
Abstract
Abstract
Background:
Axillary nodes dissection (AND) is an important risk factor for the appearance of breast cancer-related lymphedema (BCRL). The anatomy and pathophysiology leading to the development of BCRL after AND are not completely understood. Despite the existence of lymphedema models after AND, none of them were able to create lymphedemas without additional chemical inflammatory drugs or auxiliary physical techniques (radiotherapy). In this study, we aimed to describe the anatomical changes of AND on a rat's front leg before and after a new surgery technique.
Methods and Results:
AND was performed on seven Wistar rats with a new, posterior surgical approach. Indocyanine green mapping was done before and after surgery to detect “normal and secondary superficial lymphatic pathways” of the operated rat's front legs. Twelve months after surgery, dissections were performed. Subcutaneous blue dye injection of the hand was used to observe superficial and deep lymphatic pathways. Postsurgery, an acute edema of arm and shoulder appeared and persisted for 14–21 days. However, none of the rats showed a chronic secondary lymphedema. In two cases, seromas also appeared. All rats showed substitution functional lymphatic pathways as perforating lymph vessels around the surgical sites.
Conclusion:
This is the first description of perforating lymph vessels as lymphatic substitution pathways after AND on rats. These results help to understand why a chronic secondary lymphedema could not be created in rats after AND without additional chemical or physical interventions.
Introduction
B
Therefore, to understand the impact of AND on the lymphatic system anatomy, a complete AND (deep and superficial accessory axillary nodes) on rat, and lymph flow resaturation is observed on the rat's foreleg after tracer injection.
Materials and Methods
The lymphatic system of the rat's foreleg
The superficial lymphatic system of the rat's foreleg has been described in 1915 by Job, 9 in 1971 by Tilney, 10 and in 2011 by Suami 11 . After a subcutaneous injection with a colored marker on the backside of the hand, this marker is transported in a lymphatic vessels pedicel following the ulnar and radial veins of the forearm. According to some authors, these vessels make a first relay in the node called “elbow node,”5,9 and then lateralize and ascend toward the shoulder following the brachial vein before joining the group of brachial nodes, also called axillary accessories nodes11–13 (Fig. 1). The presence of this intermediary nodal relay at elbow level is not mentioned by all authors in their anatomical descriptions.10–13 However, almost all authors agree that the main superficial lymphatic drainage pathway of the rat's foreleg makes a relay at the level of the brachial nodes. Then this goes deeper to the level of the inferoposterior part of the shoulder at the level of the cephalic part of the latissimus dorsi muscle and joins the axillary nodes10,11 (Fig. 2).
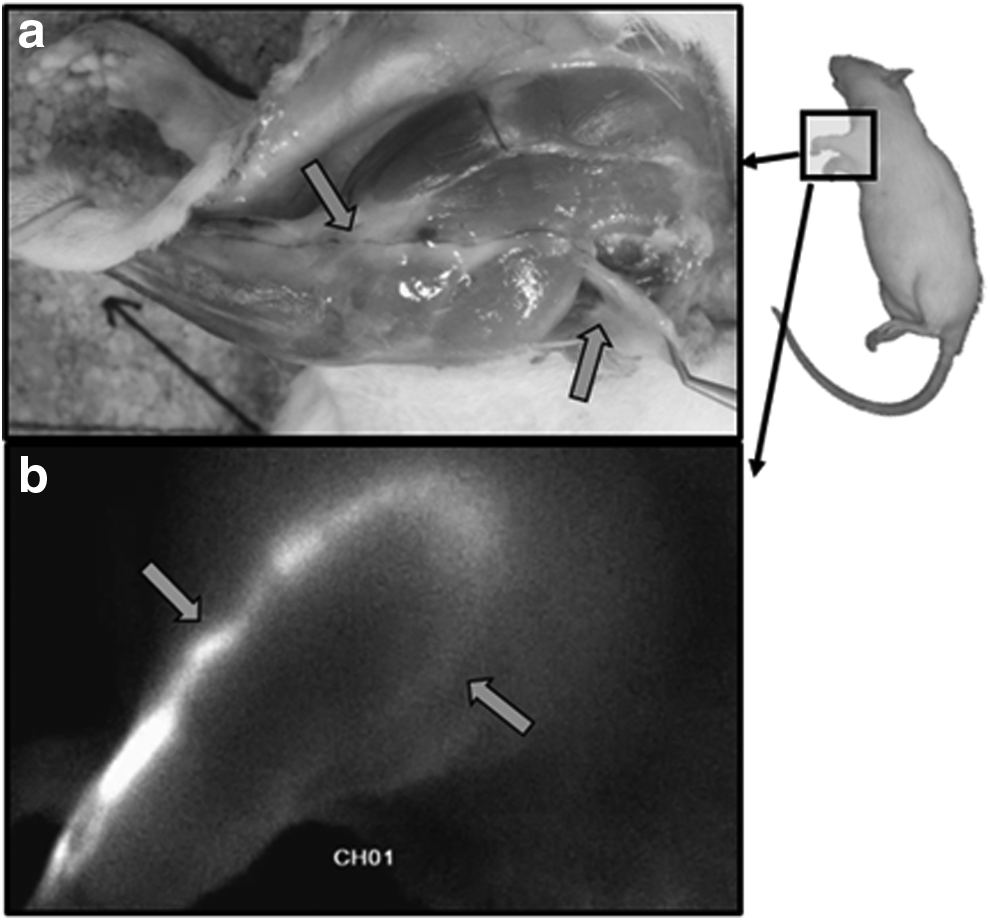
Macroscopic views of the lymphatic drainage way after the injection of Evans Blue in the back of the hand

Macroscopic and microscopic views of the brachial nodes hidden in the subcutaneous fat (right arrow) and of their efferent lymphatic vessels (middle arrow) going deeper to join the axillary nodes (black arrow).
From an anatomical point of view, the brachial lymphatic nodes are hidden in the fat tissue neighboring the posterior border of the brachial tissue (Fig. 3). They are mentioned as being overall a group of three lymphatic nodes, 10 and vascularized by one or two blood pedicels of which the superior vascular pedicel is always the most important. In general, there are three or four axillary nodes 10 (Fig. 3) lying deeply in the axillary cavity between the latissimus dorsi muscle and the rest of the thoracic cage, at the periphery of the junction between the thoracic cage and the axillary vein.
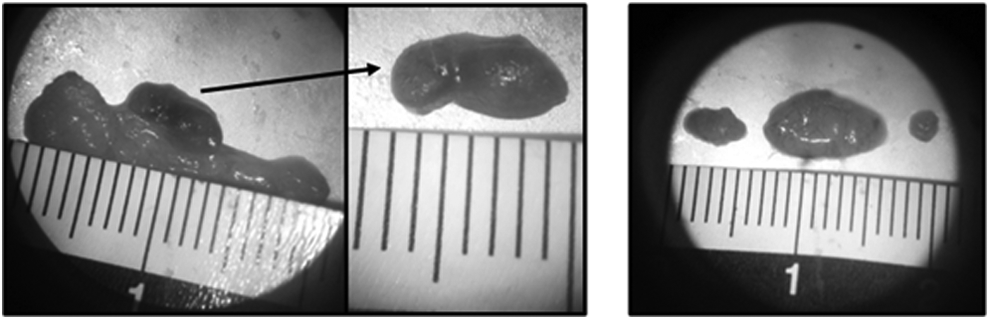
Microscopic views of the brachial nodes (left) and the axillary nodes (right) after resection.
Surgical technique of resecting the lymphatic nodes: posterior approach
The anesthetized animal, which has been shaven at the back level of the selected shoulder, is placed on heterolateral side. Its foreleg is in anterior elevation in the surgical support plane. A skin incision of 1 cm is made parallel to the posterior border of the triceps brachialis. Subsequently, the space between the interior skin side and the subcutaneous aponeurosis is split, permitting a direct access to the brachial nodes. The total fat tissue containing the brachial nodes is meticulously dissected, taking care not to transect the vascular pedicle(s) by which it is irrigated. These pedicles are also dissected and ligatured to prevent any hemorrhage (Fig. 4).

Macroscopic view of the fatty mass containing brachial nodes or axillary accessories after ligature of the vascular pedicle before final resection.
Access to the axillary cavity is made through an interaponeurotic passage between the cutaneous trunci and latissimus dorsi muscles (Figs. 2 and 5). Muscles were separated without incision (discision), permitting a direct entrance to the axillary cavity. Here, a fatty mass, in which the axillary nodes are entangled with the vasculo-nervous axillary mass, is seen. This intramuscular access is kept open by retractors. The axillary nodes hidden in the cavity are then delicately searched for, dissected, and resected, taking care that the elements of the peripheral vasculo-nervous tissues were not injured (Fig. 5). No vascular ligature is made on that spot. Then, the retractors are taken away and the muscles are carefully put back in place. Only the skin incision is closed again with stitches.

Macroscopic views of the posterior surgical approach. Global view of the anatomic connections between the teres major (blue arrow) and cutaneous trunci muscles (grey arrow) and the long portion of the triceps brachii (black arrow)
Experimental protocol
The experiments were done on seven Wistar rats (two females and five males) weighing averagely 560 g. The experiments were approved by the VUB committee of animal welfare.
Preparation and anesthesia of the animals
At the induction of isoflurane (3 minutes, O2 at 1 L/min, isoflurane at 3%), each animal received an analgesic medication administered subcutaneously (metacam: 2 mg/kg) and also eye salve to prevent eye dehydration. Subsequently, the animals were kept under general anesthesia by intraperitoneal injection of pentobarbital (Nembutal: 50 mg/kg) for about 3 to 4 hours. During the first minutes of anesthesia, the skin surfaces of the selected shoulders were shaven and prepared for intervention. Before skin incision and during intervention, a local anesthesia (0.5–1 mL of xylocaine at 2%) was administered.
Thereafter, ablation surgery of the nodes was done as already described.
The average time for surgical intervention was 45 minutes.
Mapping of the superficial lymphatic pathways
A first series of illustrations of the superficial lymphatic pathways of the selected forelegs was done with all animals at baseline before intervention (T0). The indirect “in vivo” lymphographies were done by indocyanine green (ICG) after general anesthesia to have a mapping of the normal lymphatic evacuation pathways. Then 0.05 mL of ICG Pulsion 50 mg (Pcode: PICG0050BE, 5 mg/mL; Pulsion Medical Systems AG) was subcutaneously injected in the back side of the hand of the foreleg.
A 3 minutes massage of the injection spots was done to enhance the absorption of the tracer by the initial lymphatics and to help its evacuation in the lymphatic vessels. 14 The indirect lymphographies were done with the PDE system (PhotoDynamic Eye; Hamamatsu Photonics Co.).
To compare the evolution of the lymphatic drainage pathways, a second series of ICG mappings of the superficial lymphatic vessels was done under general anesthesia (induction 3 minutes, O2 at 1 L/min, isoflurane at 3%, maintenance under mask, O2 at 0.2–0.5 L/min, isoflurane at 1.5%–2%), according to the same method as the first mappings, but at different time intervals (60, 30, 15, and 7 days after surgeries, Table 1).
T1, time periods between the operation and the second indocyanine green examination; T2, time periods between surgery and the final dissection examination.
Follow-up of the legs’ volume changes
The volume changes were observed visually by two evaluators (identical throughout the whole study) and compared with the nonoperated contralateral sides for a period of 12 months generally, and in the following way: every day of the first postoperative month, every third day during the second month, and finally every 15th day from the third month on.
Final dissections of the lymphatic vessels
After 12 months averagely, final dissections were done after euthanizing the animals. Then 0.05 mL of Evans Blue (Pcode: 1001313745, 2.5 mg/mL; Sigma-Aldrich Co) was injected bilaterally and subcutaneously in the backsides of the hands of the rats’ forelegs immediately after their death, to analyze the superficial and deep lymphatic pathways. Based on the same principle as already described, a 3 minutes massage of the injection sides was done before and during dissections.
Results
Resection of the lymphatic nodes
The number of resected brachial nodes (seven cases on seven) varies between two and four juxtaposed lymph nodes (Fig. 3). Concerning the axillary nodes, a typical morphology was found in all cases during surgical ablation. Presence of three lymphatic nodes, namely, a node of intermediate size, a node of big size, and a node of inferior size (Fig. 3).
Morphological changes of the operated forelegs
After the brachial and axillary dissections, an acute postsurgical edema of the arm and the shoulder appears in all cases (Fig. 6). However, this acute edema does not lead to chronic lymphedema of the foreleg (seven cases on seven). Indeed, this volume increase persists only for 14 to 21 days on average, except for one rat (35 days). After this postsurgical period, no visible volume difference could be observed between the operated legs and the reference heterolateral forelegs. In no case, an edema of the forearm and the hand of the operated legs appeared.
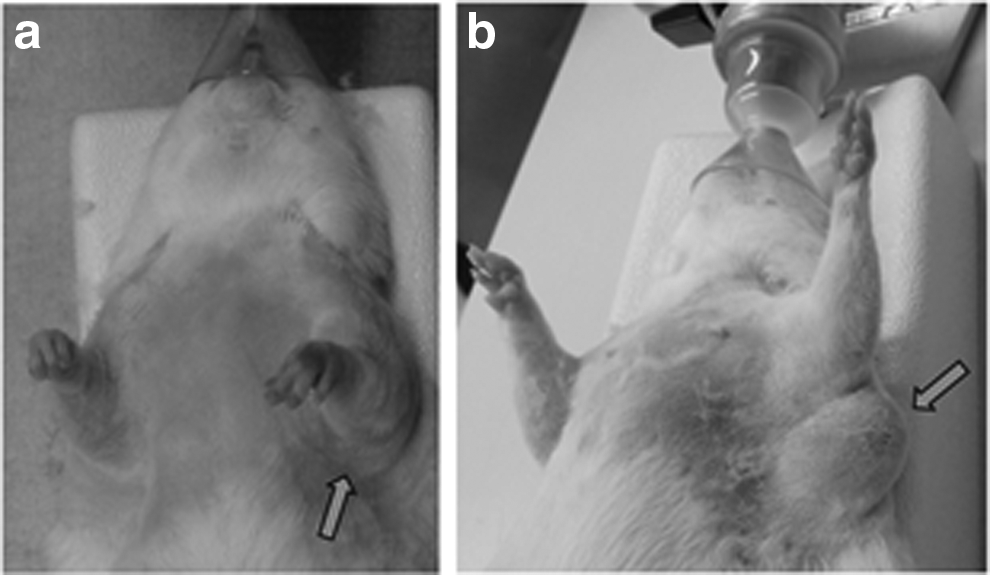
Presence of a light acute postoperative edema of the left foreleg (arm and shoulder) 28 days after surgery
A lymphocele appeared only in two operated rats. A first lymphocele appeared 3 to 5 days after surgery and reached a maximum volume 6 days after the operation and disappeared completely after 61 days (Fig. 6). The second lymphocele appeared 5 to 6 days after the intervention and resorbed completely in 70 days. Lymphoceles are not considered as foreleg secondary lymphedema.
ICG mappings
Compared with the normal lymphatic pathways of the rat's forelegs, the mappings after surgery showed functional superficial lymphatic pathways (Fig. 7).
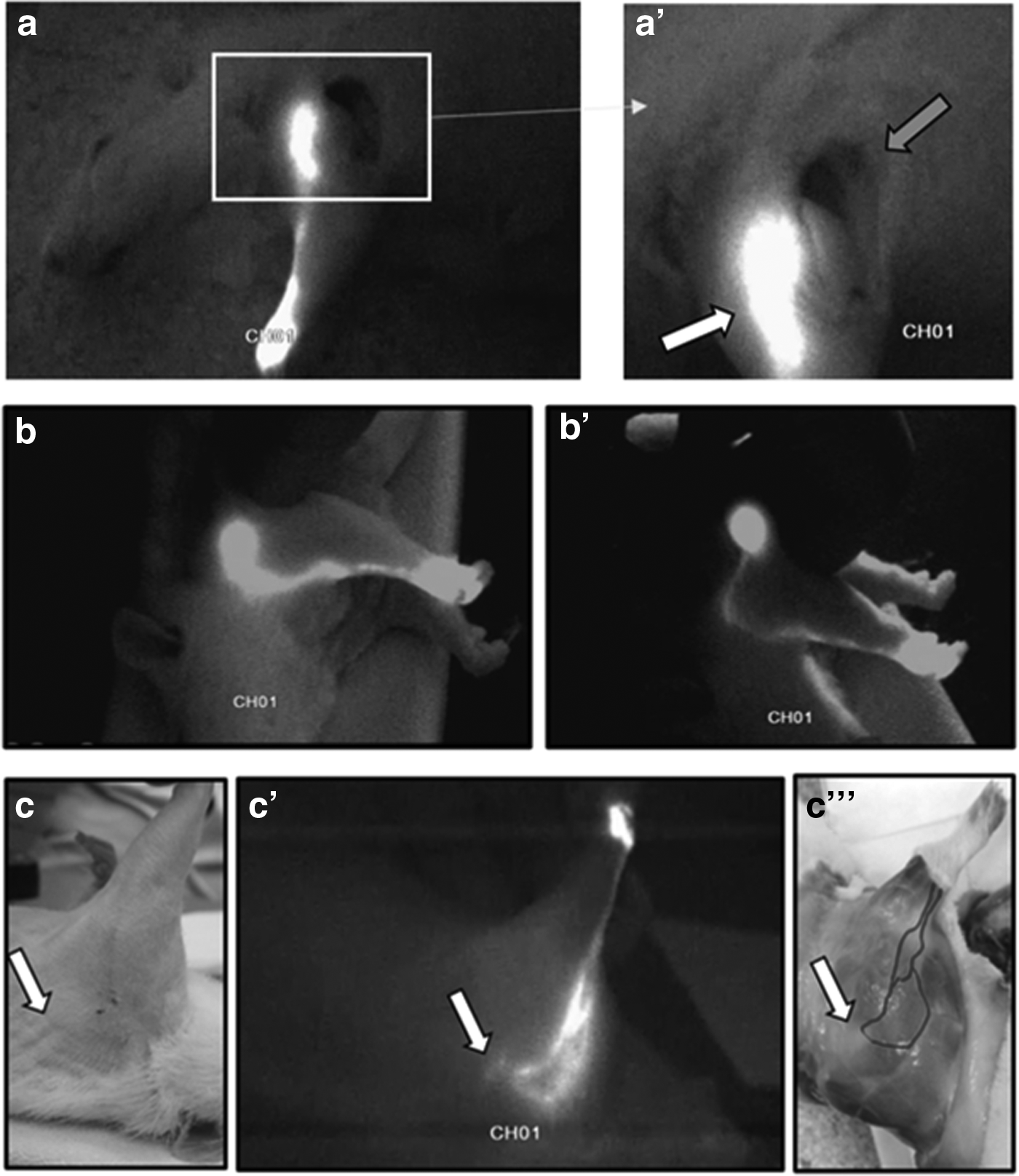
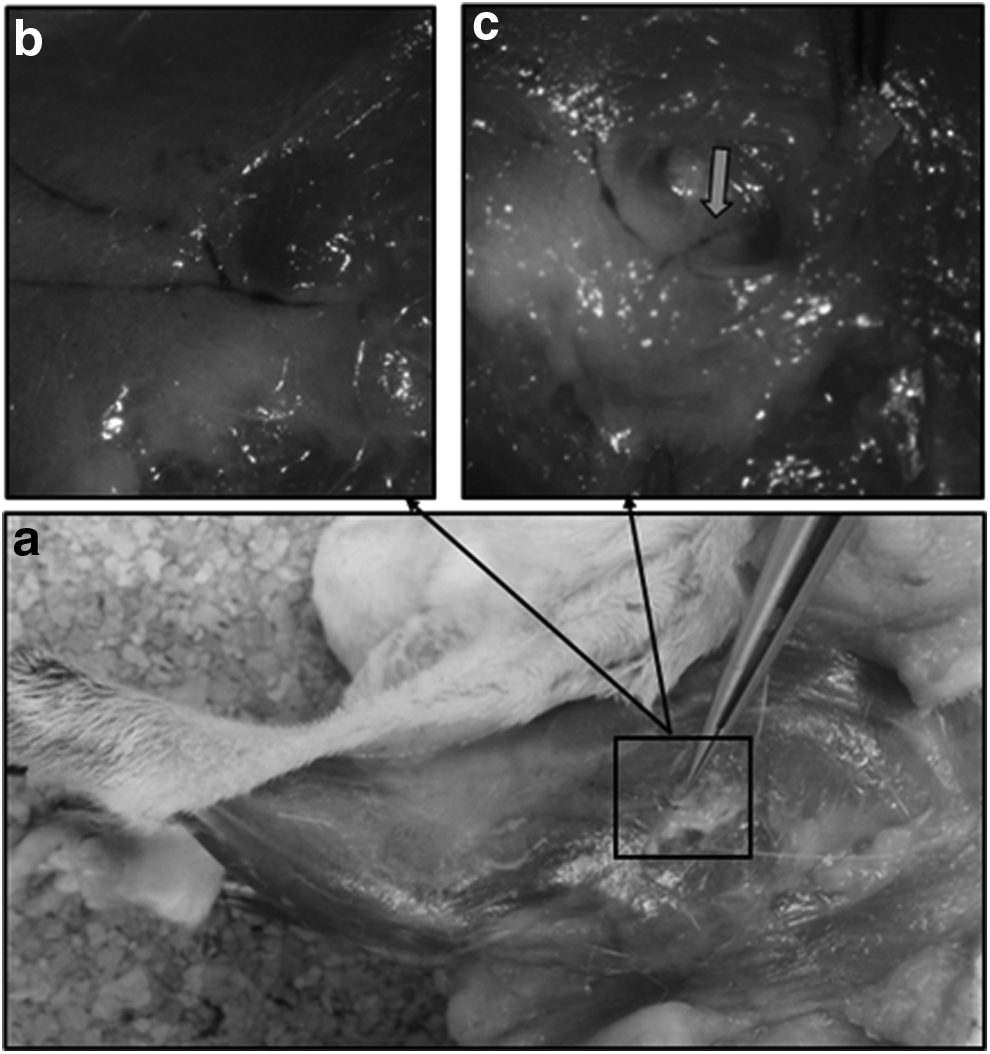
Main drainage lymphatic pathway of the foreleg after injection of ICG. This pathway was interrupted through ablation of the brachial nodes. A leak of the fluorescent tracer can be seen above the spot where the ganglion has been taken away (grey arrow) (aa′). Observation of the lymphatic drainage (after distal injection of ICG) flowing off in a lyphocele (mobilized by the fingers of the surgeon) 31 days after ablation of the brachial nodes (bb′). Bottom picture: observation through ICG (injection in the back of the hand) of the functional lymphatic drainage pathways and of its collateral median pathway after brachial and axillary curettage directed toward the zone where nodal ablation was done (white arrow) 27 days after operation (cc′). The intensity of the blue coloration of the lymphatic vessels was enhanced to facilitate the observation of the lymphatic vessels’ course (c"), the original photograph is presented in Figure 10.
The second mappings also allowed to observe the presence (four cases on seven) of a collateral supplementary and functional drainage after ablation of the brachial nodes (Fig. 7).
Dissection
After euthanasia, the dissections made after injection with Evans Blue showed a certain type of substitution lymphatic pathways. Indeed, although no contralateral pathway was observed, superficial functional pathways were observed in all cases. These secondary superficial pathways during intervention joined the deep vessels of the thorax, thanks to perforating lymphatic vessels to assure the continuity of the lymphatic drainage of the foreleg.
During dissections, several cases were observed: either the presence of only one perforating vessel called “the main one” (Fig. 8) or the presence of other perforating vessels called “accessory,” allowing to increase the drainage possibilities of the “main perforating vessel.”
Dissection and observation after coloration with Evans Blue of the functional lymphatic vessels after surgery. Macroscopic global view of the functional lymphatic drainage
The absorption of Evans Blue by the lymphatic system after massage of the injection spot allows the observation of lymph collectors of the forearm corresponding to the “normal” anatomical descriptions of the lymphatic pathways draining the hand toward the shoulder (cf. supra). After their passage at the level of the elbow, these pathways will continue to follow a normal course: namely, they lateralize and run to the posterior part of the shoulder. In all cases, at the level of a precise point where the spinodeltoid muscles and the long and lateral portion of the triceps brachii meet (Figs. 8 and 9), a perforating vessel named “principal” is formed starting from the ascending vessels and then going deeper. Past this point, the ascending vessels will continue to follow a normal course toward the latissimus dorsi. Once arrived on the ablation spot of the brachial nodes, the lymphatic collectors are either totally occluded, with a clear migration stop of the dye (five cases on seven), or they continue their course and are thrown into the axillary cavity devoid of its nodes (two cases on seven).
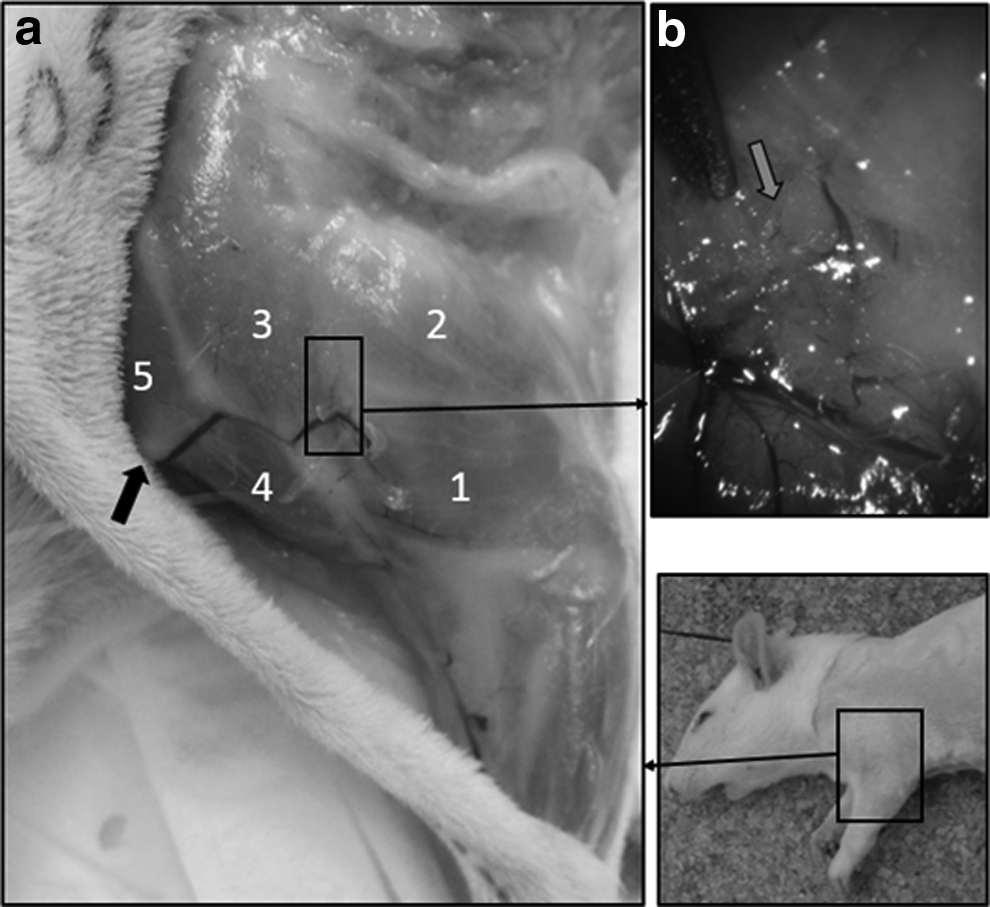
Localization of the origin of the “principal” perforating lymphatic vessel. Left: Macroscopic view of the operated foreleg with 1, lateral portion of the triceps brachii; 2, long portion of the triceps brachii; 3, spinodeltoid muscle; 4, acromial head of the deltoid and clavicular head of the deltoid, called also cleidobrachial muscle. 5, cleidobrachial muscle Black arrow: clavicle
In four on seven cases, this drainage pathway is completed by other perforating vessels that are formed starting from a pathway called “retrograde,” originating at the level of the ablation spot of the brachial nodes and assuring the continuity of the dye of the ascending lymphatic vessels arriving at the ablation spot. This pathway forms a retrograde hook going from the posterior part of the shoulder (nodal ablation spot) to the acromial head of the deltoid muscle, descending along the vein running through the spinodeltoid muscle before ramifying and giving birth to the axillo-brachial vein. At the level of the first one-fourth of the common border of the trapezius and spinodeltoid muscles (cervical head), the lymphatic flow goes deeper again through a first superior accessory channel. Then, this retrograde channel goes to the intersection of the deltoid (acromial head), cleidocephalicus, and spinodeltoid muscles, where it gives birth to another inferior accessory perforating vessel. On this very spot, the retrograde channel is joined by a (now) ascending collateral vessel coming from the “normal” ascending vessels. This collateral vessel ascends along the interaponeurotic space of the spinodeltoid and deltoid muscles (acromial head) and then continues toward the cleidocephalicus muscle. Then a complete collateral vascular loop with two drainage directions is formed from which “accessory” lymphatic perforating branches emerge (Fig. 10).

Macroscopic and microscopic views of the retrograde channel and of the collateral vascular loop with two drainage directions. Black and white checkered arrow: ablation zone of the brachial nodes. White arrow: drainage direction ascending to nodal ablation zone. Grey arrows: descending direction of the retrograde drainage vessel. Hatched arrow: drainage direction of the collateral ascending vessel. *Localization of the origin of the “principal” perforating vessel. **, ***Localizations of the origins of the other “accessory” perforating vessels.
The direction of the lymphatic flow of both the descendant and ascendant pathways of the drainage loop was observed through a vascular permeability test. The “empty-and-refill test” was adapted for the lymphatic circulation on dead animals. It consists in draining the dye from a part of the lymphatic vessel, since the latter cannot be filled because of the ceasing of lymphatic flow in the dead animal. The arrival of the dye in the drained spot is done manually by continuous soft slow and sliding pressure movements performed on the vessel above and then under the spot that has been previously drained. It is enough to observe the direction of the filling to deduce from this the direction of the physiological drainage.
In the two cases where the regenerated ascending lymphatic collectors continued their way to be thrown into the axillary cavity, accessory perforating vessels were observed. This observation leads to the supposition that the drainage offered by the regenerated vessels through the spot where the brachial nodes were ablated is not sufficient to assure a correct drainage and requires the supply of perforating vessels.
The same dissection type after injection with Evans Blue was done on the contralateral reference feet. Neither collateral pathways nor perforating vessels were ever observed.
Conclusions
Surgical technique
The posterior approach seems to be the most interesting because it is the least traumatizing. Indeed, if the animal is placed in supine position, with its foreleg held in abduction and anterior elevation, an anteromedial approach to arrive in the axillary cavity necessarily requires a dissection of the pectoral muscles. This anteromedial surgical approach turns out to be more traumatizing and does not allow the successive ablation of the brachial and axillary nodes in only one approach channel.
The resection of the brachial and axillary nodes in the rat through the posterior approach, as proposed in this study, does not require a preliminary observation of the nodes through coloration. This posterior approach allows the resection of nodes without major difficulty, despite the dissection of the axillary nodes in contact with the blood vessels and the nerves over the axillary cavities. No death or muscular paralysis was caused on whatever animal that had been operated during this study and its preliminary experiments.
During nodes dissection, two to four brachial nodes and three constant axillary nodes were resected (seven on seven). The amount and the morphology of the ablated nodes in this study are different from the data in the literature. The number of brachial nodes in the rat can vary from two to seven nodes. 13 In the anatomical description of rat's lymph nodes made by Tilney, 10 these brachial nodes are constantly presented in groups of three, whereas the axillary nodes are presented in groups of four. Concerning the axillary nodes, Job 9 described them as three distinct entities, each one composed of two joint nodes.
In our sample of 14 feet, which had been examined for this study, the presence of the “elbow” node has never been observed.
Lymphographies with ICG and terminal dissections after coloration with Evans Blue
The technique of vascular observation “in vivo” after subcutaneous injection of ICG has been chosen to map the superficial lymphatic vessels without euthanizing the animals. This technique of observing the superficial lymphatic vessels in humans 14 is more and more adopted in fundamental research on laboratory animals.
Indeed, it is used to observe the normal lymphatic pathways, but allows to detect the anatomical modifications of these vessels after alteration/perturbation of the lymphatic pathways. 8
Once injected, the ICG is fixed particularly on the proteins: endogenous lipoproteins present in the interstitial sector. These macromolecules that just have been “marked” by the ICG molecules will then be taken over by the lymphatic system. This ICG molecule is not only fixed on endogenous macromolecules but also on phospholipids. 15 For this reason, the tracer binds to the membrane macromolecules, and stays longer the lymphatic vessels. This represents an added value compared with classic Evans Blue dye mapping.
To observe this tracer and to allow the detection of subcutaneous lymphatic vessels, this tracer “bound to the macromolecules” has to be excited with a skin perforating light of the near infrared type (760 nm). The bound and excited tracer on its turn emits a radiation of which the peak emission is also within the near infrared (830 nm), which gives it the property of also perforating the skin. This emitted beam is detectable only with the use of a specific camera provided with adequate filters and sensitive to invisible near infrared radiation. 16
The medical equipment used nowadays in this field is not fully satisfactory for image quality. Indeed, the technologies do not allow obtaining sharp images at whatever acquisition distance, which represents a negative element if you want to obtain a global mapping of an important skin zone, especially in human or in the biggest laboratory animals. This last element makes it necessary to superpose images to reconstruct the global image of the zone to be mapped. 14
However, this technique of observing lymphatic vessels is very promising for indirect nonionizing investigation (contrarily to lymphoscintigraphy) of the anatomy and the lymphatic physiology of the superficial net.17,18
Nevertheless, it is important to obtain more scientific data for the validation of these techniques to obtain quantitative data concerning physiology, namely, the lymph transport parameters in the vessels. Indeed, in rats, a certain influence of the injected tracer on the lymph transport has been shown, more exactly on the contractility of the vessels. 19
This last point by no means affects the advantages of using this tracer to obtain indirect “in vivo” lymphographies in laboratory animals, but justifies the use of massaging the injection sites to facilitate the migration of the tracer in the vessels.
The main limitations of this observation technique are the fact of obtaining generally a black-and-white image nuanced by the presence of gray with an average image quality, but also its restrictive application to the superficial lymphatic system if the investigation is done in “in vivo” conditions without cutaneous dissection.
Indeed, during the “in vivo” mappings done for this study, the perforating vessels allowing the lymph transport continuity from the periphery to the depth have not been observed. However, these mappings with ICG allowed, without euthanizing, the detection of the superficial functional vessels after operation, but in no way the anatomic explanation of how the deep drainage was done. Different investigation delays have been chosen in function of the different phases of the regeneration process of the lymphatic vessels after surgical interruptions in the laboratory animal. 20 The authors thought it was interesting to realize “in vivo” lymphographies before and during the functional regeneration phase of the lymphatic vessels (two investigations on seven) at 7 and 15 days. The latter took place on average in 14 to 21 days after the intervention21,22 and especially after this phase in 30 days and more after surgery (five cases on seven).
For the already mentioned reasons (cf. limitations of the use of ICG), it has been indispensable, at the end of the experiment, to dissect the animals completely after coloration of the lymphatic network with Evans Blue. 8 The Evans Blue that has been subcutaneously injected is also fixed on the endogenous proteins of the interstitial sector. It is then taken over and transported by the lymphatic system. 23
The advantage of using a simple dye such as Evans Blue is the possibility to obtain, without any advanced technology, color images of very good quality, allowing the identification of anatomic structures of dissected tissues, which is much more difficult to perform with the method based on the bound and excited ICG near infrared detection.
In general, a 12 months period has been observed to follow up the volumes before the final dissections, to control the very slow and late appearance of the volumetric changes of the operated members. 7
The lymphatic substitution pathways
The presence of perforating vessels allowing a drainage from the surface into depth after brachial and AND in rats has never been described in the literature until now. It explains why no secondary lymphedema of the operated foreleg has ever been detected during these 12 months of follow-up. The appearance of these particular substitution pathways connecting the superficial and deeper networks also justifies the necessity of using techniques (physical or chemical) that can be helpful to nodal ablation to produce a chronic lymphedema model in rats.5–8
In humans, these perforating vessels are well described theoretically on the numerous anatomical schemes. 24 However, the in vivo observation of the functional perforating vessels of the superior limb is not really obvious. Indeed, they have been observed in a very inconstant manner during autopsy of fetuses. 25 In 2007, an autopsy on a human body of a superior limb of a patient who had been operated for breast cancer with axillary ablation allowed observation in several places these perforating vessels, although they had not been observed on the not operated second upper limb. 26 All this allows us to believe that the perforating vessels, which certainly are present at birth but mostly not functional in a normal situation, offer some not negligible derivation possibilities for lymph drainage apart from the supply pathways already described 25 in case of failure of the lymphatic system of the superior limb after oncological treatment of breast cancer. This last hypothesis is based on the results of this study on laboratory animals after ablation of the axillary lymphatic and brachial nodes.
To understand and prevent the appearance of secondary lymphedema of the superior limb in women having an important AND, it is indispensable to study on laboratory animals the impact of this type of surgery. The purpose of this research was to study on rats how the lymphatic system reacts to a complete nodes dissection of the foreleg. To do so, a new surgical approach has been tested successfully, allowing the resection of the brachial and axillary nodes using only the posterior approach channel. The observation of the functional superficial lymphatic pathways was done by indirect lymphography with ICG under general anesthesia at different postoperative times (until 60 days). The use of this technique has the advantage of allowing to map the superficial lymphatic vessels without euthanizing the animals. After an important follow-up period, the final dissections after indirect coloration with Evans Blue of the lymphatic vessels allowed to discover the defense mechanism of the lymphatic system against this traumatic surgical aggression. In rats, after total nodal ablation of the foreleg, an acute postoperative edema appears, but generally does not last longer than 21 days. Therefore, particular substitution pathways, namely, perforating vessels, allow a deep lymph drainage short circuiting the nodal resection zone.
The observation and anatomic description of this type of perforating vessels after nodal resection is a major asset in fundamental research. To researchers it offers the possibility to consider a future surgical neutralization of these perforating vessels together with brachial and axillary curettage to create, without the auxiliary use of inflammatory agents, an animal chronic lymphedema model. This model is indispensable to understand the biochemical and tissue mechanisms of the formation of secondary lymphedemas in nodal ablations.
Footnotes
Acknowledgments
The authors would like to thank the European Association for Andullation Therapy for its support during this study as well as the company Pulsion Medical Benelux for lending medical equipment.
Author Disclosure Statement
No competing financial interests exist.