
Abstract
Abstract
Background:
Although lymphedematous skin exhibits delayed wound healing, little is known about lymph drainage during wound healing. We investigated the wound healing process in the presence of lymphatic dysfunction.
Methods and Results:
The right inguinal lymph nodes (iLNs) and the surrounding tissue were excised in each mouse (the operation side), and a sham operation was performed in the left hindlimb (the control side). The next day, full-thickness wounds were made on both hindlimbs. The right hindlimb exhibited acute edema until day 3; however, it started to improve after day 4, and the wound area and epithelialization ratio were similar on both sides. Indocyanine green (ICG) was injected into both hindlimbs to observe lymph flow. On the operation side, ICG leaked out of the surgical site or remained at the injection site until day 2. Some lymph flow toward the existing lymph vessels was seen on day 3, and on day 10, lymph flow toward the axial LNs was detected on the operation side in all mice. On the operation side, the number of dermal lymph vessels was significantly increased on days 3 and 15. The dermal lymph vessel area of the peripheral wound was significantly smaller on the operation side.
Conclusions:
In a hindlimb lymphedema mouse model, lymph transiently accumulated in subcutaneous tissue, and then was gradually absorbed by the existing lymph vessels. The increase in the number of lymph vessels contributes to lymph drainage during wound healing. Acute lymphedema because of transient lymphatic dysfunction has little effect on wound healing.
Introduction
T
Harrell et al. 3 subjected a mouse to lymph node (LN) mapping. In their study, they reported that injection of Evans Blue dye into the planta of the hindlimb labeled the popliteal LN, which drain centrally to the iliac and renal LNs along the midline and also drain to the inguinal LN (iLN), followed by the midline. After this injection, drainage from the iLN to the axillary LN (aLN) was only occasionally observed. In our preliminary experiment, the subcutaneous injection of patent blue violet into the outer hindlimb labeled the iLN and the abdominal lymph vessels connecting the iLN to the aLN (data not shown).
Lymph vessels also play important roles in the pathogenesis of several diseases, such as cancer, various inflammatory conditions, and lymphedema. Lymph vessel impairment caused by surgery or radiotherapy can lead to secondary lymphedema. Lymphedema is a progressive disease characterized by gross swelling of the affected limb, accompanied by fibrosis, 2 decreased coordination and mobility, and susceptibility to infections. 4 Lymphedematous skin is so fragile that patients with lymphedema are recommended to perform various skin care procedures, such as keeping the skin clean, applying moisturizing cream, and examining their skin, in the clinical setting. 5 In addition, the skin of patients with lymphedema often exhibits repeated cellulitis and delayed wound healing due to its weakness; therefore, it is important to clarify the relationship between lymph drainage and wound healing in patients with lymphedema.
Generally, lymph vessels proliferate during inflammation. 6 After injuries, proinflammatory cytokines induce vascular endothelial growth factor (VEGF)-C messenger RNA transcription, presumably through NF-kB-mediated promoter activation. 7 As a result, lymphatic regeneration starts at the peripheral edges of wounds. 8 Previous studies have reported that increased interleukin (IL)-6 or IL-10 expression prevented macrophage migration, resulting in delayed wound healing in a mouse tail lymphedema model.9,10 Although these studies revealed that lymphedema leads to delayed wound healing associated with impaired immune responses, there have only been a few reports focusing on the relationship between lymph drainage function and wound healing.
In our previous study, 11 we made full-thickness wounds on wild-type mice and investigated the role of lymph drainage in wound healing. As a result, it was found that the lymph vessels around the wound area expanded on days 1–3 after wounding and these changes might have been due to the absorption of exudate leaking from the remaining blood vessels, and it was suggested that the peripheral lymph vessels of wounds are more important for wound healing than those that form in granulation tissue. However, little is known about the drainage of lymph during wound healing in lymphedematous skin. Therefore, the aim of this study was to induce lymphedema in mice and evaluate the wound healing process and lymphatic dysfunction after surgery.
Materials and Methods
The experimental protocol and animal care procedures were in accordance with the Guidelines for the Care and Use of Laboratory Animals of Kanazawa University, Japan (AP-122521).
Animals
In total, 75 BALB/cCrSlc female mice (Sankyo Lab Service Corporation, Inc., Toyama, Japan) weighing 16.5–21.8 g were used. They were caged individually in an air conditioned room at a temperature of 25.0°C ± 2.0°C, and the lights were kept on from 08:15 to 20:15 hours. Water and laboratory chow were given freely.
Mouse hindlimb lymphedema model
We created a new mouse model of surgically induced secondary lymphedema using a modified version of a previously published method.12–16 The mice were anesthetized with a mixture of medetomidine (Nippon Zenyaku Kogyo Co., Ltd., Fukushima, Japan), midazolam (Astellas Pharma, Inc., Tokyo, Japan), and butorphanol (Meiji Seika Pharma Co., Ltd., Tokyo, Japan) (0.01 mg/g weight) using an intraperitoneal (IP) injection, and then the bodies of the mice were shaved completely, and then hair removal cream was applied to the whole body.
In our preliminary experiments, the iLNs and the abdominal lymph vessels connecting the iLNs to the aLNs were identified after the subcutaneous injection of 4 μL of 2% patent blue violet (Wako Pure chemical Industries, Ltd., Osaka, Japan) solution into the outer hindlimb. The iLNs and the abdominal lymph vessels connecting the iLNs to the aLNs were labeled on both sides in all mice. The right iLNs were subsequently removed together with the associated peripheral lymph vessels and fat tissue using an electric knife, and then the surgical wounds were secured using 6-0 nylon simple interrupted sutures, and then they were bonded together and dressed with gauze (operation side). The left hindlimb was subjected to a sham operation, in which only the skin at 5 mm from the left iLN was excised so as not to injure the iLN (control side). After both operations, atipamezole (Nippon Zenyaku Kogyo Co., Ltd.) was intraperitoneally injected as an antagonistic drug.
Wounding
The day after the operation, the mice were anesthetized with 2% isoflurane (Wako Pure chemical Industries, Ltd.). Then, a circular (4 mm in diameter), full-thickness skin wound that included the panniculus muscle was made in each hindlimb with a biopsy punch (Kai Industries, Gifu, Japan), and the removed skin was weighed to evaluate the degree of edema.
The wounds made on the operation and control sides were covered with hydrocolloid dressing (Tegaderm; 3 M Healthcare, Tokyo, Japan) to maintain a moist environment. All mice were wrapped with sticky bandages. Elizabethan collars were used to prevent the mice from biting their wounds.
Observation of indocyanine green lymphography
Indocyanine green (ICG) is an intravascular fluorescent dye that has been used in ophthalmology to visualize the retinal and choroid vasculature for more than 40 years. 17 To investigate the lymph vessels that transport hindlimb interstitial fluid, we injected 2 μL ICG (2.5 mg/mL) into the outer hindlimb and massaged the injection site using absorbent cotton wool to promote ICG absorbance. After the injection, fluorescent images of lymphatic drainage channels were obtained using an infrared camera system (pde-neo; Hamamatsu Photonics, Shizuoka, Japan). Examinations of ICG flow were conducted on days 1, 2, 3, 6, and 10 after wounding. After detecting lymph flow, grayscale images of ICG flow were taken.
Macroscopic observations
The day on which the wounds were made was designated as day 0, and the wound healing process was examined on days 0, 3, 6, 9, 12, and 15 after wounding. We assessed each wound for edema, infection, and necrotic tissue. The wound edges were traced on polypropylene sheets, and photographs were taken at each time point. The traces on the sheets were captured with a scanner and transferred to a personal computer using Adobe Photoshop Elements 7.0 (Adobe System, Inc., Tokyo, Japan), and the area of each wound was calculated using the image analysis software, Scion Image Beta 4.02 (Scion Corporation, Frederick, MD). On day 3, the wounds on the operation side revealed marked edema, and the amounts of exudate being released from them were measured by calculating the hydrocolloid weight on day 3 minus the unused hydrocolloid weight to evaluate the degree of edema. The mice that were used for the examinations of wound healing were not used to study ICG flow.
Tissue processing
The mice were euthanized using the IP injection of a large dose of pentobarbital sodium (0.5 mg/g weight) on day 3 or 15 after wounding. The operative field or the sham operative field and the surrounding intact skin were harvested, stapled onto transparent plastic sheets to prevent the specimens contracting excessively, and fixed in 4% paraformaldehyde in 0.1 mol/L phosphate buffer (pH 7.4) for 18 hours. The specimens were then dehydrated in an alcohol series, cleaned in xylene, and embedded in paraffin to prepare 5-μm serial sections. These sections were stained with hematoxylin–eosin or immunohistologically stained with antilymphatic vessel endothelial hyaluronan receptor 1 (LYVE-1) antibody (RELIATech GmbH, Germany) to detect lymphatic endothelial cells. The antigen unmasking procedure was antigen dependent, as detailed below.
Immunohistochemical staining
After deparaffinization and rehydration, antigen unmasking was accomplished by heating the sections in a water bath, and then these sections were incubated in sodium citrate buffer (10 mM sodium citrate, 0.01% Tween 20; pH 6.0) for 20 minutes at approximately 100°C, washed with phosphate-buffered saline, and incubated with anti-mouse LYVE-1 rabbit polyclonal antibody (RELIATech GmbH, Wolfenbuettel, Germany) at a concentration of 1:100 at 4°C overnight. To detect the primary antibodies, the sections were incubated with polyclonal swine anti-rabbit immunoglobulins/horseradish peroxidase (DakoCytomation, Glostrup, Denmark) at a concentration of 1:100 for 30 minutes at room temperature, and then were incubated with the Dako liquid DAB+ substrate chromogen system (Dako North America, CA) (brown chromogen) for 5 minutes or until staining was detected at room temperature. Light hematoxylin counterstaining was applied for 1 minute to visualize the cell nuclei. Negative control sections were obtained by omitting each primary antibody.
Microscopic examination
We calculated the epithelialization ratio (%) as follows: the length of the new epithelium/the distance between the wound edges. To evaluate the lymph vessels, we counted the number of lymph vessels and calculated the area of the lymph vessel in the dermis during examinations performed with a light microscope at a magnification of ×200. The number of lymphatic endothelial cells in the dermis (number/mm2) was calculated as follows: the number of endothelial cells detected by LYVE-1 in the dermis/the area subjected to microscope observation, and the lymph vessel area of the dermis was defined as the sum total of the area of the lymph vessel in the dermis in the microscopically examined area/the number of lymph vessels. These calculations were conducted at three areas in one specimen, and the number of lymph vessels and the lymph vessel area of the dermis are described as mean values. The intact skin of wild-type mice was also evaluated using the same parameters.
Statistical analysis
Data are expressed as mean ± SD values and were analyzed using JMP ® 8.0.1 (SAS) (analysis of variance, multiple comparisons Tukey–Kramer). The differences were considered significant at p < 0.05.
Results
Wound healing process according to macroscopic observations
On day 0 (1 day after the removal of the iLNs), the mice were wounded using a punch biopsy, and the weight of skin removed by the punch biopsy was measured. The mean weight of the removed skin on the operation and control sides on day 0 was 4.6 ± 1.4 and 2.4 ± 0.8 mg, respectively (p = 0.003). From days 0 to 3, all wound surfaces were wet because of the release of exudate, and the areas of all wounds expanded (Fig. 1a).

Macroscopic examination of wound healing.
Necrotic tissue was observed on some wound surfaces; however, epithelia had started to cover the wound edges on day 3. Although the wounds on the operation side looked larger than those on the control side during the inflammatory phase, the difference was not significant (Figs. 1a, b). The mean amounts of exudate produced on the operation and control sides between days 0 and 3 were 62.0 ± 24.6 and 50.4 ± 27.9 mg, respectively (p = 0.25). After the inflammatory phase, the areas of all wounds gradually decreased, and epithelialization had been completed in all wounds (at the macroscopic level) by day 12. By day 15, all of the wounds had healed and exhibited scars. The wound area ratio did not differ significantly between the control and operation sides until day 15.
ICG lymphography
By subcutaneously injecting ICG into the outer hindlimbs of mice, we were able to preoperatively detect the iLNs and the abdominal lymph vessels connecting the iLNs to the aLNs in all mice (Fig. 2a). Table 1 shows the lymph flow detected by ICG lymphography. The ICG injected on the operation side leaked out of the surgical site field or remained at the ICG injection site until day 2 (Fig. 2b). On the operation side, lymph flow toward the aLN was detected in three of nine mice on day 3; however, lymph transiently accumulated in the subcutaneous tissue at the surgical site and was absorbed by the existing lymph vessels (Fig. 2c). On day 6, lymph flow toward the aLN was detected on the operation side in six of nine mice, and on day 10, lymph flow toward the aLN was detected on the operation side in all mice (Figs. 2d, e). On the control side, lymph flow toward the aLN was detected in all mice throughout the observation period.
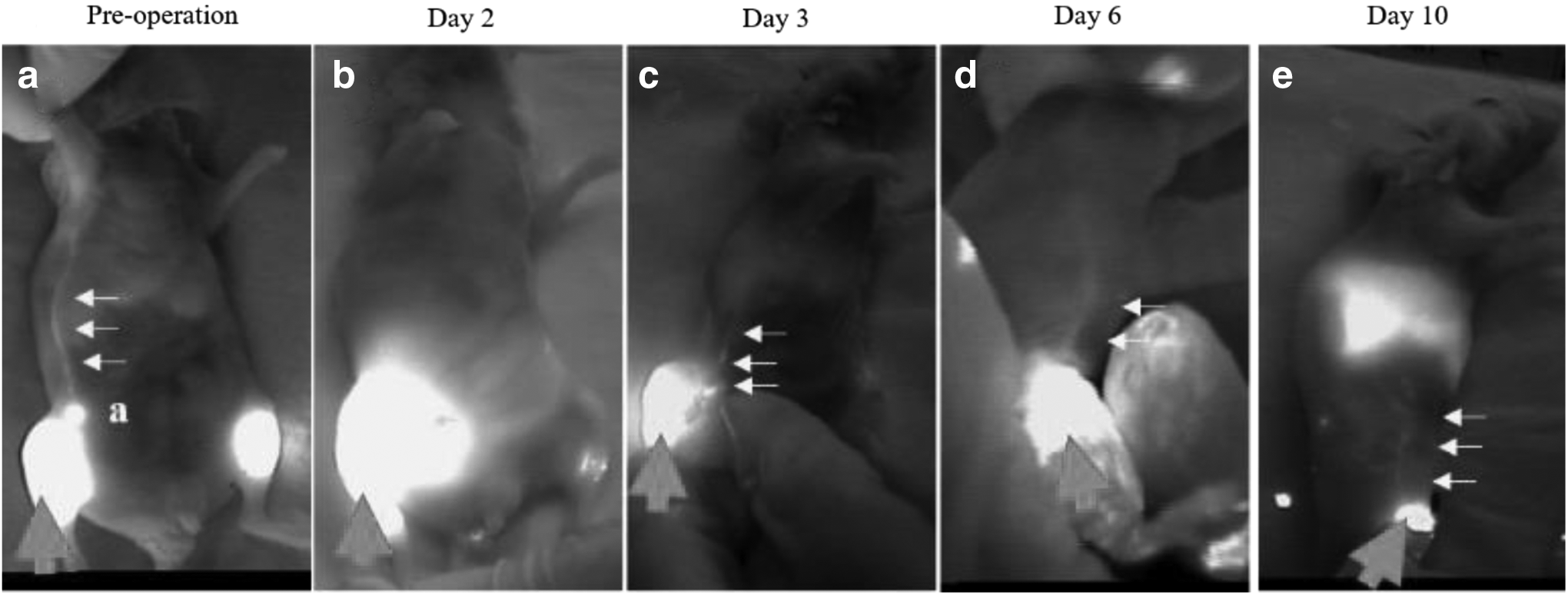
The lymph flow detected on the operation side using ICG lymphography. The small arrows indicate lymph vessels, and the large arrow indicates the ICG injection site (hindlimb).
The figures on the right indicate the total number of mice. The figures on the left indicate the number of mice in which lymph flow toward existing lymph vessels was detected. On day 10, lymph flow was detected in all mice.
Microscopic examination of lymph vessels
Figure 1c shows the epithelialization ratio data. Some epithelium had started to cover the wound edges on day 3, and the epithelialization was complete in all wounds by day 12. The epithelialization ratios of the control and operation sides did not differ significantly until day 15.
We used the anti-LYVE-1 antibody (LYVE-1 is a marker of lymphatic endothelial cells) to detect lymph vessels in the skin. Since we only observed a few lymph vessels in the subcutaneous tissue, we only evaluated the lymph vessels in the dermis. The operation or sham operation sites were located below and near to the wound areas, so we considered that the number of lymph vessels and the lymph vessel area at the operation or sham operation site would provide information about the drainage of exudate from the wound.
The number of lymph vessels in the dermis was markedly higher on the operation side than on the control side on day 3 (p = 0.0065) (Fig. 3a, c, d), and this difference persisted until day 15 (p = 0.0024). These numbers were twice as high as those seen on the control side and in the intact skin. However, the lymph vessel area of the dermis was smaller than those on the control side and the intact skin on day 3 and gradually increased until day 15 (Fig. 3b, e, f). On day 15, the lymph vessel area of the dermis was similar in all groups (Fig. 3b).
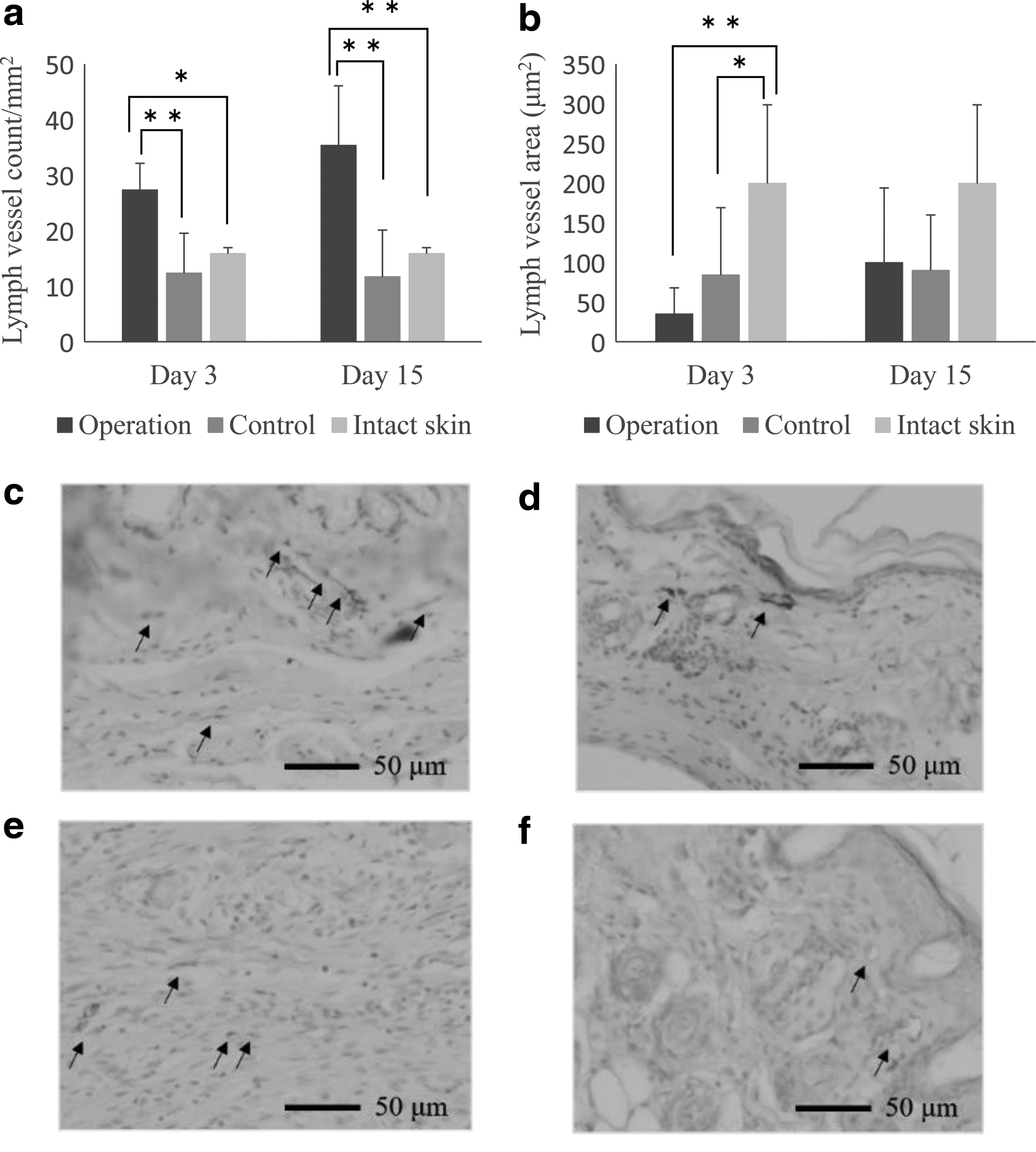
Conclusions
Hindlimb lymphedema mouse model
Many previous studies of lymphedema have used mouse tail lymphedema models.9,18–20 In this study, we wanted to evaluate the wound healing process in the presence of lymphedema and lymph flow during wound healing, so we chose a hindlimb lymphedema mouse model, in which the iLNs together with the associated peripheral lymph vessels and fat tissue were excised. Marked hindlimb edema was seen on the operation side, and the amount of exudate produced was significantly higher on the operation side than on the control side until day 3.
In addition, in ICG lymphography, no lymph flow was detected in the abdominal or axial areas of the operation side until day 2. These findings revealed that the operation performed in this study resulted in caused lymph flow obstruction in the groin region, resulting in lymphedema. However, lymph flow toward the aLN was detected in some mice on day 3, and it was found that most of the ICG flowing to the aLN transiently accumulated at the surgical site and then was absorbed by the existing lymph vessels. ICG flow on the operation side was detected in all mice by day 10. Some previous studies of chronic lymphedema demonstrated that surgery alone did not have a sustained effect on the condition,21,22 and the excess fat and muscle volume contributed to the total excess volume in excess soft tissue developed the first few years after breast cancer surgery. 23 The results in the present study did not reveal the sustained edema and the excess fat and muscle in the affected limb. Therefore, our hindlimb lymphedema model, in which the iLNs together with the associated peripheral lymph vessels and fat tissue were excised, produced acute, but not chronic, edema.
Lymph drainage during the wound healing process
In this study, we investigated the wound healing process and lymphatic dysfunction using a hindlimb lymphedema mouse model. In our previous study, 11 we produced full-thickness wounds in the dorsal skin of mice and investigated the relationships among exudate production, the wound area, angiogenesis, lymphangiogenesis, and reepithelialization during wound healing. In the latter study, we found that there were few new lymphatic capillaries in the granulation tissue that had formed approximately 2 days after wounding. Lymphatic regeneration began at the peripheral edges of the wounds from days 1 to 3, and the number of lymph vessels in the granulation tissue peaked on day 11, whereas blood vessels were first seen in the granulation tissue around day 3 or 4, and the number of blood vessels peaked on day 7. Therefore, the observed reduction in the amount of exudate produced by the wound appears to be related to blood vessel formation rather than lymph vessel formation.
In this study, the number of lymph vessels on the control side was similar to that seen in the intact skin; however, the lymph vessel area was significantly greater on the control side than on the operation side on day 3. These findings support those of our previous study and suggest that the expanded lymph vessels seen in the peripheral areas of the wound or existing lymph vessels play more important roles in exudate drainage than the new lymph vessels that arise in granulation tissue.
In addition, the number of lymph vessels on the operation side was significantly increased on days 3 and 15; however, the lymph vessel area on the operation side was significantly lower than that on the control side on day 3. Therefore, the number of new lymph vessels in the dermis increased in the peripheral areas of the wounds, which might have contributed to wound drainage, and this might have led to a reduction in edema and equivalent wound healing to that seen in the control. However, the drainage routes from the dermis to the subcutaneous peripheral wound were unclear because few lymph vessels were seen in the subcutaneous tissue in the present study.
It has been shown that VEGF-C can promote lymphangiogenesis by activating VEGF receptor (VEGFR)-2 and VEGFR-3 on lymphatic endothelial cells, and excess VEGF-C has been found to have ameliorative effects on edema produced by lymphatic obstruction in experimental models.1,24,25 In addition to the experimental distal lymphatic ablation in a mouse tail model, there is immunohistochemical evidence of a profound increase in cutaneous lymphatic vessel number and size that normalizes after VEGF-C administration. 26 However, it has recently been demonstrated that mouse tail edema can resolve even if capillary lymphangiogenesis is completely absent. In brief, this shows that the resolution of edema is not dependent on new lymphatic growth across the surgical obstruction, distal lymph fluid crosses the regenerating wound area interstitially, and the balance of interstitial forces might be more important than lymphangiogenesis for maintaining fluid drainage and tissue volume during acute lymphedema. 19
Edema resolution was also observed in the present study. Although no ICG flow was detected until day 2 after wounding, ICG transiently accumulated in the subcutaneous tissue at the surgical site and then was gradually absorbed by the existing lymph vessels. As a result, the lymphedema was resolved. Therefore, the present study supports the findings of the previous study. 19
We suggest that wound healing should not be delayed by hindlimb lymphedema because lymph drainage restarts. However, unfortunately, we could not identify the detailed dermal and subcutaneous lymph running routes from the peripheral wound areas to intact lymph vessels because lymphatic capillaries in the dermis were so small that we cannot detect them using ICG and there were few lymph vessels in the subcutaneous tissue, so we wonder how wound exudate drains to existing lymph vessels or LNs. In addition, the hindlimb edema resolved after day 4 in the present study; therefore, it will be necessary to develop a sustained lymphedema model to allow us to evaluate lymph drainage during wound healing. These are the limitations of this study, and we will investigate these issues in future.
Footnotes
Acknowledgments
This study was supported by JSPS KAKENHI Grant Number 25293430 to T. Nakatani and research funding from Kanazawa University and Meiji Yasuda Life Foundation of Health and Welfare.
Author Disclosure Statement
The authors declare that they have no competing financial interests. All the authors were involved in the study design, data collection, data analysis, and the editorial process and approved the final version of the manuscript.