
Abstract
Abstract
Background:
Skin fibrosis is a clinically serious pathological process of secondary lymphedema (SLE). Previous studies have shown that mast cells (MCs) are involved in lymphedema (LE) and play a key role in the pathological process of skin fibrosis. However, the role of the protease chymase and transforming growth factor-β1 (TGF-β1) secreted by MCs in the fibrotic skins of patients with secondary lower limb LE has not been explored.
Methods and Results:
In this study, full-thickness skin biopsies of lymphedematous limbs from seven SLE patients and control samples from seven healthy controls were harvested. The skin samples were assayed by Masson, immunohistochemical, and immunofluorescence staining and were analyzed by western blot and enzyme-linked immunosorbent assay. The number of MCs and the expression of proteases, TGF-β1, and latency-associated peptide TGF-β1 (LAP TGF-β1) were analyzed. The number of MCs and the expression of chymase, TGF-β1, and LAP TGF-β1 were increased in fibrotic skin compared with normal skin. The increased expression of TGF-β1 on lymphatic vessels, endothelial cells, and in skin interstitial tissues overlapped with chymase expression.
Conclusions:
Our results demonstrate that chymase and TGF-β1 expression was significantly increased in the fibrotic skin of secondary lower limb LE. The increased expression of chymase in the skin may play an important role in the development fibrosis in the lymphedematous skin. We speculate that chymase may facilitate the release of LAP TGF-β1 to generate activated TGF-β1, and the upregulation of active TGF-β1 can promote fibrosis in the SLE skin.
Introduction
L
Previous studies have shown that mast cells (MCs) are involved in LE and play a key role in the pathological process of skin fibrosis. 7 Human MCs, which are derived from hematopoietic stem cells and are widely distributed throughout the skin, liver, heart, lungs, and other organs, were first found and described by Paul Ehrileh in 1877.8,9 MCs play an important role in the physiological process of tissue fibrosis.10,11 Studies have shown that the number of MCs is significantly increased in fibrotic tissues, and that these MCs secrete large amounts of chymase, which is a chymotrypsin serine protease, stored in the secretory granules of the MCs. The amount of secreted chymase positively correlates with the degree of fibrosis in the affected area.12–16 Further research found that chymase can active transforming growth factor-β1 (TGF-β1) to trigger the inflammatory process of tissue fibrosis. 17 TGF-β1 is the most prevalent isoform of the TGF-β family of ligands and can be released by platelets, endothelial cells, hematopoietic cells, macrophages, and fibroblasts. 18 TGF-β1 has a wide range of biologic activities and functions, in particular, to promote inflammation during the pathological processes of fibrosis in many organs.19–23 Therefore, the increased levels of chymase secreted by infiltrating MCs may indirectly cause tissue fibrosis by activating TGF-β1.
Little is known about the expression of chymase in the skin of patients with secondary lower limb LE. We hypothesized that the process of SLE fibrosis may be closely related with the amount of secreted chymase according to previously highlighted studies. We sought to determine if increased levels of MC-secreted chymase promoted the activation of TGF-β1, in areas of skin fibrosis in SLE limbs. Therefore, we examined the expression levels of chymase and TGF-β1 in the skin of patients with secondary lower limb LE. The results provide new insights for understanding the pathophysiological mechanism of SLE fibrosis and identify new potential therapeutic targets.
Materials and Methods
Patients and tissue sample collection
Seven female patients (average age 51.4 years, range 42–59 years) with secondary lower limb LE were enrolled in this study from July to November 2014. All patients had undergone surgery, radiotherapy, and chemotherapy for endometrial cancer or cervical cancer. The average duration of SLE was 6.6 years at stages II–III. 24 Full-thickness skin biopsies of lymphedematous limbs (2.5 × 1.5 cm) were taken from the pretibial-medial leg, 5 cm above the ankle, from each patient. Control biopsies were obtained from seven healthy female volunteers (average age 48.6 years, range 39–54 years). All subjects provided informed consent. This clinical study was approved by the Ethics Committee of the Shanghai Ninth People's Hospital.
Masson staining
Skin biopsies were embedded in paraffin, and the 5 μm sample sections were deparaffinized, rehydrated, and then stained in hematoxylin solution. Excess dye was removed with hydrochloric acid in ethanol and the sections were stained in Ponceau-acid fuchsin solution, immersed in 2% glacial acetic acid, incubated in phosphomolybdic/phosphotungstic acid solution, and transferred directly to aniline blue solution. Before imaging, the sections were washed in 2% acetic acid. Collagen and mucous cells appear blue; cytoplasm, cellulose and muscle cells are stained red; cell nuclei are dark blue.
Immunohistochemical staining
Tissue sections were deparaffinized and hydrated. Pretreatment for antigen retrieval was done at 96°C in 0.01 M sodium citrate buffer solution (pH 6.2) for 20 minutes. After blocking for 30 minutes with 5% bovine serum albumin (BSA), samples were incubated with primary antibodies at 4°C overnight: rabbit anti-Mast Cell Chymase antibody (1:500; Abcam, Cambridge, United Kingdom), rabbit anti-TGF-β1 antibody (1:50; Abcam), and goat anti-LAP TGF-β1 antibody (1:100; R&D, Minneapolis, MN). After overnight incubation, sections were incubated with the relevant secondary antibodies (Envision+ system +HRP; DAKO, Glostrup, DK). Sections were imaged with a Nikon 90i light microscope (Nikon, Tokyo, Japan). Ten visual fields magnified 200 times for each sample were randomly selected in the lymphedema and control groups to observe the immunopositive expression. We counted the number of chymase-positive MCs and LAP TGF-β1-positive granules, the total number of cells, the areas of TGF- β1 expression and determined the integrated optical density (IOD) to calculate the chymase-positive MC ratio; the amount of chymase-positive MCs and LAP TGF-β1 per mm2 of skin was quantified using Photoshop CS3 and Plus Image-Pro 6 software.
Immunofluorescence staining
Paraffin tissue sections were deparaffinized and hydrated. Pretreatment for antigen retrieval was done for 25 minutes at 96°C in 0.01 M sodium citrate buffer solution (pH 6.2). The sections were rinsed in 0.2% tritonX-100 for 5 minutes, blocked for 30 minutes with 5% BSA, and then incubated at 4°C overnight with mixtures of two primary antibodies: mouse anti-Mast Cell Tryptase antibody (1:100; Abcam), rabbit anti-Mast Cell Chymase antibody (1:100; Abcam), rabbit anti-TGF β1 antibody (1:50; Abcam), mouse anti-TGF-β1 antibody (1:200; Abcam), and mouse anti-podoplanin antibody (1:50; AngioBio, San Diego, CA). After overnight incubation, sections were incubated in the dark for 2 hours at 37°C with secondary antibodies, Alexa fluor 555 goat anti-mouse and Alexa fluor 488 goat anti-rabbit (1:300; Invitrogen, Carlsbad, CA). After counterstaining with 4′,6-diamidino-2-phenylindole (KeyGen Biotech, Nanjing, China), the sections were covered with a fluorescent mounting medium. Fluorescence microscopy was performed using an LSM 710 confocal laser scanning microscope (Carl Zeiss, Jena, DE), and digital pictures were taken with Zen 2010 imaging software (Carl Zeiss). Ten visual fields magnified 200 times for each sample were randomly selected for analysis; the number of MCs was counted and Photoshop CS3 and Plus Image-Pro 6 were used for quantification.
Western blot analysis
Lysates from the skin samples were extracted and centrifuged at 12,000 g for 10 minutes at 4°C. Protein concentrations in the supernatant from each sample were determined. Equal amounts of protein from each sample were resolved by sodium dodecyl sulfate–polyacrylamide gel electrophoresis and transferred onto polyvinylidene fluoride membranes. Membranes were incubated overnight at 4°C with the primary antibody following block solution treatment. The primary antibodies were rabbit anti-Mast Cell Chymase antibody (1:1000; Abcam) and mouse anti-TGFβ1 antibody (1:2000; Abcam). Following incubation, the membranes were washed with Tris-Buffered Saline Tween-20(TBST), incubated for 2 hours with the appropriate secondary antibody anti-mouse or anti-rabbit (1:5000; Daixuan Company, Shanghai, China), and then developed with the ECL kit (Pierce ECL 32106; Thermo Fisher, New York, NY) according to the protocol. The relative intensity of the immunoreactive bands was quantified using the Image-Pro Plus6 software.
Enzyme-linked immunosorbent assay
Each skin sample was weighed, cut up, and homogenized in cold phosphate-buffered saline containing protease inhibitors. The slurry was drawn into an anticoagulant tube, centrifuged at 5000 g for 5 minutes at 4°C, and the supernatant was collected. Enzyme-linked immunosorbent assay (ELISA) kits for human chymase (SEG515Hu, USCNLIFE, Wuhan, China), TGF-β1 (88–8350; eBioscience, San Diego, CA), and LAP TGF-β1 (88–50390, eBioscience) were used according to the manufacturer's introductions. From each skin sample, 100 μL of supernatant was used per well, readings were taken at 450 nm, and the data were analyzed using a microplate ELISA machine (DNM-960PERLONG 2; Perlong, Nanjing, China).
Statistical analysis
Data are presented as mean (range) or number (percent). Statistical analysis was performed using t-test with SPSS 17.0 software (IBM, Armonk, NY). All tests were two sided, with a significance level of 0.05 or 0.01.
Results
Elevated expression of collagen in the lymphedematous skins
Increased collagen deposition was observed in lymphedematous skins by Masson staining compared with the control skins, indicating severe and obvious fibrosis in the LE (Fig. 1).

Images of collagen in Masson-stained skin sections. The staining of collagen (black arrows).
Increased numbers of MCs and expression of proteases in the skin of SLE
The number of MCs was significantly enhanced in lymphedematous skin compared with normal skin. Two proteases secreted by MCs: chymase and tryptase were found in the cytoplasm of MCs as well as in the interstitium. Two subtypes of MCs were distinguished—MCT and MCTC—the former containing tryptase (T) and the latter containing tryptase and chymase (TC). The MCT expression ratio (the numbers of MCT/total numbers of cells) was 0.0407 ± 0.03, and the MCTC expression ratio (the numbers of MCTC/total numbers of cells) was 0.1513 ± 0.06 in the lymphedema group, which was higher than the control group (0.0242 ± 0.02 and 0.0645 ± 0.04, respectively; p < 0.01). We also observed that the amount of MCTC was more than MCT. This confirmed previous studies which showed that MCTC was the primary MC type in the skin and that these two proteases could be simultaneously released into tissues by MCs (Fig. 2). 25
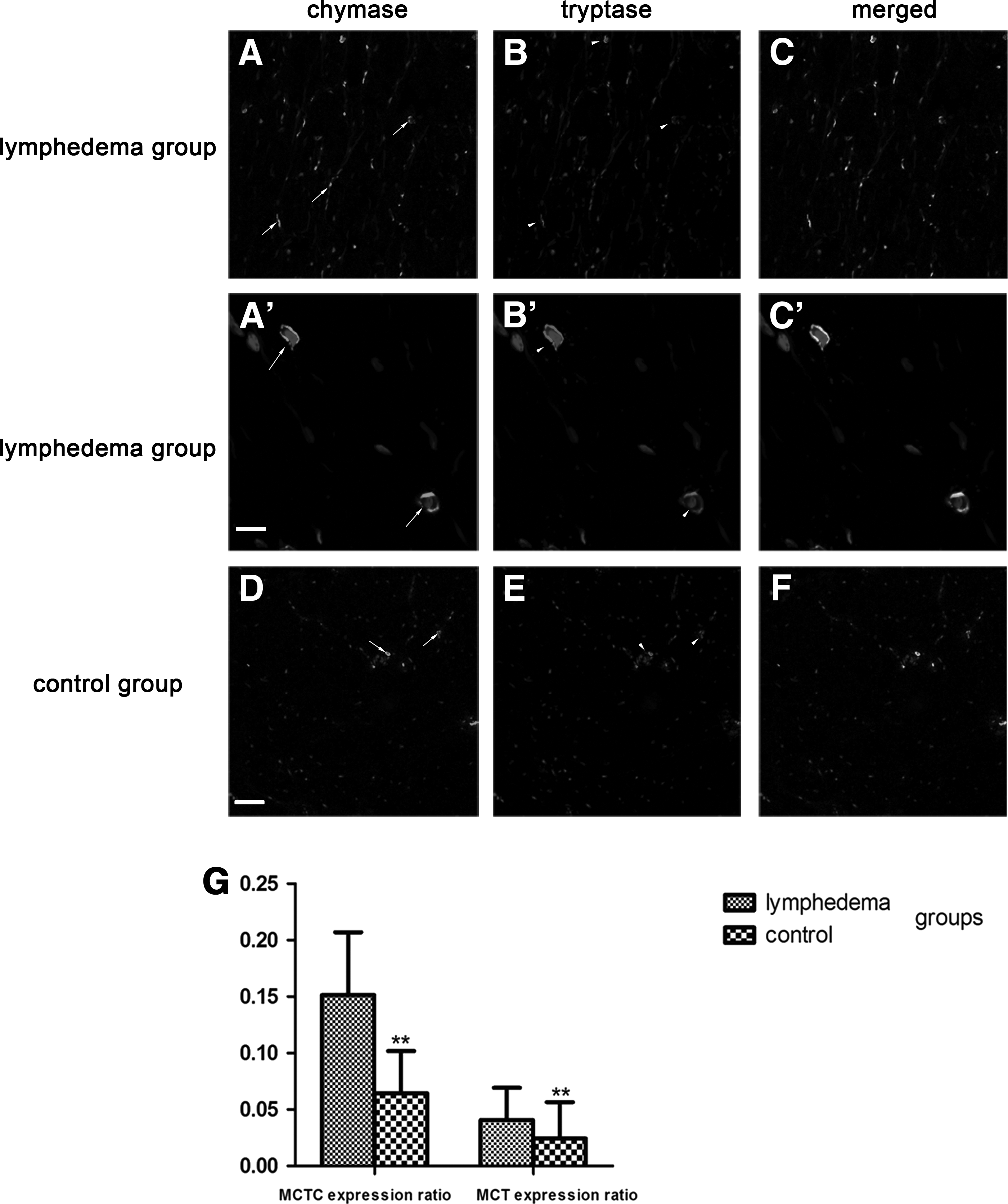
Increased chymase expression in the skin of secondary lower limb LE
Chymase expression showed an increasing trend in the skins of SLE. A large number of chymase-positive MCs were diffusely distributed in the dermal papilla and reticular layers, and aggregated near blood vessels, sweat glands, and hair follicles. The total amount of chymase in the lymphedema group was greater than the control group (Fig. 3A, B). The ratio of chymase-positive MCs (the numbers of MCs containing chymase/total numbers of cells) was 0.1381 ± 0.04 in the lymphedema group, which was higher than in the control group, at 0.0632 ± 0.17 (p < 0.01). The number of chymase-positive MCs per mm2 of skin in the lymphedema group was 164.45 ± 58.38, and 75.12 ± 24.04 in the control group, which was significantly different (p < 0.01) (Fig. 3C, D). These results were confirmed by western blot analysis, in which the lymphedema group had significantly higher levels of chymase than the control group (p < 0.01) (Fig. 4), and by ELISA, where the chymase content in the lymphedema group was 193.14 ± 48.42 ng/g, significantly higher than the control group (97.57 ± 17.86 ng/g; p < 0.01) (Fig. 5).
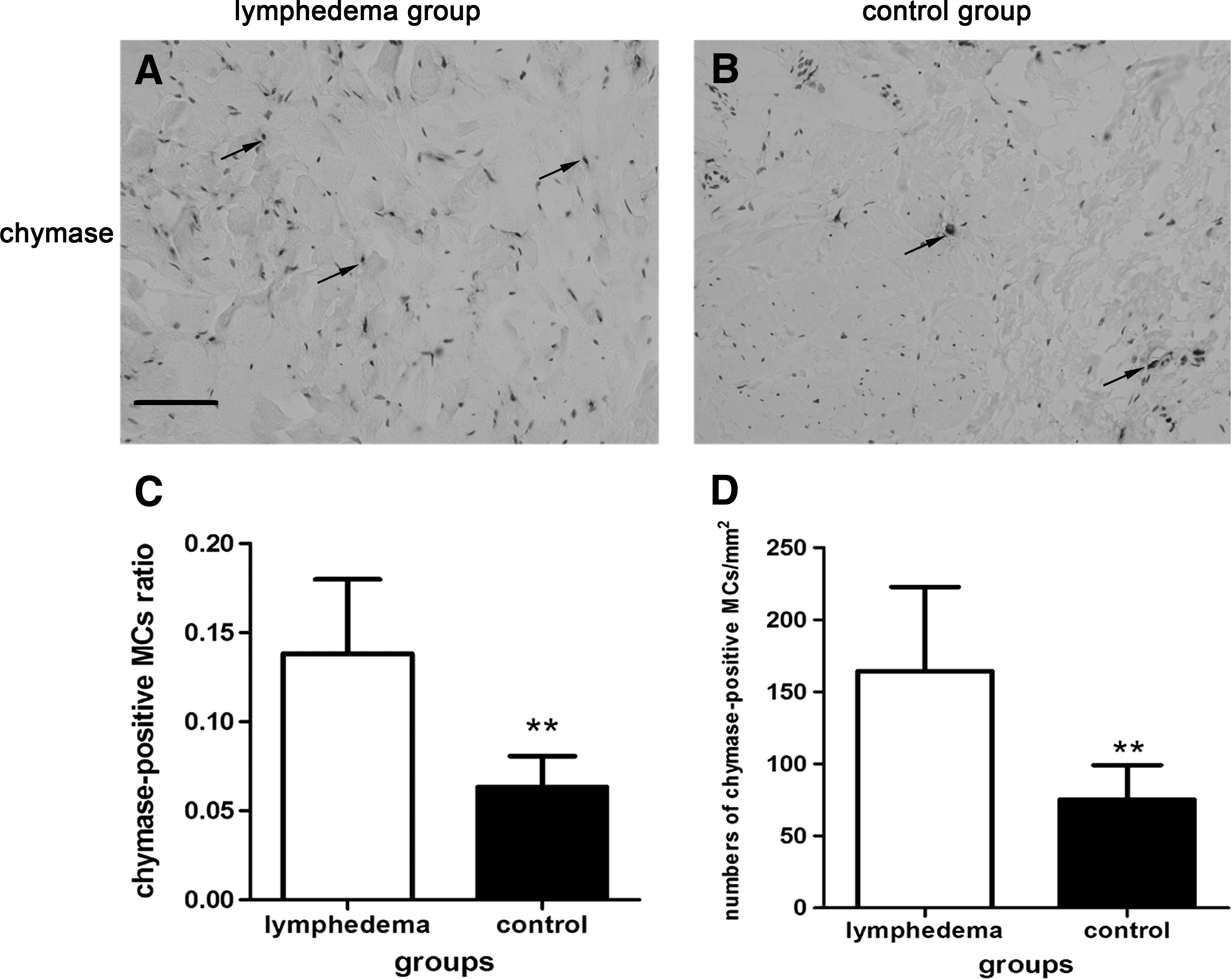
Immunohistochemistry-stained images of chymase (black arrows) and cell nucleus in lymphedematous skin
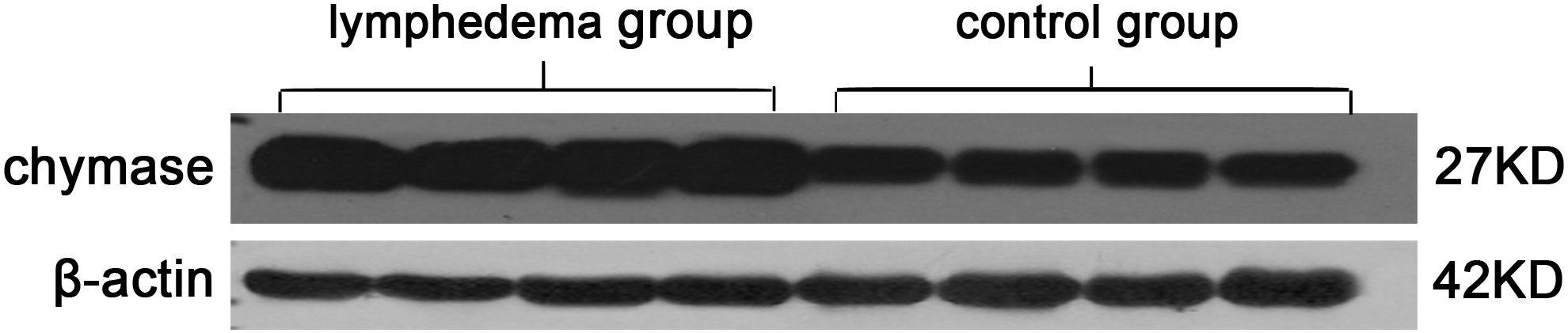
Western blot analysis of chymase in the skins of lymphedema group and control group. The concentration of chymase in the lymphedema group was higher than in the control group.
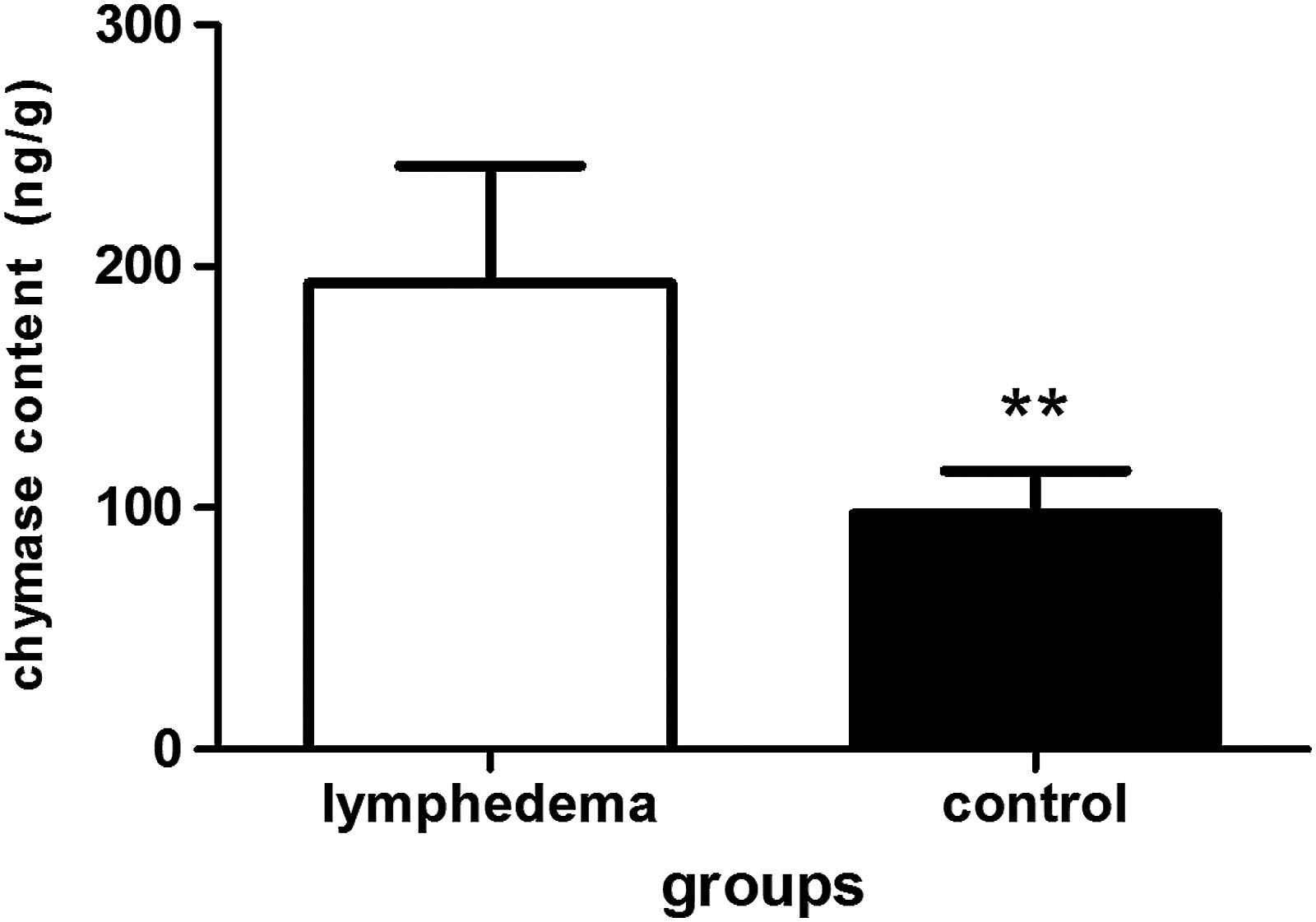
ELISA of chymase in the skins of the lymphedema group and control group. The lymphedema group had higher chymase concentrations than control group. Data are presented as mean ± SD, **p < 0.01.
Increased expression of both TGF-β1 and LAP TGF-β1 in the skin of secondary lower limb LE
TGF-β1was expressed predominantly near blood vessels, lymphatic vessels, hair follicles, and sweat glands (Fig. 6A–D). The region of TGF-β1 expression was 169,112.63 ± 150,801.43 and the IOD was14,055.4323 ± 10,801.15 in the lymphedema group and 63,921.85 ± 68,981.64 and 5518.275 ± 6151.42, respectively, in the control group. The differences between the two groups were significant (p < 0.01). The mean number of LAP TGF-β1 per mm2 of skin in the lymphedema group was 1439.46 ± 663.91, significantly higher than the control group (1020.90 ± 170.30; p < 0.01) (Fig. 6E–G). On western blot analysis, the concentration of TGF-β1 in the lymphedema group was higher than in the control group (Fig. 7). By ELISA, the content of TGF-β1 in the lymphedema group was 95.48 ± 52.98 ng/g, significantly higher than the control group (28.47 ± 25.15 ng/g; p < 0.05). The content of LAP TGF-β1 in the lymphedema group was 2145.29 ± 796.72 ng/g and 677.7 ± 215.22 ng/g in the control group, a statistically significant difference (p < 0.05) (Fig. 8).
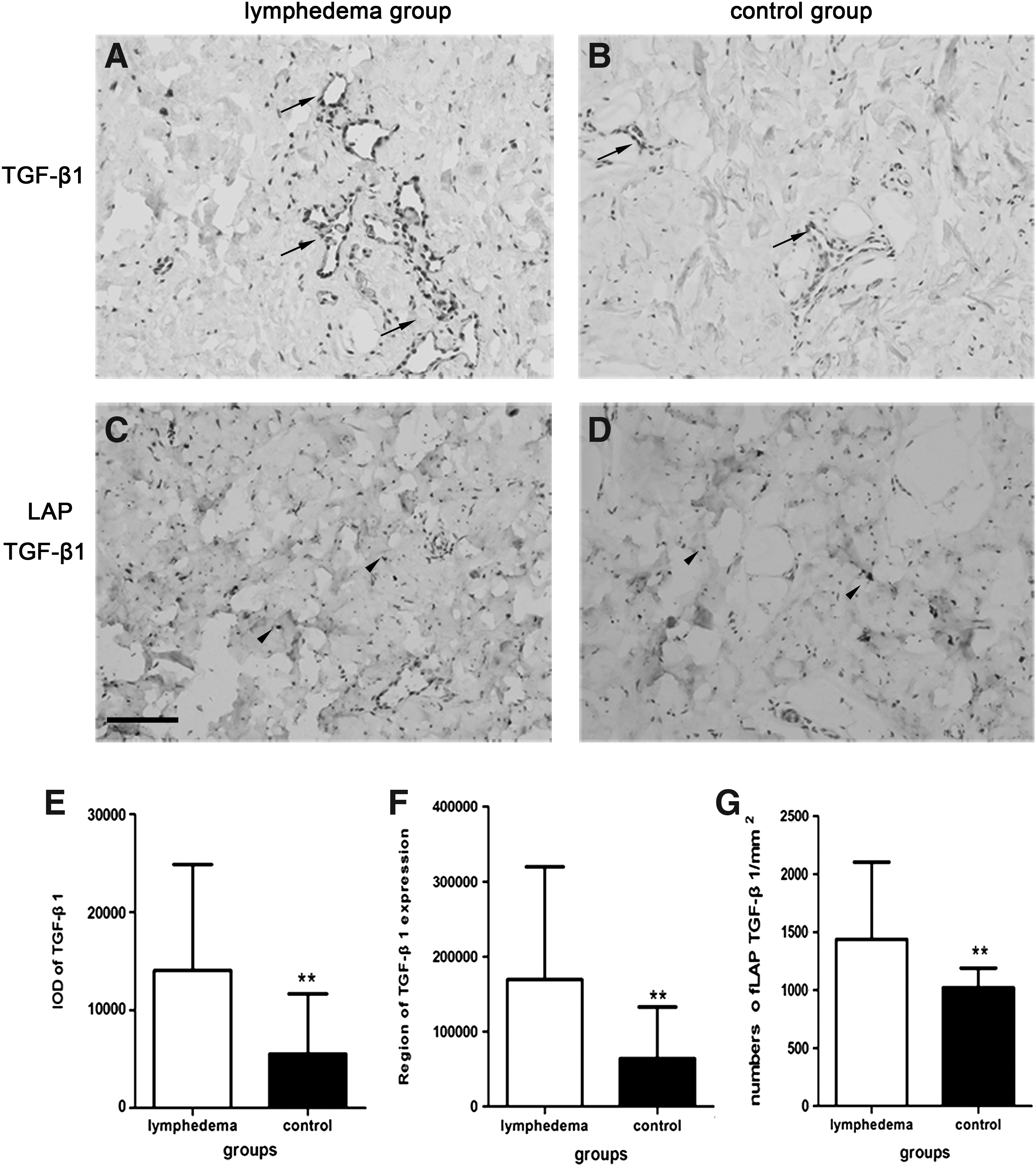
Immunohistochemistry-stained images of TGF-β1 (black arrows) and cell nucleus in lymphedematous skin
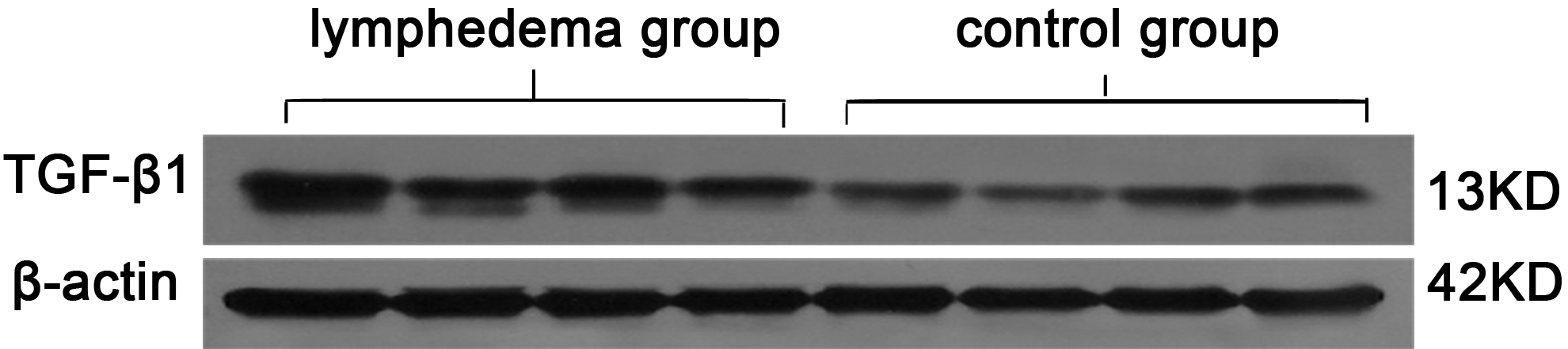
Western blot analysis of TGF-β1 in skins of lymphedema group and control group. The concentration of TGF-β1 in the lymphedema group was higher than in the control group.
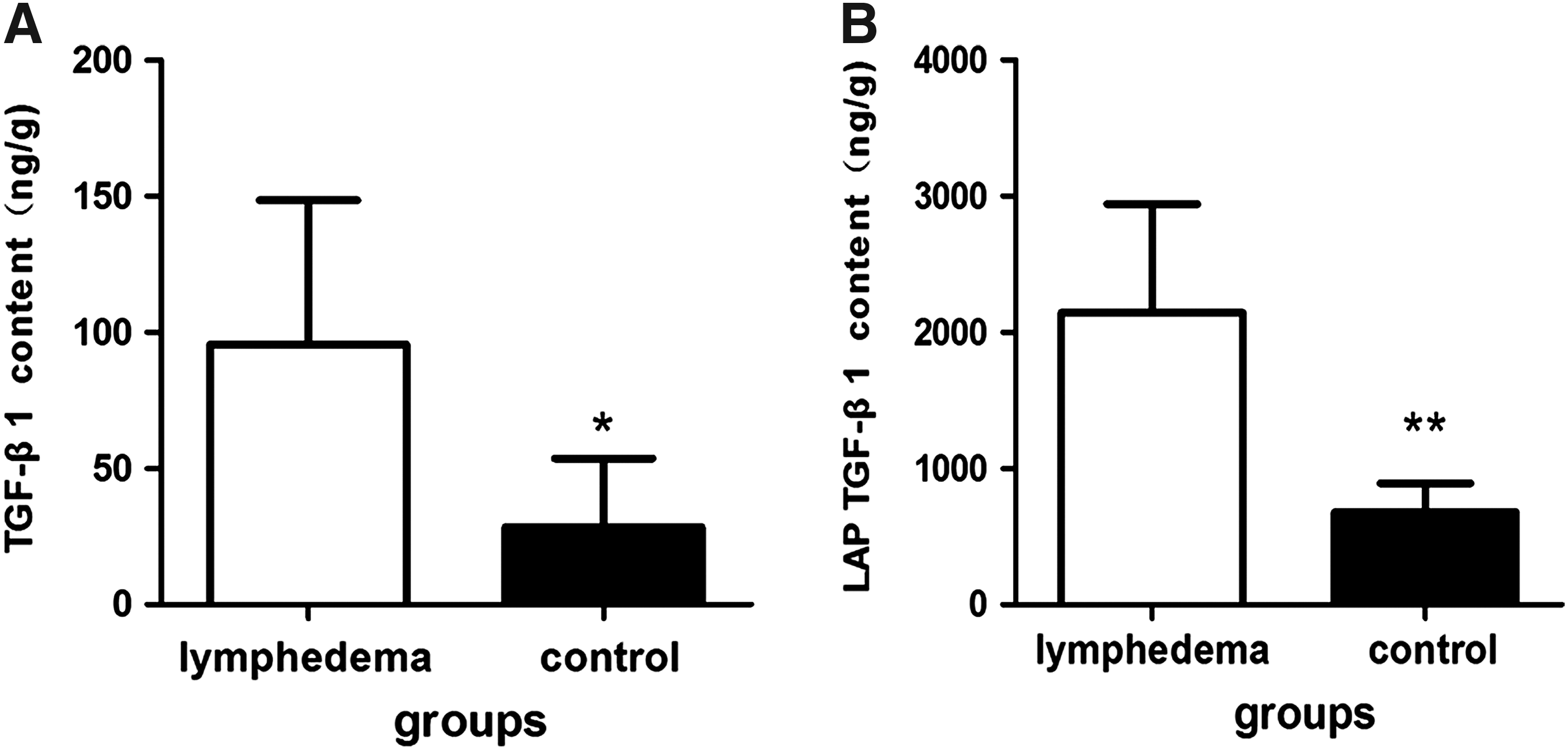
ELISA of TGF-β1
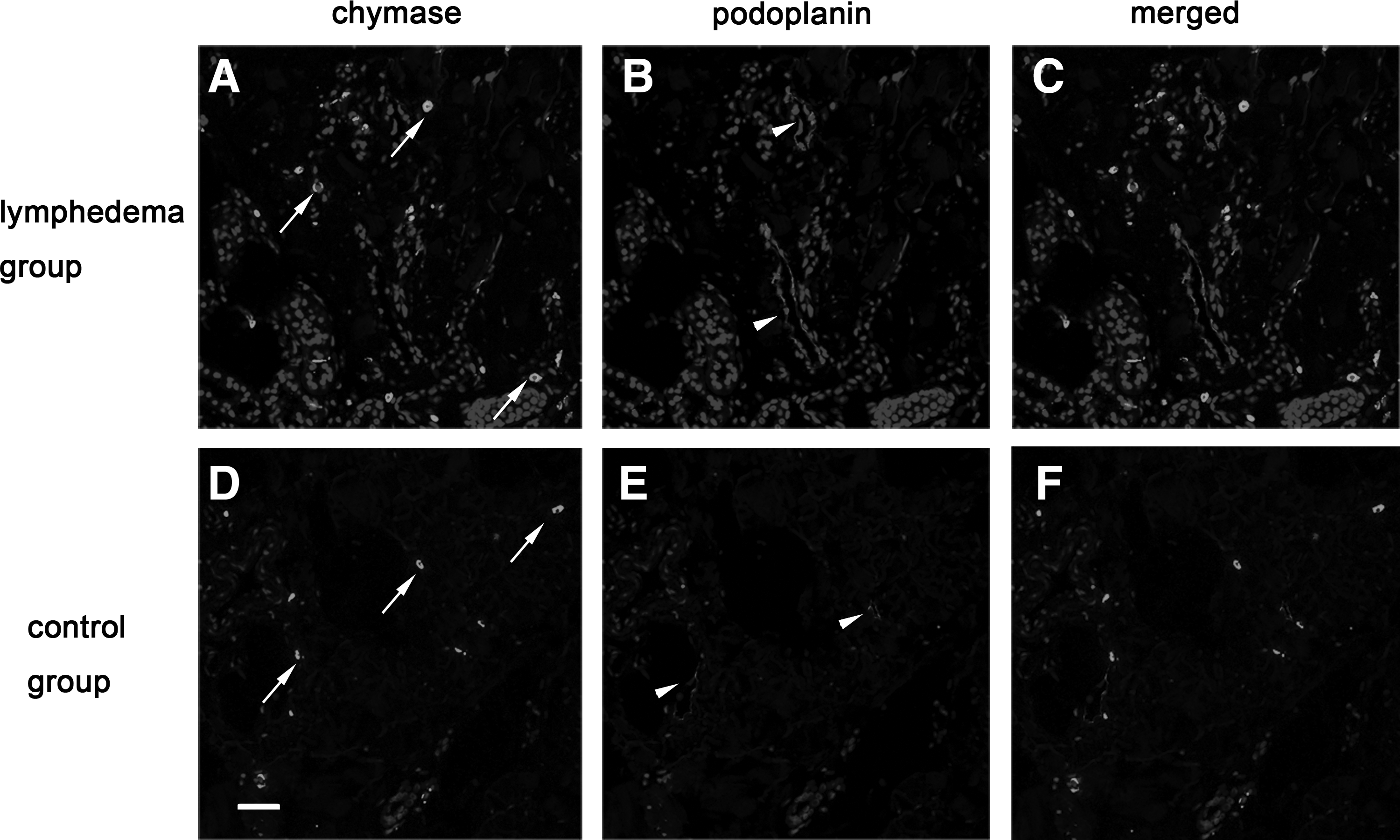
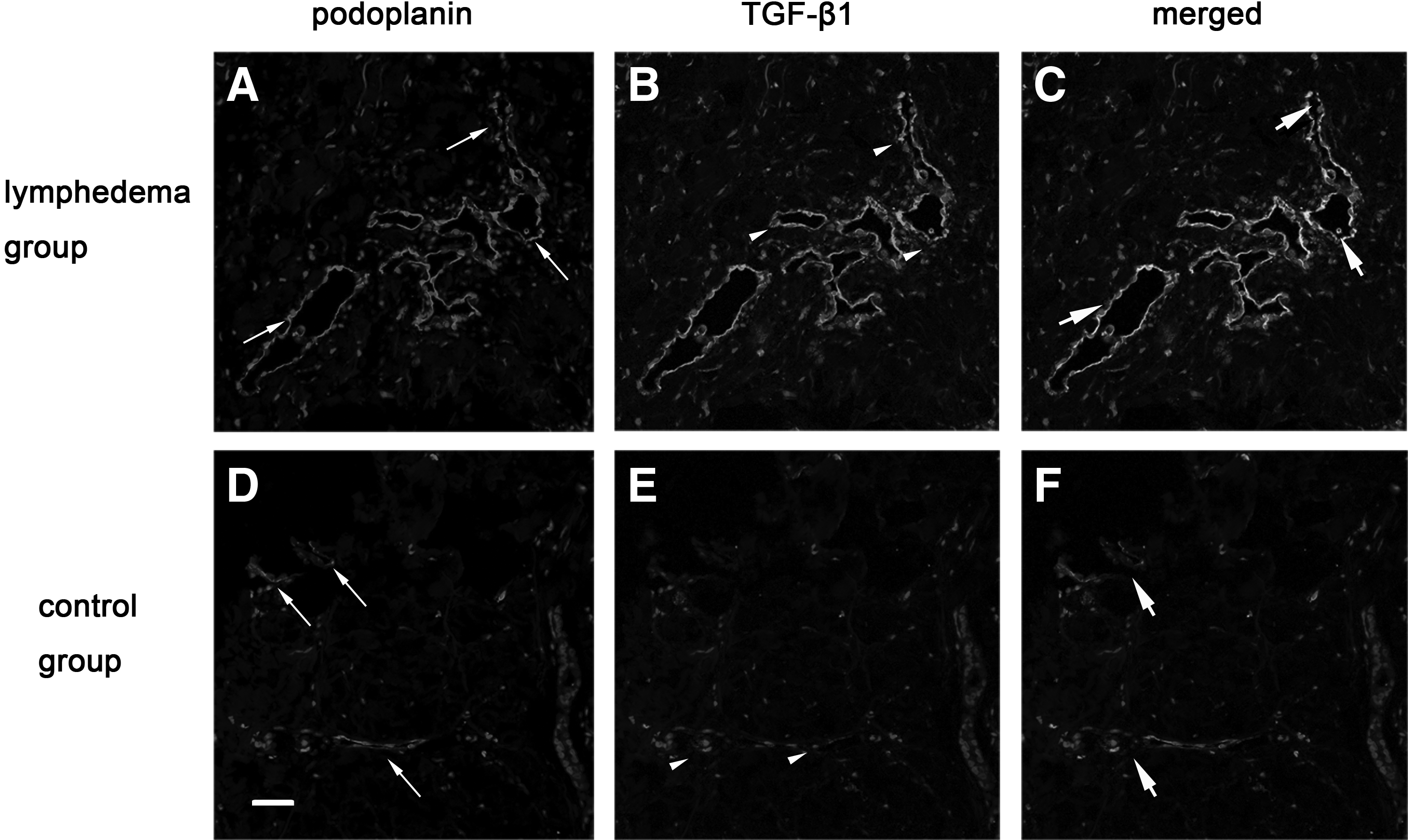
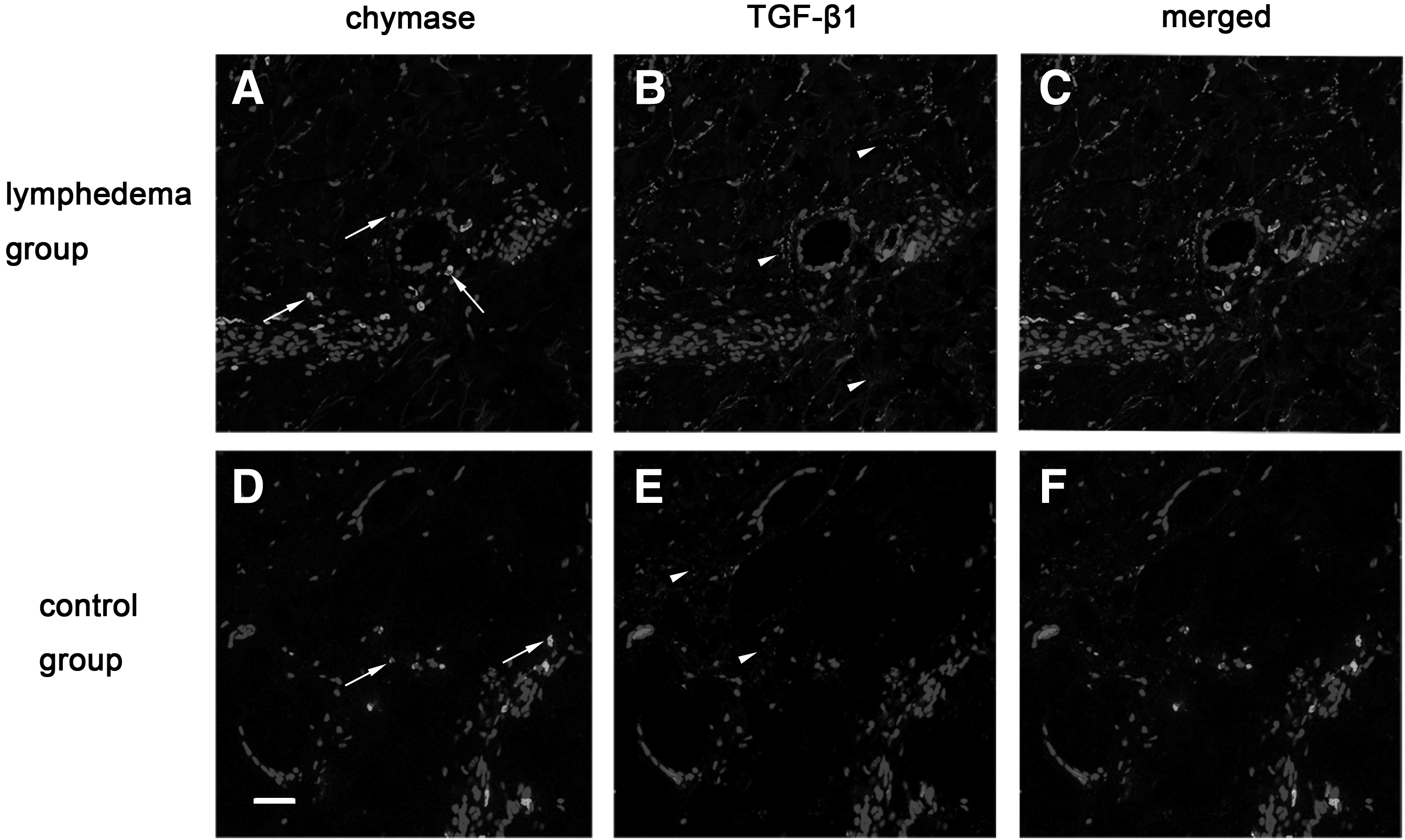
The increased expression of TGF-β1 on lymphatic vessels, endothelial cells, and interstitial tissues coincided with chymase expression
The relationship between chymase and TGF-β1 was explored because they were both overexpressed in SLE skins. The immunofluorescence staining showed that the lumens of lymphatic vessels were dilated and that more chymase aggregated near these abnormal lymphatic vessels. The amount of chymase in the lymphedema group was found to be increased compared with the control group by immunofluorescence double staining. Importantly, we observed that the increased expression of TGF-β1 was on lymphatic vessels and in interstitial tissues where a lot of chymase was recruited. To further observe, we found that TGF-β1 expressed on lymphatic endothelial cells, and this result was rarely reported by other scholars. On the contrary, in the control group, the lumens of lymphatic vessels were smaller and the lymphatic walls were smooth, and there were few chymase and TGF-β1 aggregations in the interstitial tissues or on lymphatic endothelial cells (Figs. 9–11).
(
(
(
Discussion
Our results demonstrate that chymase and TGF-β1 expression was significantly increased in the fibrotic skin of secondary lower limb LE. The increased levels of chymase in the skin may play an important role in the development fibrosis in the lymphedematous tissue. The increased expression of chymase may facilitate the release of LAP TGF-β1 to become activated TGF-β1. The upregulation of TGF-β1 can promote the process of fibrosis in the SLE skin.
Secondary lower limb LE is always accompanied by skin thickening, hyperpigmentation, and tissue fibrosis. 26 In the specimens taken from the limbs with stage II or III LE, we observed that a large number of MCs had been recruited to the thickened dermis, especially near blood vessels and dilated lymphatic vessels. Normally, there are few MCs in the upper dermis of skin, but in the lymphedematous skin, the numbers of MCs had increased. A previous study has shown that MCs promote skin fibrosis in the mouse through the production of multifunctional cytokines and proteases. 27 We speculate that MCs have similar functions in human lymphedematous skin, in particular when fluids accumulate in the SLE skin so that infiltrating microbes or specific antigens cannot be effectively eliminated, which will induce IgE-mediated hypersensitivity reactions. MCs localize around the blood vessels and lymphatic vessels that facilitate the binding of IgE antibodies via FceRI receptors to the surfaces of MCs. 28 This activates MCs, which can proliferate and secrete a whole variety of proteases, cytokines, and other mediators of inflammation. These products participate in a complex network of events that ultimately trigger collagen formation from type I procollagen, promote fibroblasts to proliferate, and cause local tissue remodeling responses.29–31
MCs secrete two important proteases, chymase and tryptase, and can be classified into two subtypes, MCT and MCTC, based on the enzymes it secretes. 32 In SLE skin, we found that the levels of tryptase and chymase were significantly increased compared with normal skin. It is interesting to note that both MCT and MCTC can secrete tryptase, but only MCTC release chymase, and there were greater numbers of MCTC than MCT in both the skin of SLE patients and controls. Although there is less overall chymase than tryptase in the skin tissue, previous work has demonstrated that chymase promotes tissue fibrosis. Katsuaki and colleagues measured and compared chymase expression in liver tissue from patients with primary biliary cirrhosis and autoimmune hepatitis, finding a positive correlation between the degree of liver fibrosis and chymase expression. 33 Based on the results of this study, we also believe that high expression of chymase is a critical factor in promoting the development of SLE skin fibrosis. Haimanot et al. suggested that increased chymase expression in the renal interstitium in response to a primary disease can induce renal fibrosis by activating TGF-β and angiotensin II signaling. 34 In both in vivo and in vitro experiments, Orito and Djuro have confirmed that chymase can cause pulmonary fibrosis in lung tissue.35,36 Xiaolin Dong has also shown that the expression of chymase in the keloid could promote the proliferation of fibroblasts by activating TGF-β1. 37
Many correlative studies have shown that TGF-β1 is involved in the pathological processes of organ fibrosis; TGF-β1 overproduction plays a crucial role in lung fibrosis, liver cirrhosis, cardiac fibrosis after infarction and skin scars, and thus, it is considered to be the most pivotal cytokine of induced fibrosis.38,39 MCs, macrophages, platelets, and fibroblasts can all produce large amounts of TGF-β1. In the process of skin fibrosis, the expression of TGF-β1 can promote fibroblast proliferation and induce the production of extracellular matrix (ECM). Although MCs are not the sole source of TGF-β1, this pleiotropic cytokine is perhaps the most well-characterized profibrotic mediator produced by MCs. 11 TGF-β1 secreted by MCs is an inactive form, a latent complex of TGF-β1, which is composed of mature TGF-β1, LAP TGF-β1, and latent TGF-β binding protein (LTBP). The latent TGF-β1 complex needs to release LAP TGF-β1 to expose the mature TGF-β1, which converts into a biologically active form.17,40 We found that both active TGF-β1 and LAP TGF-β1 were overexpressed in SLE skin. These data suggest that significant amounts of TGF-β1 has released LAP TGF-β1 and become the active form. Many experiments have shown that chymase and TGF-β1 can be released by MCs, and chymase immediately activates latent TGF-β1 complexes by disrupting the noncovalent interaction between the LAP TGF-β1 and mature TGF-β1. 41 We therefore speculate that in SLE skin, chymase has the same effect of inducing activation of TGF-β1. Activated TGF-β1 can not only trigger the pathological process of skin fibrosis but can also inhibit the proliferation and function of lymphatic endothelial cells to directly cause fibrosis of lymphatic vessels. 42
We found that the lymphatic vessels were dilated and the vessel walls had become thickened in SLE skin. In addition, much of the chymase and TGF-β1 was expressed near these malformed lymphatic vessels. In some regions, TGF-β1 expression was increased in areas where chymase was aggregated. Most significantly, we observed that TGF-β1 was not only diffused in the interstitial space but also expressed on lymphatic endothelial cells. Our in vivo result is consistent with the recent report by Wang peiliang, which showed that human lymphatic endothelial cells can express TGF-β1 in vitro. 43
There are several possible explanations for the above pathological phenomenon. During the early stages of SLE, lymphatic vessels may become dilated in response to the impaired lymph back flow. As the disease progresses, overloaded lymph results in the dysfunction of lymphatic vessels causing lymphatic fluid stasis in these dilated lymphatic vessels as well as in the interstitial space. Therefore, infiltrating microbes, specific antigens, and plasma proteins cannot be timely eliminated, which leads to metabolic disorders in the interstitium. These immunogenic substances will induce a chronic inflammatory reaction, for example, monocyte- and macrophage-produced inflammatory cytokines, such as tumor necrosis factor-α (TNF-α), interleukin (IL)-6, and IL-8.6,44,45 MCs also can be induced to proliferate and trigger IgE-mediated hypersensitivity reactions. The increased numbers of MCs secrete chymase and inactive TGF-β1, and chymase can cleave LAP TGF-β1 allowing TGF-β1 to become activated. Activated TGF- β1 combines with the type II and type III receptors in fibroblasts to form a complex, which phosphorylates Smad2/3 to activate downstream target genetic transcription and promote fibroblast proliferation and collagen deposition. TGF-β1 can also prevent matrix metalloproteinase-1 from cleaving collagens and activating the expression of tissue inhibitor of metalloproteinase by TGF-β1/Smad signaling, which breaks the balance between the production and degradation of ECM. 46 In addition, TGF- β1 can directly induce the expression of cytokines, such as connective tissue growth factor, to promote collagen synthesis.47,48 Finally, the increased collagen and the excessive deposition of ECM result in SLE skin fibrosis. 49
Collectively, we demonstrated a specific pathological phenomenon—significantly increased expressions of chymase and TGF-β1 in SLE skin. This phenomenon may be closely connected with the pathological events that lead to fibrosis. The results suggest that reducing the expression of activated TGF-β1 by decreasing chymase activity or expression may be helpful to inhibit tissue fibrosis. Therefore, our study may provide a novel means of treating chronic skin fibrosis in patients with secondary lower limb LE.
Footnotes
Acknowledgments
This study was supported by the Chinese National Science Foundation (grant number 81272146), National Key Specialty of Clinical Plastic Surgery Project, Shanghai Key Specialty of Clinical Reconstructive Surgery Project; Shanghai Science and Technology Committee (grant number12401900504).
Author Disclosure Statement
No competing financial interests exist.