
Abstract
Abstract
Background:
Mesenteric lymphatic vessels (MLVs) from various animal species have been intensively studied. We aimed to establish the viability and basic contractile characteristics of human MLVs maintained in vitro and to determine the reactivity of MLVs with norepinephrine (NE) and substance P (SP) and to compare with the thoracic duct (TD).
Methods and Results:
Isolated human lymphatic vessels were mounted on a wire myograph under isometric conditions and tension was recorded. The diameter–tension characteristics for MLVs were generated by stretching the vessels and stimulating with a 125 mM K+ solution containing 10 μM NE. The diameter–tension data generated for MLVs from two separate surgical patient groups were found to be similar: maximum active tension for MLVs occurred when the passive stretch corresponded to a transmural pressure of 22 mmHg. Subsequent experiments on human MLVs were performed by normalization with 22 mmHg as the equivalent target pressure. The majority of MLVs were responders (spontaneous activity and/or reactivity with 10 μM NE or 125 mM K+ solution). Nonresponders (16% of vessel segments) had significantly smaller inner diameters. MLVs responded consistently to NE (1 nM–10 μM) but the responsiveness of MLVs and TD to SP (0.1 nM–10 μM) was poor: TD reacted only with 10 μM SP, whereas MLVs were sensitive to nanomolar concentrations and the contractile response declined with higher concentrations.
Conclusions:
Under in vitro isometric conditions, human MLVs generate maximum tension when stretched to a passive level corresponding to 22 mmHg, and the majority of MLVs are responsive when normalized to this pressure. MLVs respond to NE and SP though NE produces a more consistent response in the concentration range tested.
Introduction
T
Lymphatic pumping is modulated by neuronal and humoral input as well as pressure and flow. Substance P (SP) is a neuropeptide that was discovered in 1931 8 and is released by nerves and immune cells. It can increase blood capillary permeability and act both as a vasoconstrictor and as a vasodilator of blood vessels. 9 Furthermore, SP is involved in different inflammatory conditions of the respiratory tract, gastrointestinal tract, and joints. 10 The effect of SP on lymphatic pumping has been extensively studied in animal models, in which chronotropic and inotropic responses have been reported.11,12 Vasoactive intestinal peptide (VIP) is another neuropeptide released by nerves and immune cells. In contrast to SP, VIP inhibits lymphatic pumping. 13 Neuropeptides thus modulate lymphatic pumping and lymph flow can be affected in a beneficial or detrimental manner. Given the close connection between the lymphatic vasculature and inflammation and in light of several studies of animal lymphatic vessels employing SP, the characterization of this mediator in human lymphatic vessels is paramount to elucidate how inflammatory mediators influence human lymphatic function.
In this study, we test the following hypotheses: (1) that human MLVs show a similar diameter–tension relationship to the human TD and (2) that norepinephrine (NE) and SP are vasoconstrictors of human MLVs.
Materials and Methods
Tissue preparation
A piece of mesentery was harvested from jejunum and in proximity to the superior rectal artery from patients undergoing gastric bypass surgery (GBPS) (n = 33, age span 25–58 years) and ileal pouch–anal anastomosis (IPAA) (n = 7, age span 19–35 years) surgery because of inflammatory bowel disease. GBPS was performed at Department of Surgery, Viborg Hospital, whereas IPAA surgery was conducted at Department of Surgery, Aarhus University Hospital. TDs were harvested from patients undergoing esophageal and cardiac cancer surgery (n = 4, age span 56–69 years) at the Department of Cardiothoracic Surgery, Aarhus University Hospital. Lymphatic vessels were identified under a stereomicroscope and dissected free from surrounding tissue. TDs were furthermore harvested from adult Wistar rats (n = 9) sacrificed by CO2 inhalation. The study was approved by the Central Denmark Region Committees on Health Research Ethics (File nr M-20070194) and conducted in accordance with the ethical principles specified in the Helsinki declaration.
Isometric force measurements
Ring segments (2 mm long) of MLVs and TD were prepared and mounted on 40 μm stainless steel wires in multichannel myographs for isometric force recordings. The preparation, mounting, and experiments were performed in physiological saline solution (PSS; for composition, see hereunder). The vessels were maintained at 37°C in PSS equilibrated with a mixture of 20% O2 and 5% CO2 throughout the experiments (pH 7.4). Isometric force development was recorded at 40 Hz with a Powerlab4/25 (ADInstruments) using LabChart software. Data files were saved for offline analysis. Force data were converted to tension, Newton/meter (Nm−1), by dividing the force (mN) by two times the segment length (mm). Transmural pressures were calculated according to Laplace's law: transmural pressure = tension/radius. Radius was calculated by the following equation: [(total wire circumference +2 × wire diameter) + (2 × the distance between the inner edges of the wires)]/2π. Contraction frequency and amplitude of spontaneous activity were extracted from all experiments except when stimulation with NE or K-PSS (125 mM K+, for composition see Solutions Section) was used, in which average tension during the stimulation period was extracted.
Diameter–tension protocol
MLVs from both GBPS (n = 7) and IPAA (n = 7) patients were used to establish the diameter–tension relationship. The vessels were mounted on the myograph and allowed to equilibrate for 30–60 minutes under zero tension. The vessels were first stretched and after the vessels reached steady-state tension, the vessels were activated with K-PSS and 10 μM NE. After a peak in force was observed, the activation solution was washed out by exchanging the chamber solution with PSS several times until baseline tension was reached. The stretch–activation procedure was repeated stepwise with 50–100 μm increments in diameter until the active force produced by the activation solution declined at two consecutive diameters. For each diameter step, the passive tension before activation and peak activation tension (total tension) were determined with the calculated difference representing active tension. Passive, active, and total tension were plotted against diameter and fitted with second (active tension) and third order (passive and total tension) polynomial regressions. The diameter producing peak active tension was termed L0 and all diameters were plotted in relation to this (L/L0). The passive wall tension at L0 was converted to a pressure using LaPlace's law and an average for all vessels was calculated that permitted subsequent vessel segments to be normalized using this value.
Normalization, spontaneous, and evoked contractility
After mounting, the ring segments were stepwise stretched to a diameter at which the wall tension was equivalent to 21 mmHg (TD) 7 or 22 mmHg (MLVs: determined in this study), allowing maximal force production. After normalization, the vessels equilibrated for 30–60 minutes.
The equilibration period was used to observe and analyze spontaneous activity. Both spontaneously and nonspontaneously active vessels were included in our experimental protocol. Only vessel segments displaying regular phasic contractions were included for analysis of spontaneous activity because of difficulties in analyzing irregular patterns of spontaneous activity. Nonspontaneously active vessels were classified as viable (responders) if they showed reactivity with NE. Vessels unreactive with NE were challenged with K-PSS and deemed nonresponders if they did not contract. Rat TDs were passively stretched to 0.40 mN. All SP experiments were performed in PSS supplemented with 0.5% bovine serum albumin.
MLVs (n = 17–19) from patients undergoing GBPS were challenged to cumulative concentrations of NE (1 nM–10 μM, half-log steps every fifth minute) or SP (0.1 nM–10 μM, half-log steps every fifth minute). Each vessel was tested with NE and SP, with washouts between each cumulative concentration–response experiment and the order of the drugs randomized.
TDs from humans (n = 4) and rats (n = 9) were exposed to a cumulative concentration–response curve of SP (1 nM–10 μM, half-log steps every fifth minute).
Solutions
All salts and drugs were purchased from Sigma-Aldrich. All drugs except SP (acetate buffer) were dissolved in distilled water and aliquots were stored at −20°C until required. PSS of the following composition was used (in mM): 119 NaCl, 4.7 KCl, 1.17 MgSO4, 25 NaHCO3, 1.18 KH2PO4, 0.026 EDTA, 5.5 glucose, and 1.6 CaCl2. K-PSS (125 mM) was made by substituting all NaCl for KCl.
Statistics
Data are presented as mean ± standard deviation. Grouped data were compared using Student's t-test. Correlation between contraction amplitude and frequency was analyzed using a linear regression. Significance level was set to p = 0.05 in all analyses. Each patient provided us with one TD or one to several MLVs. The vessels were divided into vessel segments, which allowed us to use the specimen for multiple experiments. The reported n always represents the number of patients.
Results
Diameter–tension properties
The diameter–tension relationship generated for MLVs from IPAA and GBPS patients was quite similar (Fig. 1). The passive tension at L0 for IPAA and GBPS patients was 0.47 ± 0.24 Nm−1 and 0.64 ± 0.46 Nm−1, respectively, whereas active tension was 0.78 ± 0.4 Nm−1 and 0.93 ± 0.45 Nm−1, respectively. The passive tension values correspond to equivalent transmural pressures of 22 ± 8.2 mmHg and 22 ± 11 mmHg (p = 0.88), according to Laplace's law, when the vessels are set to L0. The maximal pressure the vessels could generate (derived from the active tension curve) at this diameter was 37 ± 18 mmHg and 41 ± 19 mmHg (p = 0.64) for IPAA and GBPS, respectively. Both diameter–tension relationships revealed that >80% of the maximal force was available at 0.7–1.3 × L0. MLVs were normalized in subsequent experiments by a passive stretch protocol until the tension attained corresponded to 22 mmHg.
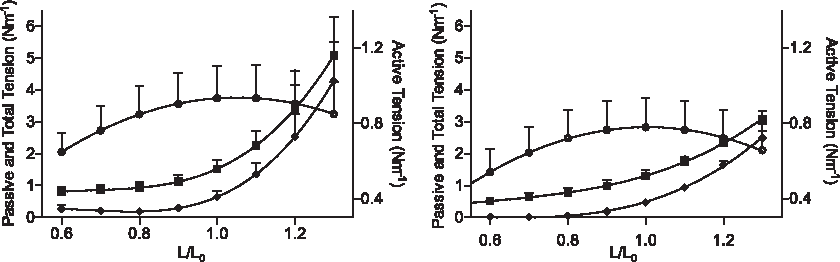
Diameter–tension properties of mesenteric lymphatic vessels (MLVs) harvested from patients undergoing bariatric surgery (left panel) and ileal pouch–anal anastomosis surgery because of inflammatory bowel disease (right panel). The passive (♦) total (■) tensions measured are depicted on the left y-axis, whereas the derived active tension (¡) is depicted on the right y-axis to show the shape of the curve more clearly. Tension is plotted against normalized internal diameter with L0 being the diameter corresponding to maximum active tension. In both plots n = 7.
Spontaneous activity
A total of 105 MLV segments from 29 patients undergoing GBPS were mounted on a wire myograph and normalized to 22 mmHg. In the initial 30–60-minute equilibration period, 37 vessel segments displayed spontaneous contractions. A further 24 vessel segments displayed spontaneous contractions after viability check with 10 μM NE and subsequent washouts or at a later time point during the protocol. A total of 61 vessel segments thus displayed spontaneous activity (58%) and out of these, 28 displayed regular spontaneous contractions, whereas 33 had an irregular contraction pattern. Of the nonspontaneous vessels, 27 vessels responded to NE or KPSS stimulation (26%), which left 17 nonviable vessel segments, which we termed nonresponders (16%). The majority of patients (26 out of 29) thus provided us with at least one responding vessel segment. Comparison of the inner diameters of the responders (149 ± 40 μm) and nonresponders (114 ± 36 μm) revealed that the nonresponders were significantly smaller in diameter (p = 0.0139; Fig. 2).
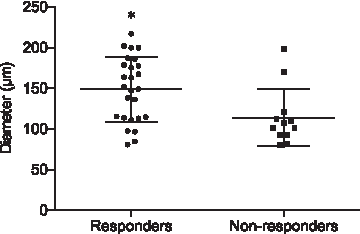
Distribution of average internal diameter of MLVs displaying either spontaneous activity or reactivity with norepinephrine (NE) or KPSS (responders) and nonviable vessels (nonresponders). Nonresponders were significantly smaller in diameter. Bars depict mean ± standard deviation (SD) (n = 26). Asterisk marks represent significant difference (p = 0.0139).
The spontaneous contractions had an average frequency of 3.4 ± 1.7 minutes−1 with contraction amplitude of 0.41 ± 0.24 Nm−1, which corresponds to 20 ± 10 mmHg (Fig. 3).
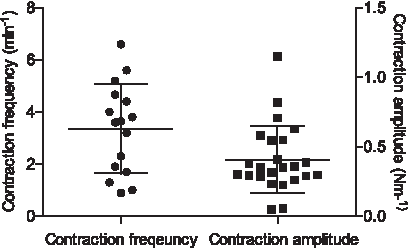
Distribution of spontaneous contraction frequency and contraction amplitude for MLVs mounted on an isometric wire myograph. In the instance of a patient vessel providing multiple segments with spontaneous activity, the average was calculated (n = 24). Bars indicate mean ± SD.
Reactivity with SP and NE
MLV segments from patients undergoing GBPS were exposed to cumulative concentrations of both NE and SP (Figs. 4 and 5A). NE induced contractions in a concentration-dependent manner in all vessel segments, with a single exception. The generalized NE response consisted of phasic contractions on an increased baseline tension with small oscillations occasionally superimposed on some of the phasic contractions. This response was similar to what we have reported previously for the human TD, except the phasic contractions did not develop into a plateau of small high-frequency oscillations with high NE concentrations. In MLVs exposed to SP, 6 of the 19 MLVs tested did not show any reactivity. Only two vessel segments responded by increasing tension in a concentration-dependent manner. In the remaining 13 vessels, SP induced contractions in the low concentration range, but the response diminished with increasing concentrations until a plateau was reached at approximately 10 nM. From 10 nM to 10 μM SP the responsiveness was poor. There was no difference in the maximum contractile response elicited from either SP or NE (p = 0.45).
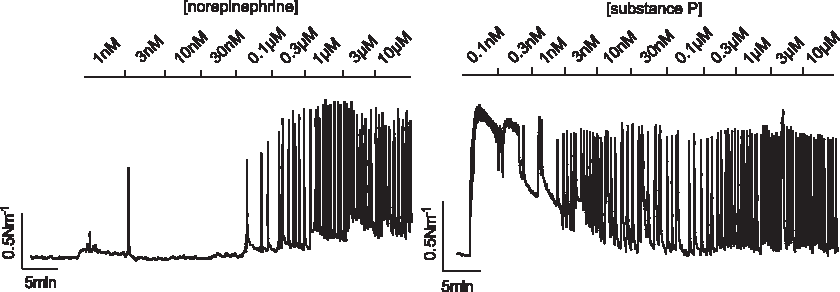
Representative traces from cumulative concentration–response experiments examining NE (left) and substance P (SP) (right) responsiveness in human MLVs. Whereas NE increased contractile level across the range of concentrations tested, SP induced a strong initial response that declined thereafter.
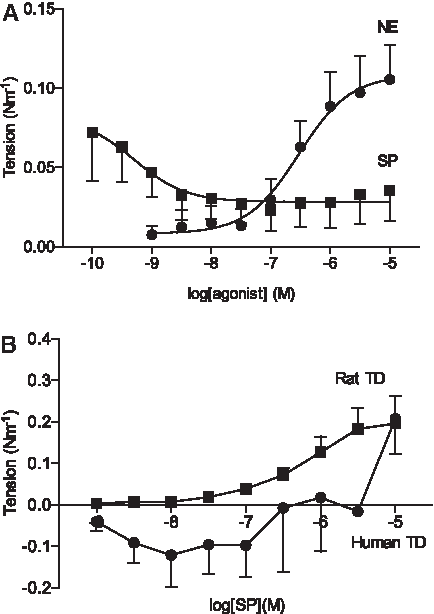
Average concentration–response profiles of human MLVs, human thoracic ducts (TDs), and rat TDs after exposure to NE or SP.
Human TD segments were also challenged with increasing concentrations of SP, but these vessels only showed a weak response at the highest concentration despite consistent reactivity with NE (Fig. 5B). To validate the SP viability, a parallel series of experiments using the same SP batch on rat TD ring segments was performed. In these experiments, SP induced contractions in the expected concentration range (Fig. 5B), as described previously in rats. 11
Discussion
Here we have studied the basic characteristics of human MLVs, including diameter–tension relationship and spontaneous activity, as well as reactivity with NE and SP. Mesenteric vessels and SP are widely employed in animal studies and it is, therefore, crucial to understand how accurately these results translate to the human situation.
We have previously described the diameter–tension relationship of human TDs 7 and have since used this knowledge to successfully investigate TDs in vitro under standardized manners.3,4,14,15 We used MLVs from two different patient groups to generate diameter–tension relationships and calculated the transmural pressure at the diameter producing maximal active force to be 22 mHg in both patient groups. It is noteworthy that the passive and active pressures obtained in this study from the diameter–tension relationships are highly congruent with our previous study of the human TD. In particular, the average passive transmural pressure only differed by 1 mmHg between the two vessel types and the standard deviations were similar (8–11 mmHg). 7 Although the normalization value for the TD resembled in vivo measurements of TD cannulations, there are, to the best of our knowledge, no equivalent pressure measurements of human MLVs in vivo to compare our values with. The active force available was also similar between the MLVs and TD: 37 ± 18, 41 ± 19, and 47 ± 15 mmHg and are of physiologically relevant magnitudes.
Spontaneous activity
It is well known that lymphatic vessels have an inherent, spontaneous contractile ability, but the mechanism underlying these contractions is not yet completely understood. From published literature, it is evident that, regardless of vessel type and species, not all lymphatic vessels studied in vitro exhibit spontaneous contractions: nonspontaneously active vessels have been reported to range from 10% 16 to 40%–50%.17–19 In this study, we observed that initially (within 30–60 minutes after normalization) 35% of the vessel segments exhibited spontaneous contractions, although later in the protocol, this number increased to 58%. In comparison with our similar study of the human TD, both the number of MLVs displaying spontaneous contractions initially was higher (35% vs. 22%) as was the contraction frequency (≈3.5 in MLVs vs. ≈1.4 minutes−1 in TD). The difference in contraction frequency between the two vessel types is consistent with a previous comparison of the TD and MLVs from rats. 20 The amplitude of the spontaneous contractions was slightly higher than values obtained from cows and rats: 20 mmHg compared with ≈14 mmHg 21 and ≈8 mmHg, 22 it is important to note that our pressure is a derived value whereas the other two values are measured directly. However, our results confirm that contraction amplitude is generally related to animal size and that humans have lymphatic vessels capable of generating high pressures. 2
Sixteen percent of the vessel segments were nonresponders, that is, without spontaneous activity or reactivity with NE or KPSS, and further analysis showed that these vessel segments were significantly smaller in diameter than the responders. We speculate that these vessel segments might represent precollectors. Precollectors are lymphatic vessels connecting the amuscular initial lymphatics with the muscular collecting vessels. The precollectors are smaller than the collecting lymphatics and it is unclear whether they have smooth muscle cells,23,24 which could explain the lack of force production from the vessel segments we classified as nonresponders. Another plausible explanation could be the higher risk of damaging smaller diameter vessels during mounting and we cannot exclude that this could have influenced our findings.
NE and SP reactivity
SP is one of the most commonly used vasoactive neuropeptides in lymphatic physiology research and proposed to alleviate inflammation-induced edema by increasing lymphatic removal. 25 SP has been shown to be a potent vasoconstrictor in both lymphatic vessels from rodents and elicits contractions in quiescent vessels: consistent concentration-dependent increases in tension or diameter, as well as contractile frequency, have been reported in the nanomolar to micromolar range under isometric and isobaric conditions.11,12,26 Even when SP has ultimately induced severe constriction and reduced contraction amplitude, the chronotropic effect has been shown to maintain increased flow.26,27 Based on immunohistochemistry, small and large human lymphatic vessels have been shown to possess nerves containing SP.28,29 We have previously observed that a discrete high concentration (10 μM) of SP constricted isolated human TD ring segments. 15 We expanded upon this earlier observation by testing the range of SP concentrations reported to induce vasoconstriction (1 nM–10 μM) in both human TD and MLVs ring segments. Although NE proved to be a consistent vasoconstrictor, the response to SP was atypical in that it differed from observations published from animal experiments and between the two vessel types. In the TD experiments, SP elicited a contractile response only at the highest concentration (10 μM) tested. We excluded technical limitations regarding SP's viability by testing the same batch in parallel on rat TD, and in these experiments, contractions were generated in the expected concentration-dependent manner and were comparable to previously published findings. 11 The peak tension response to SP was similar between the human and rat experiments, confirming poor reactivity in the human TD because there is considerably more force available for contraction in the ≈5–10 × larger human TD. The MLVs responded in a different manner to SP: an initial reactivity with low nanomolar concentrations was observed in 13 out of 19 vessels, which then declined with increasing concentrations. We speculate that the decrease in response with increasing concentrations could be caused by tachyphylaxis, which is a well-known phenomenon to SP. 30 It is, however, interesting to note that tachyphylaxis has not been reported in animal lymphatic vessels using the concentrations of SP employed here. We cannot exclude that the sensitivity for human MLVs to SP is in the subnanomolar range nor that SP may have vasodilatory effects in human lymphatic vessels as previously reported for human arteries and veins. 9 Another possibility is a limited release of TxA2 from the endothelium, 12 which would be consistent with our previous finding of U46619 being a vasoconstrictor in human TD. 7 Given the dramatic difference between human SP reactivity and that reported from animal studies, further investigation into the mode of action of SP in human vessels is warranted.
Summary
We demonstrate here that human MLVs, a vessel type widely employed in animal studies, can be successfully isolated and maintained in vitro for assessment of tone. Isolated human MLVs maintained under our conditions generate tension increases that correspond to physiologically relevant pressures. Discrepancy between the SP reactivity described here and that obtained from animal studies indicates care must be taken when extrapolating SP data generated from animals to human lymphatic physiology and pathophysiology.
Footnotes
Acknowledgments
The authors thank Anders Tøttrup, MD and Jonas Funder, MD as well as several staff members at the Department of Surgery, Aarhus University Hospital, for their invaluable contribution in acquiring tissue.
Author Disclosure Statement
No competing financial interests exist.