymphatics in the lymph nodes following a normal salt diet (NSD) and a high-salt diet (HSD).
Materials and Methods
Animals
Male ICR mice, 5 weeks of age (n=34, Tokyo Jikken Dobutsu, Tokyo, Japan), were divided into NSD (n=17) and HSD (n=17) groups. NSD mice were fed a standard pellet diet and water for 9 weeks. HSD mice were fed a NSD for 5 weeks and then received a HSD (8% NaCl; w/v) and 1% (w/v) saline for 4 weeks. The Animal Ethics Committee of The University of Tokyo School of Medicine approved all experimental protocols in accordance with the principles and guidelines on animal care of the Physiological Society of Japan.
In vivo studies: Blood pressure and heart rate measurements
Arterial blood pressure and the heart rate of NSD (n=5) and HSD (n=5) mice were measured using the tail-cuff method (BP-98A-L, Natsume).
Ex vivo studies: Tissue preparations
Mice were sacrificed by a forceful impact to the head, followed by decapitation. After an incision to the abdominal cavity, afferent and efferent lymphatics with the iliac lymph nodes were excised and placed in a Petri dish containing cold (4°C) Krebs-bicarbonate solution (in mM: 120.0 NaCl, 5.9 KCl, 2.5 CaCl2, 1.2 MgSO4, 1.2 NaH2PO4, 5 glucose, and 25.0 NaHCO3).14,15
With microsurgical instruments and an operating microscope, lymphatic vessels were isolated and then transferred to a 10 mL organ chamber with two glass micropipettes containing Krebs-bicarbonate solution. After each lymph vessel was mounted on a pipette (proximal) and secured with sutures, the perfusion pressure was raised to 4 cmH2O to flush out and clear the vessel. Subsequently, the distal end of the vessel was mounted on the outflow micropipette (distal). The proximal and distal micropipettes were connected via Tygon tubing with a 10 mL syringe and a stopcock, respectively. Krebs-bicarbonate solution bubbled with 5% CO2/10% O2/85% N2 (v/v) was perfused extraluminally over the lymphatics within the organ chamber. The flow rate of the superfused solution was kept at 2.5 mL/min throughout the experiment. After cannulation of the lymphatics, the chamber was transferred to the stage of a microscope (Olympus BH-2, Tokyo, Japan). The lymphatics were then warmed slowly to 37°C and allowed to equilibrate for ∼90 min.
Measurement of mechanical characteristics of lymphatic vessels
To measure the mechanical activity of isolated lymphatic vessels we used a video-microscopy system, as previously described.14,15 An objective lens (x10) and a monochrome charge-coupled device camera (KOCOM, KCB-270A, Korea) were used to obtain images of the lymphatics, which were displayed on a monochrome television monitor (Hamamatsu Photonics, C1846, Hamamatsu, Japan). The diameter of the lymphatics was measured with a diameter-detection device with an edge-detection method. 16 They were recorded on a DVD recorder (Pioneer, DVR55, Tokyo, Japan) and a direct-writing oscillograph (Sanei-Sokki, Recti 8K, Tokyo, Japan). Length of isolated afferent and efferent specimens after cannulation were 2–3 mm and 4–5 mm, respectively. The vessels used in the present study contained one valve and we measured its diameter at an upstream of the valve. 14,15
In the present study, we measured the passive diameter (PD; μm), active diameters (maximum diameter; Dmax μm and minimum diameter; Dmin μm), and frequency (min−1) of lymphatic activity in NSD and HSD mice. We calculated the amplitude (Dmax−Dmin), ejection fraction (EF; (πDmax2−πDmin2)/πDmax2), stroke volume index (SVI; πDmax2−πDmin2), and frequency times SVI. 16
Effect of intraluminal pressure on lymphatic function in NSD and HSD vessels
After the equilibration period at an intraluminal pressure of 5 cmH2O, intraluminal pressure was increased from 3 to 7 cmH2O in 2 cmH2O steps by elevation of a 10 mL syringe connected to the inflow tubing, while the outflow tubing was closed with a stopcock throughout the experiment. Each level of pressure was maintained for ∼15 min to allow the vessels to exhibit stable and spontaneous diameter oscillations. At the end of each experiment, Krebs-bicarbonate solution was changed to a Ca2+-free Krebs-bicarbonate solution that also contained ethylenediaminetetraacetic acid (EDTA; 1 mM) and nifedipine (a blocker of Ca2+ channels, 10 μM). Lymphatic vessels were incubated with Ca2+-free solution for ∼20 min and the pressure steps were repeated. PDs were measured at each pressure value.9–11
Drugs
All salts and EDTA were obtained from Wako (Osaka, Japan); nifedipine was from Sigma Aldrich (St. Louis, MO, USA).
Statistics
Changes in the diameters of vessels following increases in intraluminal pressure were expressed as a percentage of the corresponding PD (100×diameter in the presence of extracellular Ca2+/diameter in the absence of extracellular Ca2+).9–11 Data are presented as the mean±standard error of mean, and n indicates the number of vessels. Significant differences (p<0.05) were determined using the unpaired Student's t-test, as appropriate.
Results
Effects of a HSD on blood pressure and heart rate
Systolic arterial pressure of HSD mice (110±3 mmHg, n=5) at 9 weeks was significantly higher than that of NSD (102±1 mmHg, n=5). There was no significant difference in heart rate between NSD (662±44 beats/min) and HSD (675±44 beats/min, n=5) mice.
Passive and active diameters of afferent and efferent lymphatics in NSD and HSD mice: Responses to elevated intraluminal pressure
To examine the effect of salt loading on physiological parameters of lymphatic vessels, we first measured the diameters (PD, Dmax, and Dmin) of afferent and efferent lymphatic vessels in NSD and HSD mice in response to increases in intraluminal pressure.
In ICR mice, PD, Dmax, and Dmin of afferent lymphatic vessels were consistently smaller than those of efferent lymphatics at the corresponding intraluminal pressure (Fig. 1A–F). The three parameters (PD, Dmax, and Dmin) of both afferent and efferent lymphatic vessels increased in response to elevated intraluminal pressure from 3 to 7 cmH2O with or without salt loading (Fig. 1A–F). A HSD significantly decreased PD, Dmax, and Dmin of afferent lymphatic vessels, but not those of efferent lymphatic vessels over the range of intraluminal pressures we tested (Fig. 1A–C).
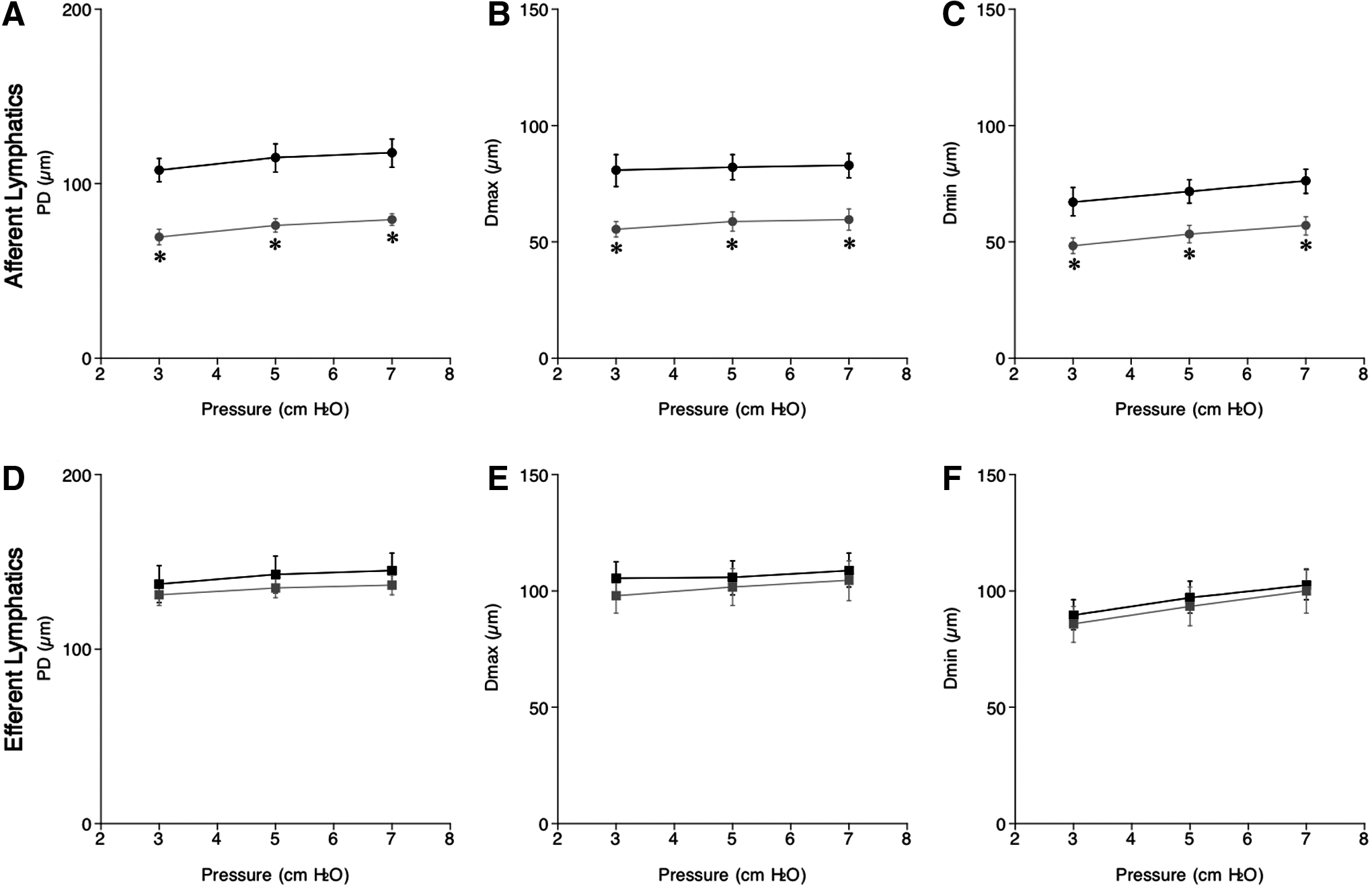
Changes in passive diameter (PD; μm,
Percent diameters of afferent and efferent lymphatic vessels in NSD and HSD mice: Responses to elevation in intraluminal pressure
Using measured values of PD, Dmax, and Dmin, we then calculated % Dmax and % Dmin, standard parameters for myogenic tone and normalized contractility in ex vivo vessels, respectively, using the following formula: 100×diameter in Ca2+ Krebs-bicarbonate solution/diameter in Ca2+-free Krebs-bicarbonate solution.9,10
In response to increases in intraluminal pressures from 3 to 7 cmH2O, % Dmax of afferent lymph vessels in NSD (from 75±2% to 71±2%, n=6) and HSD (from 80±4% to 75±4%, n=6) mice only slightly decreased, showing a negative relationship between pressure and % Dmax in NSD (−1.0±0.5, n=6) and HSD (−1.3±1, n=6) mice. There were no significant differences in % Dmax and the slopes between NSD and HSD mice (Fig. 2A). Thus, a HSD did not affect the myogenic tone of afferent lymphatics that was slightly increased by the higher pressure. The % Dmin of afferent lymph vessels in NSD and HSD mice was constant under intraluminal pressures from 3 to 7 cmH2O. The % Dmin of afferent lymphatics in HSD (70±3%, n=6) mice at 5 cmH2O was significantly larger than that of NSD (63±2%, n=6) mice, suggesting decreased contractility following a HSD (Fig. 2B). The relationship between pressure and % Dmin were not significantly different between NSD (0.7±0.4, n=6) and HSD (−0.5±0.7, n=6) mice.
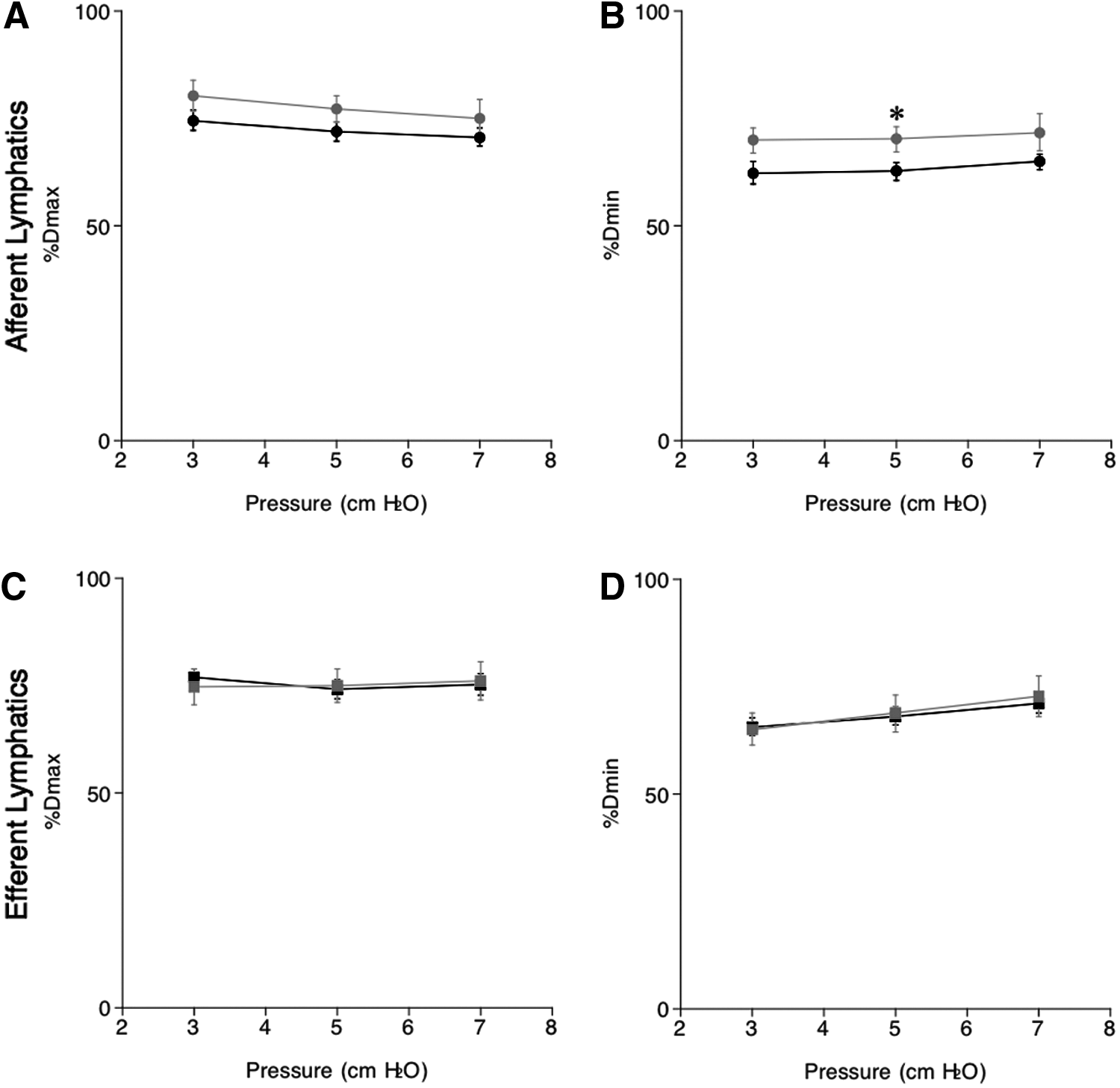
Intraluminal pressure-induced changes in % maximum diameter (% Dmax,
The % Dmax of efferent lymph vessels in NSD and HSD mice was constant under intraluminal pressures from 3 to 7 cmH2O without significant differences between NSD and HSD mice (Fig. 2C). Elevation of pressure increased % Dmin of efferent lymphatics in NSD and HSD mice without significant differences between NSD and HSD mice at pressures from 3 to 7 cmH2O. A positive relationship between pressure and % Dmin was observed in NSD (1.4±0.1, n=6) and HSD (1.9±0.6, n=6) mice, but there was no significant difference between the groups (Fig. 2D). Thus, myogenic tone of afferent and efferent lymph vessels was almost identical, remained constant in response to increments in intramural pressure, and was unaffected by salt loading. Contractility of both afferent and efferent lymph vessels decreased similarly in response to the increment in intramural pressure. However, contractility of only afferent lymph vessels was decreased by a HSD at a pressure of 5 cmH2O.
Inotropism and chronotropism of afferent and efferent lymphatic vessels in NSD and HSD mice: Responses to elevated intraluminal pressure
To further clarify the effects of a HSD on inotropism and chronotropism of pumping activity of inlets and outlets in the iliac lymph nodes, we calculated the amplitude, EF, frequency, SVI, and frequency×SVI of lymphatic vessels from NSD and HSD mice in response to increases in intraluminal pressure.
Elevation of intraluminal pressure from 3 to 7 cmH2O reduced amplitude and EF, inotropic parameters of pumping activity, and of afferent and efferent lymphatics, indicating that increments in intraluminal pressure caused a negative inotropic effect on spontaneous contraction of afferent and efferent lymphatics both in NSD and HSD mice (Fig. 3A–D). Although the amplitude and EF of afferent and efferent lymphatics following a NSD were similar, a HSD significantly decreased the inotropic parameters in afferent lymph vessels (Fig. 3A, B), but not in efferent lymph vessels (Fig. 3C, D), indicating that high salt loading selectively inhibits inotropic pump activity in afferent lymph vessels of the lymph nodes not affecting the efferent parts.
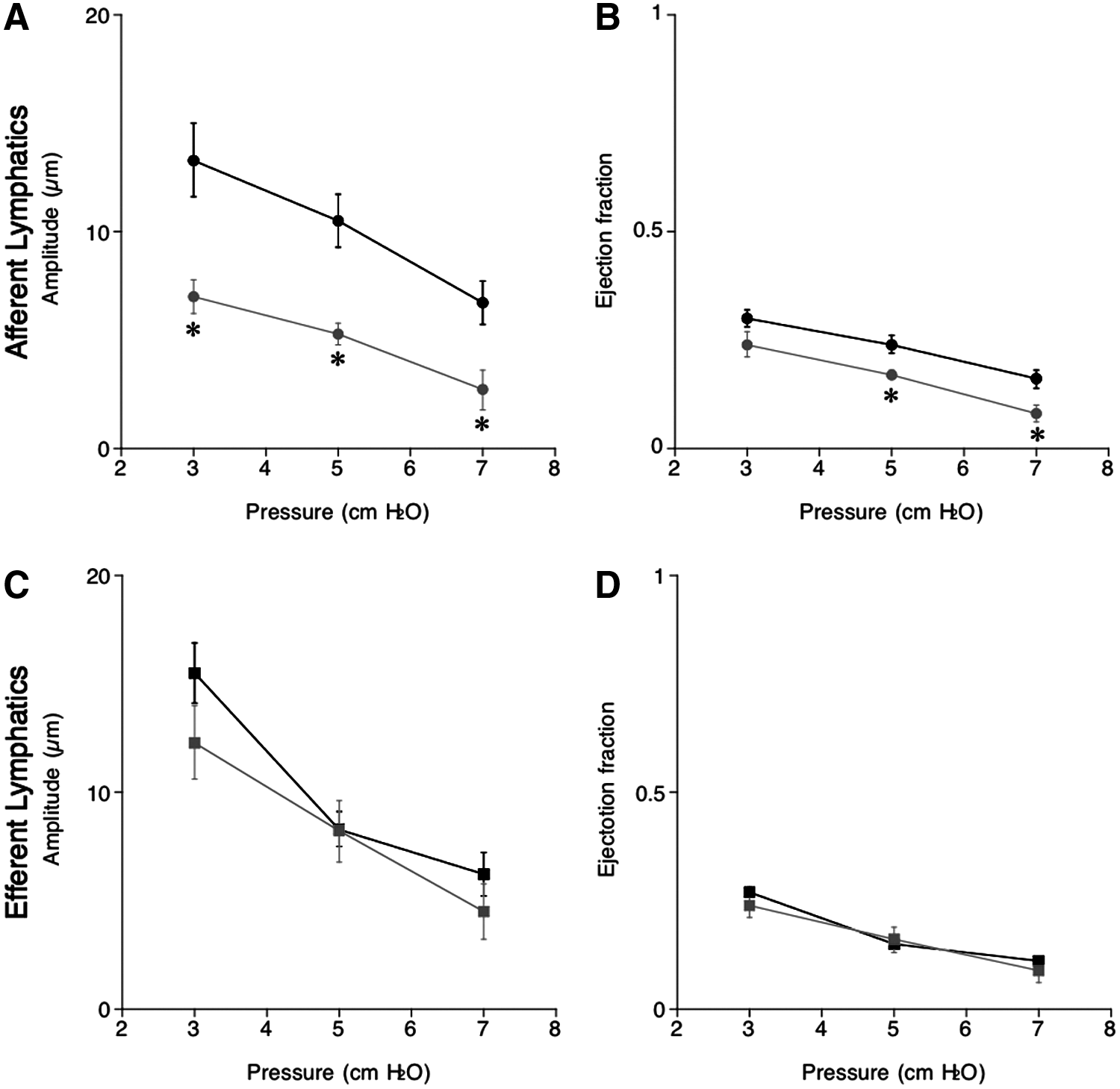
Changes in amplitude (μm,
In the present study, all of the afferent and efferent vessels with or without salt loading exhibited spontaneous oscillations of lymphatic diameter under intraluminal pressures ranging from 3 to 7 cmH2O. Increases in intraluminal pressures from 3 to 7 cmH2O augmented the frequency of afferent and efferent lymphatics in NSD and HSD mice, indicating the higher pressure exerts a positive chronotropic effect on lymphatic vessels (Fig. 4A, D). Though there was no significant difference in afferent lymphatic frequency between NSD and HSD mice (Fig. 4A), efferent lymphatics of HSD mice showed significantly higher frequencies compared with NSD mice at all pressure points measured (Fig. 4D).
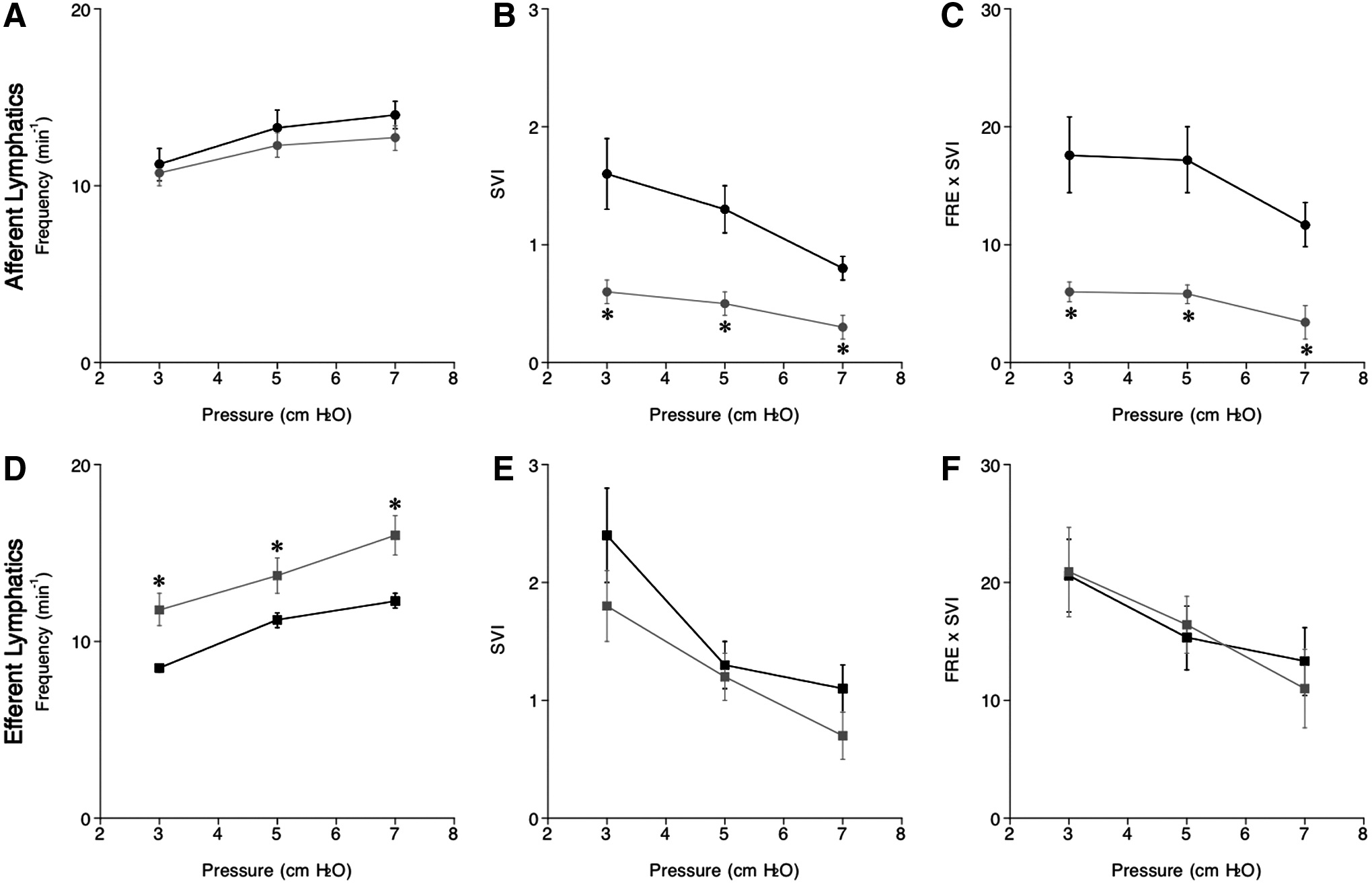
Intraluminal pressure-induced changes in frequency (min−1,
The increment in intraluminal pressure reduced SVI (Fig. 4B, E) and frequency×SVI (Fig. 4C, F) of afferent and efferent lymph vessels in NSD and HSD mice. Values of SVI and frequency×SVI of afferent HSD vessels were significantly smaller than those of NSD mice mainly due to marked reductions in the amplitude induced by a HSD (Fig. 4B, C). In contrast, SVI and frequency×SVI remained unaffected by a HSD (Fig. 4E, F). Effects of a HSD on parameters for lymph mechanical activity are summarized in Table 1.
Afferent and efferent lymphatics of the iliac lymph nodes were isolated from NSD and HSD ICR mice. We measured changes in diameters of afferent and efferent lymph vessels using video-microscopy and analyzed pressure-dependent mechanical activity of the vessels. Upper and lower arrows indicate increases and decreases of parameters. Right arrows indicate parameters that remained unchanged. EF, ejection fraction; FRE, frequency; SVI, stroke volume index.
Discussion
Spontaneous contraction of collecting lymphatics in mice
Diameters of both afferent and efferent lymphatics have been reported to oscillate in different tissues of various mouse strains,11,14–19 showing that intraluminal pressure is a critical factor for regulating oscillations. However, previous reports on lymphatic pump activity often lack direct measurements of pressure 19 or are limited to only afferent or efferent lymphatics in different species. For example, pressure-flow relationships of afferent lymphatics were demonstrated to form a bell-shaped curve in response to increments in transmural pressure in sheep mesenteric lymphatics, suggesting more resistance to pressure loading in comparison to bovine efferent vessels. 20 In the present study, we overcame this technical difficulty by isolating both afferent and efferent lymphatic vessels connected to identical lymph nodes of mice and direct compared pressure-dependent mechanical activity between afferent and efferent lymphatics. We have clearly demonstrated differential responses between afferent and efferent lymphatics to a HSD for the first time.
HSD alters the diameters and myogenic activity of afferent lymphatic vessels
Four weeks of a HSD reduced passive and active diameters in afferent, but not efferent, lymphatics in ICR mice. Calculated ejection fraction and SVI were also reduced by a HSD only in afferent lymphatics. In contrast to our observations, Kwon et al., demonstrated using a fluorescent tracer in FVB mice that 2 weeks of a HSD dilated afferent lymphatics in the inguinal lymph nodes, which receive lymph from part of the tail, and proposed a HSD increases lymph transport from the skin. 19
Contradictory results on responses of lymphatics to a HSD may be due to differences in mouse strain or the tissues examined. The iliac lymph nodes we used in the present study mainly drain lymph from the hind legs of ICR mice. Lymphatics of ICR mice appear to be resistant to a HSD because changes in mechanical parameters were not observed after 2 weeks of HSD loading, and it took 4 weeks before significant changes were observed.
Myogenic tone (% Dmax) and contractility (% Dmin) reflect intrinsic characteristics of generated forces by smooth muscle microvessels ex vivo. The % Dmax values of afferent and efferent lymphatics were 70%–80% with no significant differences between NSD and HSD mice. In our previous study, rats had an 80%–90% Dmax of afferent lymphatics of the iliac lymph nodes, suggesting that mice lymphatics have greater myogenic tone compared with rats.9,10 In contrast, mice lymphatics exhibited a larger % Dmin compared with rats (30%–40%), especially at a lower pressure range, indicating weaker lymphatic contractility.9,10 Thus, the smaller Dmax and the larger Dmin lead to a lower amplitude and ejection fraction. Although there were no significant differences in % Dmin of efferent lymphatics between NSD and HSD mice, HSD mice exhibited a significantly higher value of afferent % Dmin at a pressure of 5 cmH2O. These results indicate that a HSD decrease the contractility of afferent lymphatics only, however, myogenic tone remains unaffected in both afferent and efferent lymphatics. Collectively, alteration and reactivity of lymphatics in response to a HSD depend on the murine strain, location of lymphatics, and period and intensity of salt loading.
HSD enhances chronotropic effects on efferent vessels and reduces inotropic effects on afferent lymphatics
Frequency of lymphatic contraction is known to increase in response to elevation of intraluminal pressure. In this study, a HSD significantly increased the frequency of efferent, but not afferent, lymphatics in iliac lymph nodes even at the lowest pressure of 3 cmH2O. Kown et al. 19 reported in the inguinal lymphatics of FVB mice that the frequency of both afferent and efferent lymphatic contraction increased up to ∼200%, showing much higher responsiveness to a HSD than observed in our study. Discrepancies in responsiveness may be because of the method used, where the fluorescent dye indocyanine green (ICG) was injected into the tail for visualization of lymphatic flow. ICG could induce dose-dependent modulation of lymphatic contraction as well as oscillation frequency. 21
Effects of a HSD on pumping activity were different between afferent and efferent vessels. The pumping activity, measured and calculated as amplitude, EF, SVI, and frequency×SVI, was decreased by a HSD in afferent lymphatics. In contrast, efferent lymphatics were resistant to a HSD and pumping activity was preserved after 4 weeks of a HSD by enhanced chronotropism.
A previous report performed in cynomolgus macaques revealed that chronic responses of endothelial cells to inflammation were different between afferent and efferent lymphatics. 22 Thus, intrinsic characteristics of endothelial cells may be different between afferent and efferent lymphatics and endothelial responsiveness may be locally regulated. Additionally, high-salt-induced local differences of intraluminal physiological or chemical conditions may cause differential responses between afferent and efferent lymphatics. However, how high salt loading exacerbates pumping activity of only afferent lymphatics remains unclear.
Differences between the lymphatic mechanical activities in rats and mice
We recently demonstrated in afferent lymphatics of rat iliac lymph nodes that a 2-week HSD causes a compensatory increase in pump efficiency (SVI and frequency×SVI) compared with NSD rats. 23 This previous study seems contradictory to our present report showing that murine afferent lymphatics are vulnerable to a 4-week HSD. We speculate the different responses to HSD are mainly because of differences in the species used in the experiments and period of salt loading. Mechanical activities of lymphatics are intrinsically heterogenous depending on the anatomical location 24 and body size. The distance between inguinal inlets at the abdominal cavity and iliac lymph nodes is relatively shorter in mice than in rats, suggesting a more passive role of mice afferent lymphatics of iliac lymph nodes. In fact, the lymphatics of NSD mice had weaker lymphatic contractility, and the frequency of spontaneous contraction of NSD mice (∼10 min−1) was approximately half of NSD rats (∼20 min−1) at a pressure of 3 cmH2O. 9 Thus, mechanical activity responses to a HSD could be different even if the location of lymphatics is the same.
Roles of collecting lymphatics in hypertension
A recent report suggests that a HSD causes VEGF-C dependent hyperplasia of lymph capillaries in the rodent ear and increases the initial lymph draining space by working as a buffering system of excess body fluid and regulating blood pressure.13,25 Na+ content in the muscles of hypertensive patients are larger than normotensive individuals, as revealed by 23 Na magnetic resonance imaging. 26 Thus, accumulating evidence indicates that Na+ overloading in the interstitial spaces and draining capacity of the lymphatic system are associated with the pathophysiology of a certain form of salt-induced hypertension.
In support of the hypothesis by Machnik et al. that a HSD induces VEGF-dependent lymphangiogenesis, high salt intake in individuals from a rural community of northern China has been reported to increase plasma VEGF-C levels in salt-sensitive subjects more than in salt-resistant subjects 27 and in patients with refractory hypertension. 13 In the present ex vivo study, we demonstrated for the first time that a HSD induces functional changes in pre- and post-nodal collecting lymph vessels. A HSD for 4 weeks inhibited contractility of afferent lymphatics and increased the frequency of efferent lymph vessels of the iliac lymph nodes in ICR mice, suggesting that these lymphatics could adapt to high-salt conditions. Although the mechanism of such functional adaptation remains unclear, VEGF may be involved because VEGF-C and VEGF receptor-3-dependent mechanisms can stimulate lymphatic pump activity in the normal rat mesentery. 28
Further investigation needs to clarify the roles of collecting lymphatics in hypertensive patients and the mechanisms on how a HSD differentially modulates the mechanical activity of afferent and efferent lymphatics. In conclusion, mechanical activities of afferent and efferent collecting lymphatics in iliac lymph nodes are similar in ICR mice, and a HSD suppresses pumping activity in afferent lymphatics only and enhances chronotropism in efferent lymphatics leading to preservation of local pumping activity.
Footnotes
Acknowledgments
The authors would like to thank Novartis Pharma K.K. for their financial support.
Author Disclosure Statement
No competing financial interests exist.
Funding: This study was supported in part by a grant-in aid from the Ministry of Education, Culture, Sports, Science and Technology of Japan (No. 25670748 to RM, No. 21590948 to MI).