
Abstract
Abstract
Background:
In the murine cornea, which is an established model for analyzing pathologic lymphatic vessel growth, phenotypic heterogeneity of the endogenous lymphatic vessels in the limbus of the cornea was previously described. In this study, the cornea of BALB/c, C57BL/6, and FVB mice with different limbal lymphangiogenic phenotypes was analyzed to identify novel candidates potentially influencing lymphatic vessel growth.
Methods and Results:
Pathway specific expression analysis of the cornea was performed to identify novel candidate genes. Corneal protein expression of the respective candidates was analyzed by fluorescent immunohistochemistry. The effect of the candidates on proliferation of human dermal lymphatic endothelial cells (HDLECs) was analyzed by BrdU proliferation ELISA. Thirteen genes were differentially regulated in corneas of mouse strains with more endogenous limbal lymphatic vessels (high-lymphangiogenic) (C57BL/6) compared to mouse strains with less endogenous limbal lymphatic vessels (low-lymphangiogenic) (BALB/c, FVB). Two candidates, Tumor necrosis factor (ligand) superfamily member 10 (Tnfsf10/Trail) and Plasminogen activator, tissue (Plat/tPA) were expressed in the cornea of BALB/c and C57BL/6 mice on the protein level. In vitro, Trail and recombinant tPA inhibited the proliferation of human dermal lymphatic endothelial cells.
Conclusion:
Molecular analysis of the naive cornea in mouse strains with different limbal lymphatic phenotypes is a valuable model to identify novel endogenous regulators of lymphangiogenesis.
Introduction
L
Several endogenous molecules that promote lymphangiogenesis were already identified, such as the main lymphangiogenic growth factors VEGF-C and -D, which directly affect lymphatic endothelial cell proliferation/growth via VEGFR32,3 and other growth factors like fibroblast growth factor-2 (FGF-2) 4 or hepatocyte growth factor (HGF). 5 However, in contrast to numerous identified anti-hemangiogenic factors, inhibitors of lymphangiogenesis are far less investigated. In the physiologically lymphatic vessel-free cornea, which is a widely used in vivo model for analyzing the growth of pathologic blood and lymphatic vessels, TSP1, 6 soluble VEGFR-2, 7 soluble VEGFR-3, 8 and Vasohibin (VASH1) 9 were shown to inhibit lymphangiogenesis. Hence, the normally transparent nature and good accessibility of the cornea is of great benefit for vascularization studies, as it enables a precise morphometrical analysis of specifically stained lymphatic vessels in corneal flat mounts. 10 The effect of different molecules on the outgrowth of new blood and lymphatic vessels from existing ones (neovascularization) can be nicely studied in different assays (e.g., the micropocket assay 11 or the suture-induced neovascularization assay).12,13 Additionally, the cornea maintains its avascularity through several mechanisms, including the expression of anti-hem and lymph- angiogenic factors6–8,14–16 and therefore provides a suitable source to identify novel inhibitors of lymphangiogenesis.
Although the normal central cornea is free of both lymph and blood vessels, in the transition zone between the vascularized conjunctiva and the avascular cornea (the limbal region), both vessel types are physiologically present (Fig. 1). Previously several groups demonstrated strain-specific heterogeneity of endogenous lymphatic vessels in the murine cornea.17–19 The endogenous resting limbal lymphatic vasculature (i.e., lymphatic vessels found in the limbal region of the naïve cornea) was higher in C57BL/6 mice compared to BALB/c and FVB mice. 19 Naïve corneas of C57BL/6 mice also contain more CD11b+ cells compared to BALB/c. 17 These strain-specific differences suggest genetic factors and expression differences of genes influencing lymphatic vessel growth to be involved in the different limbal lymphatic vessel phenotypes.
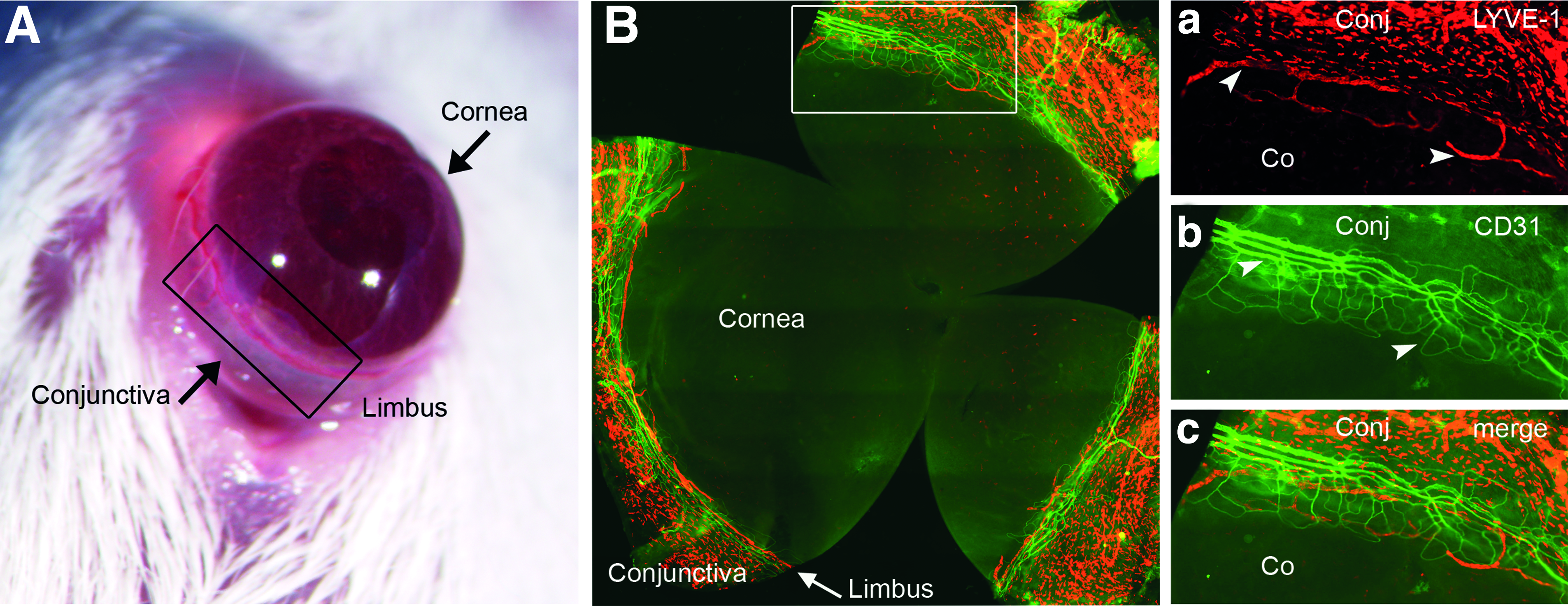
Limbal transition zone between physiologically avascular cornea and physiologically vascularized conjunctiva.
Based on these observations, we hypothesized that analyzing the naïve cornea of phenotypically different strains on a molecular level might be a suitable approach to identify candidate genes that have an effect on lymphatic endothelial cells. In this study we provide first evidence for the feasibility of using the naïve cornea of mice with phenotypic different resting limbal lymphangiogenesis as a powerful model to identify novel endogenous modulators of lymphangiogenesis.
Materials and Methods
Corneal tissue preparation
Corneal tissue samples were taken from BALB/cAnNCrl (BALB/c), C57BL/6NCrl (C57BL/6), and FVB/NCrl (FVB) female mice between 6 and 9 weeks of age (Charles Rivers Laboratories, Germany). Mice were euthanized and the central 2 mm Ø of the corneas were excised, rinsed in PBS, transferred into cryogenic vials, and quickly deep frozen in liquid nitrogen to prevent RNA degradation. Experiments (tissue harvesting) were conducted in accordance with the institutional and national guidelines for the care and use of laboratory animals.
RNA isolation
Corneas from two mice per strain were pooled for expression analysis. Corneal tissue samples were disrupted and homogenized using the Precellys® 24 tissue homogenizer in RLT lysis buffer from Qiagen. Total RNA was extracted using the RNeasy kit (Qiagen, Hilden, Germany) including an on-column DNase I digestion step according to the manufacturer`s instructions. RNA purity and concentration was assessed spectrophotometrically using NanoDrop.
cDNA preparation
RT 2 Profiler PCR arrays
First strand cDNA synthesis was carried out with 0.5 μg of total RNA isolated from corneal tissue using the recommended RT 2 first strand kit (SABiosciences) including a residual genomic DNA elimination mixture (containing 0.5 μg RNA, 5x gDNA elimination buffer (2 μL) and H2O to final volume of 10 μL). The contents were mixed gently, incubated for 5 min at 42°C, and chilled on ice. For the first strand cNDA synthesis 10 μL RT cocktail (5x reverse transcription buffer, Primer and external control mix, RT Enzyme Mix3, and H2O) were mixed with 10 μL genomic DNA elimination mixture and incubated at 42°C for 15 min. The reaction was stopped by heating at 95°C for 5 min, and finally 91 μL H2O were added.
Reverse-transcription quantitative real-time PCR (RT-qPCR)
RT 2 Profiler PCR Arrays
The “Mouse Endothelial Cell Biology” (PAMM-015A, SABioscience) and the “Mouse Angiogenesis” Array (PAMM-024A, SABioscience) were used to analyze differentially expressed genes in corneal tissue. RT-qPCR was performed on MyiQ thermal cycler with iQ5 software (Biorad).
Reaction set up and conditions
102 μL of the diluted cDNA synthesis reaction were mixed with 1275 μL of the 2x RT 2 qPCR Master Mix and 1173 μL H20 to a final volume of 2550 μL. 25 μL of the experimental cocktail were added to each well of the PCR array. PCR was performed with the recommended two-step cycling program, including a 10 min step at 95°C to activate the HotStart DNA polymerase, and 40 cycles with an initial denaturation step at 95°C (15 sec) and an annealing step at 60°C (1 min).
Cell culture experiments
Adult human dermal lymphatic endothelial cells (HDLECs, Promocell) were cultured in complete endothelial cell growth medium MV2 (Promocell) containing 5% fetal calf serum (FCS), 5 ng/mL recombinant human epidermal growth factor, 10 ng/mL recombinant human basic fibroblast growth factor, 20 ng/mL insulin-like growth factor, 0.5 ng/mL recombinant human vascular endothelial growth factor 165, 1 μg/mL ascorbic acid, and 0.2 μg/mL hydrocortisone (full medium). Minimal medium consisted of endothelial cell basal medium MV2 containing 0.5% FCS. HDLECs were used between passage six to nine.
Lymphatic endothelial cell proliferation ELISA
Cell proliferation was quantified indirectly by measurements of BrdU incorporation into newly synthesized DNA during cell proliferation (BrdU Cell Proliferation ELISA, colorimetric, Roche). Briefly, 100 μL of HDLECs were plated onto a 96-well plate at a density of 30,000 cells/mL. Cells were left to attach overnight in full medium. The next day the cells were incubated for 2 h in minimal medium. Next, medium was changed to either minimal medium or full medium supplemented with recombinant Trail (Abcam) or recombinant tissue plasminogen activator, rtPA (Actilyse®: alteplase, rtPA, Boehringer Ingelheim) in the denoted concentrations. BrdU was added 5 hours later to each well at a concentration of 100 μM. Cells incubated in minimal medium or full medium only were used as control.
After 24 and 48 hours, BrdU incorporation was analyzed: Cells were fixed, the DNA was denaturated by adding FixDenat, and the cells were incubated with a monoclonal anti-BrdU-antibody conjugated with peroxidase. After three washing steps, 100 μL substrate solution (tetramethyl-benzidine) were added to the wells. The reaction was stopped with 25 μL 1 M H2SO4 and absorbance was measured at 450 nm (with 690 nm as reference wavelength) on a microplate reader (BioTek).
For statistical analysis the absorbance values of the reference wavelength (690 nm) and blank values were subtracted from the absorbance values measured at 490 nm. Untreated control cells were set to 100% and values of the treatment group were related to the control. Statistical analysis was performed using GraphPad InStat and GraphPad Prism.
Immunohistochemistry
BALB/cN and C57BL/6N mice were obtained from the Animal Facility of the Center for Molecular Medicine Cologne (CMMC). Eyes were enucleated, embedded in OCT Tissue Tek Compound (Sakura), and cryosectioned to 8μm. Cryosections were fixed with acetone for 15 min, rinsed in PBS three times, and blocked in PBS with 2% BSA (PBSA) for 1 h in a humid chamber at room temperature. Cryosections were stained with the primary antibody (see below) in 2% BSA overnight. Next day, after washing with PBS (3×5min) the cryosections were incubated in the secondary antibody in PBS for 45 min in the dark. The cryosections were covered with mounting medium (DAKO, Germany) containing DAPI for nuclear counterstaining. The polyclonal rabbit anti-TRAIL antibody (1:100, Novus Biologicals) was used and rabbit IgG, polyclonal (1:100, Abcam) as isotype control. Tissue plasminogen activator (Plat/tPA) expression was analyzed with the monoclonal rabbit anti-tPA antibody (1:200, Abcam), and rabbit IgG, monoclonal (1:200, Abcam) as isotype control. Images were taken with a fluorescence microscope (Olympus BX53).
Statistical analysis
RT 2 Profiler PCR arrays
Three samples of each strain were collected and analyzed with the web-based RT 2 Profiler™ PCR array data analysis tool from SABiosciences (Delta-Delta Cq method (ΔΔCq) and Student`s t-test for relative quantification). Selection criteria were a more than ≥2-fold differential expression, a p-value ≤0.05, and a cut off for genes with a cq-value >32 in both sample groups to reduce unspecific results. Proliferation assays were analyzed with ANOVA and a multi comparison post-test if ANOVA p-values <0.05.
Results
Several angiogenesis and endothelial cell biology-related genes are differently regulated in corneas of BALB/c, C57BL/6, and FVB mice
The phenotypic analysis of naïve corneas 19 indicated strain-dependent differences in endogenous limbal lymphangiogenesis in a non-inflammatory setting. In the normal untreated cornea, the lowest endogenous limbal lymphatic vessel area was found for the BALB/c mice (“low-lymphangiogenic”), the highest for the C57BL/6 mice (“high-lymphangiogenic”), and an intermediate phenotype for the FVB mice. 19
To analyze whether the phenotypic differences may be reflected in expression differences between the strains, we analyzed the expression of 84 genes relevant to angiogenesis and endothelial cell biology by RT-qPCR in corneal tissue from BALB/c, FVB, and C57BL/6 mice. The C57BL/6 strain, which shows a high-lymphangiogenic phenotype in the limbus, was used as reference strain. In the angiogenesis array (Table 1, A), three candidates genes were identified to be differently regulated between the low-lymphangiogenic BALB/c and the high-lymphangiogenic C57BL/6 strain and nine candidates in the endothelial cell biology array (Table 1, B). Expression analysis of corneal tissue from the low-lymphangiogenic FVB and the high-lymphangiogenic C57BL/6 strain revealed five differently regulated candidates genes in the angiogenesis array (Table 2, A) and seven in the endothelial cell biology array (Table 2, B).
Modified from Reference 44.
Modified from Reference 44.
In BALB/c mice, the most upregulated gene (14.1-fold) was Tumor necrosis factor (ligand) superfamily, member 10 (Tnfsf10/Trail), the most downregulated gene (-5.1-fold) NLR family, apoptosis inhibitory protein 1 (NAIP1) (Table 1). Similarly in the FVB mice, Tnfsf10/Trail showed a 9.8-fold higher expression compared to C57BL/6 and was the most upregulated gene identified also in this experimental setup. Fibroblast growth factor 1 (FGF1) was downregulated 13.6-fold in the low-lymphangiogenic FVB (Table 2).
In summary, 13 out of 84 analyzed genes had significantly (p<0.05) different expression values between low- (BALB/c, FVB) and high- (C57BL/6) lymphangiogenic strains. Nine genes were altered between BALB/c versus C57BL/6 (Table 1), including Thrombospondin 1 (Thbs1) and Transforming growth factor, beta 1 (Tgfb1) and nine genes, including Thbs1 and Tgfb1 between FVB and C57BL/6 mice (Table 2). Five genes were identified in both low-lymphangiogenic strains, in BALB/c as well as in the FVB strain: Naip1, Tgfb1, Thbs1, Trail and Plasminogen activator, tissue (Plat/tPA). Among these genes, Trail was upregulated 14.1-fold in corneas of BALB/c and 9.8-fold in FVB mice compared to C57BL/6 corneas. Trail is a type II transmembrane protein belonging to the tumor necrosis factor family. 20 Similarly Plat (up to 4.6-fold) and Tgfb1 (up to 2.4-fold) were upregulated in the low-lymphangiogenic BALB/c and FVB corneas. Naip1 and Thbs1 mRNA expression was downregulated in BALB/c and FVB mice. Two of the candidates, which were regulated in both low-lymphangiogenic strains BALB/c and C57BL/6, Tgfb1 and Thbs1 are already known to have an influence on lymphangiogenesis.6,21 We therefore focused on Trail and tPA, which were the most upregulated candidates in both low-lymphangiogenic strains and analyzed their expression in the cornea on protein level.
Protein expression in BALB/c and C57BL/6 corneas
The protein expression pattern of Trail and tPA was analyzed in naïve corneas of the high-lymphangiogenic C57BL/6 and in BALB/c as a representative for a low-lymphangiogenic strain. Trail was clearly expressed in the epithelial and the endothelial layer of the cornea in both strains; the corneal stroma however, showed only a very weak staining (Fig. 2). For tPA, a strong expression in the corneal epithelium and endothelium and a weak expression in the stromal layer in the BALB/c and C57BL/6 strain (Fig. 3) was found. The immunohistochemical data confirm that beside mRNA expression Trail and tPA are also expressed on protein level in the cornea of both strains.

Trail protein is expressed in the epithelium and endothelium of BALB/c and C57BL/6 corneas.

Plat protein is expressed in the cornea of BALB/c and C57BL/6 mice.
Trail and rtPA inhibit proliferation of lymphatic endothelial cells
Various reports are available about the physiological effect of Trail on angiogenesis in vascular endothelial cells. Trail was reported to increase proliferation and to promote survival of vascular endothelial cells.22,23 In contrast to these findings, no effect in HUVECs and inhibition of hCMEC/D3 proliferation (a human brain endothelial cell line) was described. 24 We therefore were interested in Trail`s effect on the proliferation of lymphatic endothelial cells (HDLECs). Proliferation assays were carried out in minimal medium containing 0.5% FCS only and in full medium containing a combination of several growth factors to optimize cell proliferation and viability. Under full medium conditions we were not able to detect a significant effect on the proliferation of HDLECs either after 24 or 48 h treatment (Fig. 4A and 4C).However, when proliferation was analyzed in minimal medium, after 24 hours 10 ng/mL Trail and after 48 hours all tested concentrations (200 ng/mL, 100 ng/mL, and 10 ng/mL TRAIL) inhibited the proliferation of HDLECs up to 22% compared to the untreated control (Fig. 4B and 4D).
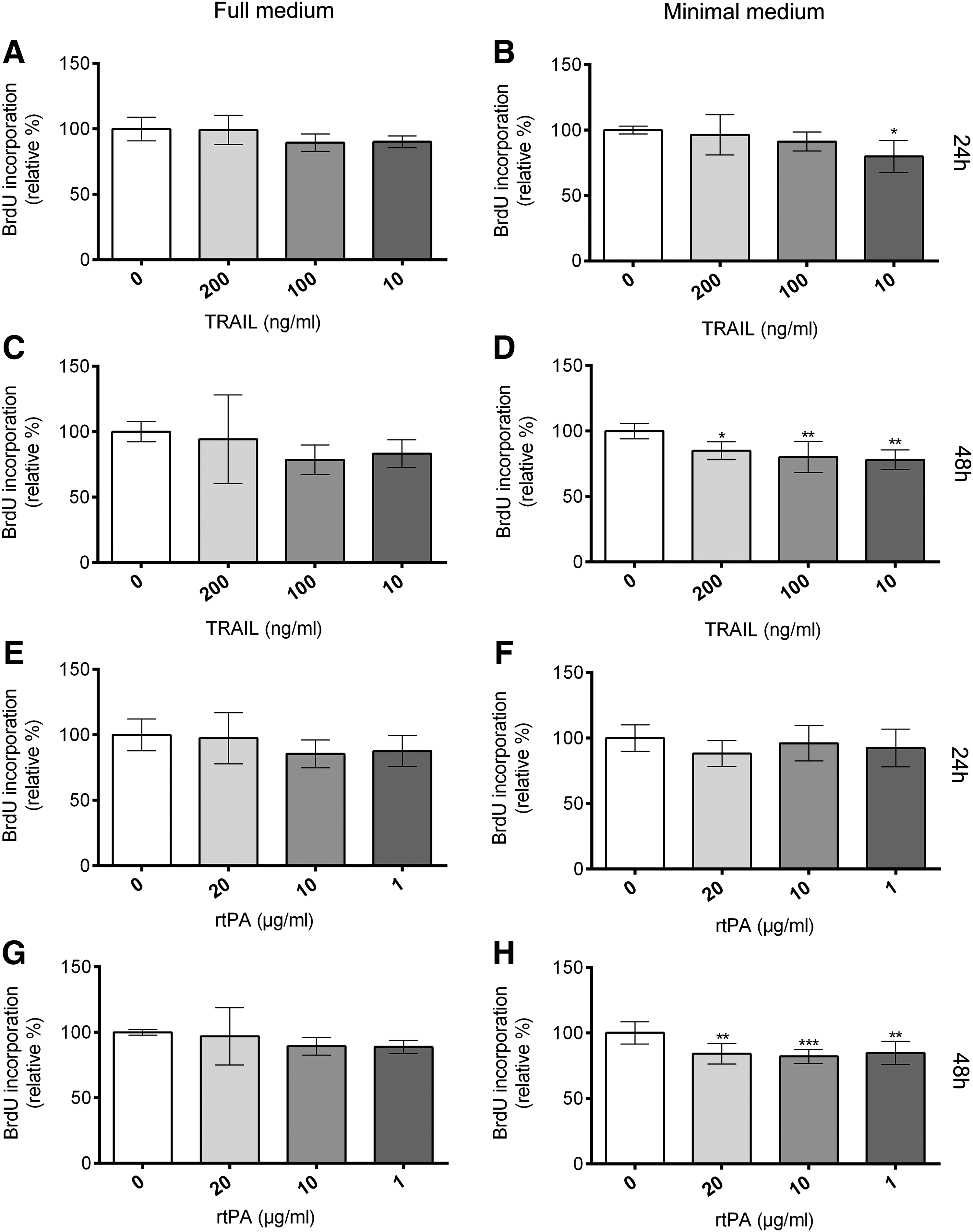
TRAIL and rtPA inhibit proliferation of lymphatic endothelial cells in vitro. BrdU incorporation into newly synthesized DNA was analyzed in HDLECs treated with 200 ng/mL, 100 ng/mL, and 10 ng/mL Trail (A-D) or 20 μg/mL, 10 μg/mL, and 1 μg/mL rtPA (alteplase, Actilyse®). HDLECs were seeded at a density of 30.000 cells/mL. Incubation was carried out in full medium
tPA was the second most upregulated gene in corneas of the low-lymphangiogenic BALB/c and FVB mice. tPA induces endothelial cell proliferation 25 and stimulates the expression of VEGF in the endothelial cell line ECV304. 26 We could not detect a significant effect of rtPA on endothelial cell proliferation of HDLECs in full medium. However, treatment of HDLECs cultured in minimal medium with 20 μg/mL, 10 μg/mL, and 1 μg/mL rtPA for 48 hours reduced BrdU incorporation significantly.
Conclusions
Corneal avascularity, the absence of blood and lymph vessels under normal physiological conditions, is actively maintained by several mechanisms.14–16,27 However, only few endogenous anti-lymphangiogenic factors such as sVEGFR-2 and TSP1 have been identified in the cornea so far.6,7 Recently, phenotypic analysis of corneal lymphatic vessels in the limbal transition zone between the avascular cornea and the physiological vascularized conjunctiva in several mouse strains revealed differences in the amount of endogenous lymphatic vascularization.17–19 These phenotypic differences between high-lymphangiogenic (C57BL/6) and low-lymphangiogenic (BALB/c and FVB) strains may reflect different genetic backgrounds that could influence tissue-specific mRNA expression of genes involved in the regulation of corneal lymphatic vessel growth and the restriction of lymphatic vessels to the limbal region in order to maintain an alymphatic cornea.
We therefore hypothesized that comparing corneal tissue on mRNA expression level could enable the identification of novel regulators of lymphangiogenesis. Using pathway-specific expression analysis of corneal tissue from C57BL/6, BALB/c and FVB mice 13 more than 2-fold differentially regulated genes related to angiogenesis and lymphatic endothelial cell biology were identified (Tables 1 and 2). Some of the identified genes are already known to have an effect on lymphangiogenesis, such as Ephrin B2,28,29 endothelial PAS domain protein 1 (Epas1), 30 FGF1, 31 Tgfb1,21,32,33 and Thbs1. 6 Therefore it seems that our approach indeed enables identification of regulators of lymphangiogenesis. Thbs1 can inhibit inflammatory corneal lymphangiogenesis via downregulation of macrophage-released prolymphangiogenic VEGF-C. 6
However, in our assay Thbs1 mRNA was expressed upto 2.8-fold higher in the high-lymphangiogenic C57BL/6 mice. This is consistent with expression data from growth factor stimulated skin biopsies of C57BL/6 mice who showed a 2.95-fold upregulation of Thbs1 compared to FVB mice. 34 Since presence and concentration of several cytokines in the environmental tissue can influence vessel growth, 35 the upregulation of Thbs1 may reflect that the concerted interaction of several pro- and anti-angiogenic molecules instead of a single factor is critical for the growth characteristics of the vasculature. Similar, in the above mentioned skin biopsy assay the “low-growth” C57BL/6J strain had higher mRNA levels of pro-angiogenic factors (VEGF and placental growth factor). 34
Tgfb1 mRNA was upregulated in the cornea of both low-lymphangiogenic BALB/c and FVB strains. This is in line with recent studies about the function of Tgfb1 in lymphangiogenesis, which suggest an inhibitory effect of Tgfb1 on proliferation, VEGF-C induced migration of HDLECs, 21 and lymphatic regeneration during wound repair. 32 However, in an inflammatory rat model Tgfb1 induces expression of VEGF-C and lymphatic vessel growth, 33 again suggesting that the overall condition and the balance of several factors is important for vessel growth.
Five candidates, including the above mentioned Tgfb1 and Thbs1, were similarly expressed in both low-lymphangiogenic strains (FVB and BALB/c) compared to the high lymphangiogenic C57BL/6. Among these, two novel candidate genes (Plat / tPA and Trail) were identified, which to our knowledge were not described to influence lymphangiogenesis before. Trail was the most upregulated gene in the cornea of BALB/c and FVB (with less endogenous lymphatic vessels in the limbal area). Our data confirm the expression of Trail mRNA in the eye 36 and show that Trail is expressed in the epithelium and endothelium of both low- and high- lymphangiogenic strains. The upregulation of corneal Trail in both low-lymphangiogenic strains in the mRNA analyses suggests an inhibitory effect on lymphangiogenesis. The BrdU proliferation assay in full medium showed no effect on HDLECs. However, under starvation conditions of minimal medium with 0.5% FCS only, all three tested concentrations inhibited the proliferation of HDLECs after 48 hours treatment.
In the literature, Trail's effect on endothelial cells seems to be ambiguous and ranges from no effect on proliferation or migration in HUVECs, 24 the inhibition of proliferation and migration in the human brain endothelial cell line hCMEC/D3, 24 to an increase in proliferation of HUVECs comparable to VEGF 23 and the induction of migration. 22 It is assumed that Trail is able to induce apoptosis in endothelial cells only in the presence of specific conditions such as serum- and extracellular matrix deprivation. 37 Since we found reduced BrdU incorporation under conditions of minimal medium (0.5 % FCS) but not with full medium, the reduced proliferation rate reported herein may therefore be due to apoptosis. In a corneal transplantation model, grafts treated with TRAIL had a better survival time and more apoptotic cells compared to the control. 38 In contrast to other studies we could not reproduce the pro-angiogenic role for TRAIL reported for HUVECs in HDLECs.22,23
The second candidate gene tPA was upregulated in both low-lymphangiogenic strains BALB/c and FVB (4.6-fold and 2.9-fold, respectively), plasminogen activator urokinase (Plau/u-PA) only in the BALB/c mice. Plat/t-PA and Plau/u-PA can convert plasminogen to active plasmin 39 and play a role in extracellular matrix degradation in endothelial cells. 40 The presence of corneal plasminogen activator was already demonstrated n=6 from 3 experiments (A–E, G); n=8 from 4 experiments (F, H).41,42 In BALB/c and C57BL/6 mice, tPA was mainly present in the epithelium and endothelium of the cornea. Although tPA induces endothelial cell proliferation 25 and the expression of VEGF in endothelial cells, 26 in our study rtPA (alteplase) inhibited incorporation of BrdU in HDLECs. An inhibitory effect by rtPA was previously shown on bFGF induced proliferation in HUVECs. 43 Since the inhibitory effect of rtPA was not specific for endothelial cells, but was also found for the breast and prostate tumor cell lines MCF-7 and DU145, Carroll et al. suggested that other mechanisms like proteolytic degradation are responsible for proliferation inhibition by rtPA in HUVECs. 43 However, the exact mechanisms of the anti-angiogenic properties of rtPA in HDLECs need to be studied in further detail.
In summary, using pathway-specific expression analysis of corneal tissue from phenotypically different low- (BALB/c and FVB) and high- (C57BL/6) lymphangiogenic strains 13 differentially regulated candidate genes were identified. The candidates include genes that are already known to influence lymphangiogenesis (e.g., Tgfb1, Thbs1, EphrinB2), but more importantly also novel candidates (Trail, tPA) were identified and shown to have an effect on lymphangiogenesis in vitro. This led us to conclude that expression analysis of the naïve cornea in strains with different endogenous lymphatic vessel phenotypes is useful to identify potential candidates for novel endogenous regulators of lymphangiogenesis. In addition, the newly identified candidates may also contribute to the corneal anti-lymphangiogenic privilege. Comparing the limbal lymphatic vasculature of the naïve cornea in low- and high-lymphangiogenic strains (e.g., together with additional genetic mapping strategies) could provide a powerful tool to analyze the underlying genetic basis of lymphangiogenesis.
Footnotes
Author Disclosure Statement
No competing financial interests exist for all authors.
This study was supported by the Interdisciplinary Center for Clinical Research (IZKF) Erlangen (A9), German Research Foundation (DFG Cu 47/4-1, 47/4-2, Cu 47/6-1) and SFB 643 (B10), EU (FP7 “STRONG”), EU (FP7 COST “BM1302”) Ruth und Helmut Lingen-Stiftung Cologne (to CC), FOR 2240 (to BR, DH, FB, CC), National Eye Institute EY015472 (to SM).