
Abstract
Abstract
The lymphatic system serves as the primary route for the metastatic spread of breast cancer cells (BCCs). A scarcity of information exists with regard to the advection of BCCs in lymph flow and a fundamental understanding of the response of BCCs to the forces in the lymphatics needs to be established. This review summarizes the flow environment metastatic BCCs are exposed to in the lymphatics. Special attention is paid to the behavior of cells/particles in microflows in an attempt to elucidate the behavior of BCCs under lymph flow conditions (Reynolds number <1).
Abstract
Background:
The lymphatic system is an extensive vascular network that serves as the primary route for the metastatic spread of breast cancer cells (BCCs). However, the dynamics by which BCCs (10–40 μm) travel in the lymphatics (100–300 μm) to distant sites, and eventually establish metastatic tumors, remain poorly understood. The lymphatic flow environment is extremely complex, and despite the number of studies carried out in recent years, an accurate description remains unclear.
Methods and Results:
Over the past two decades, significant advancements have been made in characterizing lymphatic flow, using numerical and experimental approaches, however, none of these studies addresses the dynamics of BCCs flowing in lymph. This review summarizes the flow environment metastatic BCCs are exposed to in the lymphatics. Special attention is paid to the flow behavior of cells/particles in microdevices in an attempt to elucidate the behavior of BCCs under lymph flow conditions (Reynolds number <1). The advection of BCCs in microflows is governed by the hydrodynamic forces present in the flow.
Conclusions:
A scarcity of information exists with regard to BCC advection in the lymphatics and this review highlights important areas for future research. A fundamental understanding of the response of BCCs to the forces in the lymphatics needs to be established. More comprehensive theoretical and experimental techniques are needed to quantify the effect of the lymphatic hydrodynamic forces on the behavior of BCCs, resulting in a more concrete basis for the numerical simulation of flow-induced shear stresses experienced by BCC membranes.
Introduction
T
Accurate measurement of the fluidic environment BCCs are exposed to in the lymphatics is important as the flow induced shear forces and the flow environment surrounding cancer cells can modulate their metastatic potential. 54 Therefore, establishing the fluidic environment present in the lymphatics is vital to accurately characterize metastatic cell transport in this vascular system.
Microfluidic systems are regularly used to sort, separate, and analyze biological cells, providing a better approximation of the cellular environment compared with conventional cell culture techniques. 55 Microfluidic devices can mimic the cellular and fluidic environment in vivo by precisely controlling system inputs such as intercellular interactions, gradients of soluble factors, and application of mechanical forces,56,57 making it possible to investigate the biological response of cells to various physiological stimuli on a micrometer scale.
Typically the focus is on high-throughput devices whereby the goal is to exploit microscale fluidics, using small quantities and high surface to volume ratios, to drastically reduce processing times over conventional laboratory-based assays. Consequently, there is a considerable wealth of research that examines transport properties of particles and biological cells in microfluidic devices.58–70 However, there is very limited data available on the behavior of large particles/cells (>10% the channel width) under very low flow rates (Reynolds number (Re) < 1) such as those seen in the lymphatics.
Lymph contains cells that are 5–8 μm in diameter and breast cancer cells (BCCs) vary from 10 μm to 40 μm.66–69 The dynamics by which BCCs travel in the lymphatics to distant sites, and eventually establish metastatic tumors, remain poorly understood. However, the mechanical microenvironment surrounding cancer cells is known to play an important role in modulating their response and function. 7 In addition to the mechanical environment, the immune environment is known to play a major role in regulating cancer metastasis, as studies have shown that cancer cells are guided into the lymphatics by chemokines produced by lymphatic endothelium.71,72
Originally, the lymphatics were viewed as a transportation system, which played a passive role in cancer metastasis; however, recent studies indicate that their role is far more complex.71,73 Interest in the use of microvascular networks has increased dramatically over the past decade due to the several advantages they provide with regard to their 3D nature and real-time monitoring of the response of cells to stimulants such as drugs or hydrodynamic forces. 57 The development of microfluidic models that can recapitulate the immune environment surrounding cancer cells is required to further our understanding of the responsiveness of certain cancer types to immunotherapy. 57 Accurate assessment of this environment will present a powerful tool for predicting patient treatment outcomes, by providing a means to characterize drug efficacy and responsiveness.
This review summarizes the recent advancements made in lymphatic system research, with special attention paid to modeling the lymphatic flow environment. A number of specific research modalities, including analytical modeling, computational modeling, in situ studies, and in vitro studies are discussed. The fundamental mechanisms that govern particle advection (two-phase flow) in confined flows are then introduced before the transport of biological cells is examined.
Finally, key areas for future research are highlighted, including the development of microfluidic models of BCC flow and the use of multiphase numerical modeling techniques to characterize the flow-induced surface forces experienced by BCC membranes. The dynamics of BCC transport through lymph is relatively unknown, defining the flow-induced shear stresses and the shear stress gradient BCC membranes experience within the lymphatics may provide insight into the BCCs biological response, which could be exploited in future cell culture models, potentially leading to improved knowledge on BCC advection in the lymphatic environment.
Lymphatic Vessels in Cancer Metastasis
Since the discovery of lymphatic-specific growth factors and markers, research in the lymph system has increased exponentially, with many investigating the role lymphatics play in metastasis.1,3,71–78 Traditionally, dissemination of cancer cells to organs distal to the primary tumor has been thought to be initiated only through tumor-associated blood vessels and not lymphatics. However, it is now well established that lymph vessels invade tumors in a fashion similar to blood vessels. The flow environment of the lymphatics provides a more attractive route for BCCs compared with the highly dynamic environment in the circulatory system. 79 Studies have shown that circulating tumor cells (CTCs) can actively induce lymphatic metastasis and tumor-associated lymphangiogenesis by secreting certain growth factors, which promote their spread to sentinel lymph nodes.1,2,80
Several reviews have summarized this biological aspect of metastasis4,5,81–91; however, the main focus of this review is the lymphatic fluidic environment and how metastatic BCCs respond to the flow conditions from a mechanical/fluidic perspective. Research has shown that the interaction between cancer cells and their environment can enhance the cancer cell progression.54,92 Hemodynamic shear stresses of up to 60 dynes/cm2 have been reported to increase cancer cell apoptosis in the circulatory system,93,94 whereas to the authors' knowledge, the effect of the shear stress levels found in the lymphatics on the biological response of BCCs has yet to be quantified. Thus, elucidating the effect the lymphatic flow field has on BCC behavior is of necessary importance.
The mechanisms of cancer cell intravasion into the lymphatics remain elusive. 73 The passive role the lymphatics were originally assumed to play in cancer metastasis has recently been questioned as studies have shown that cancer cells are directed into the lymphatics by chemokines produced by the lymphatic endothelium.71,73 Tumor cells utilize interstitial flow to create autologous chemokine gradients and thus chemotact toward draining lymphatic vessels. 72 The lymphatic endothelial cells (LECs) play an active role in cancer metastasis, and recently lymph node LECs were found to control cancer cell entry into lymph nodes. 78 Lymph nodes act as holding reservoirs, where BCCs take root and form metastatic tumors, (Fig. 1).
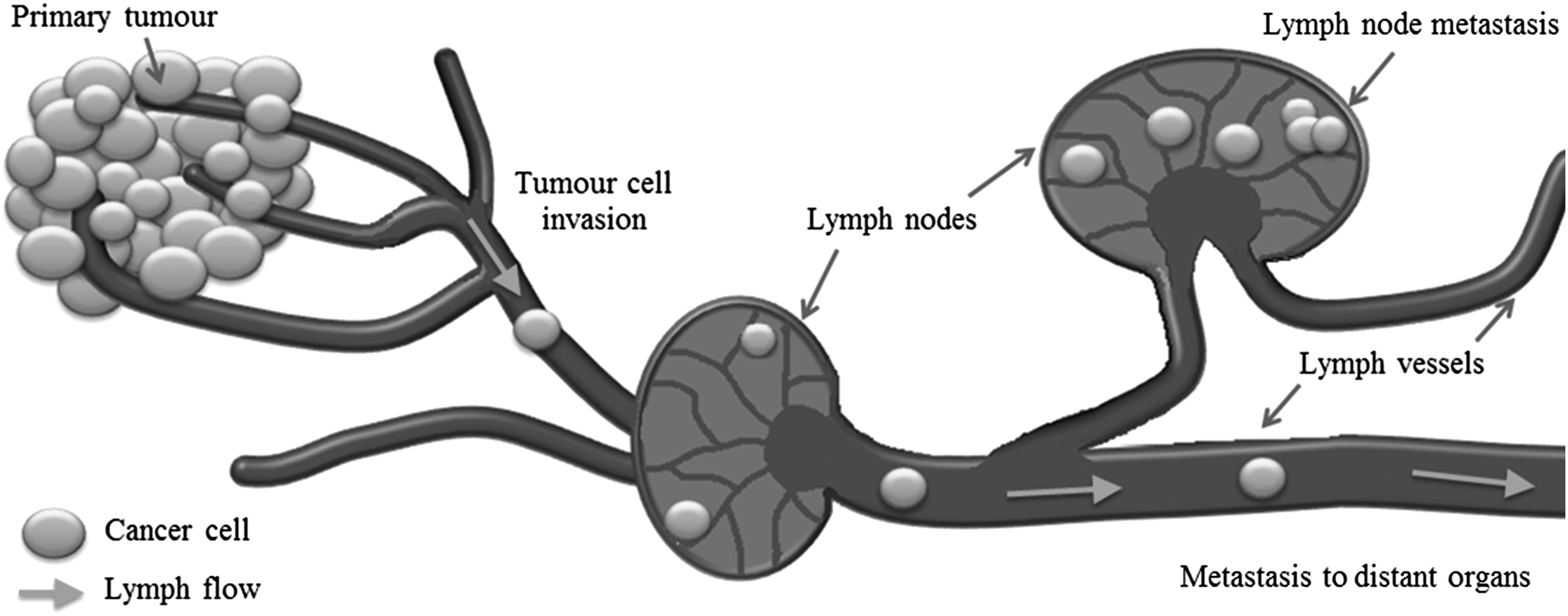
Lymph vessel and lymph node anatomy. Tumor-draining lymph vessels cause cancer cells to spread from the primary tumor site to distant locations in the body through the lymph nodes. Metastatic foci take up root in the lymph nodes forming secondary tumors.
Rogue BCCs have evolved ways of evading the immune surveillance mechanisms present in lymph nodes and thus are able to successfully colonize and proliferate. Approximately 8% of tumor cells that arrive in the nodes are successful in forming overt tumors. 95 The exact mechanisms by which BCCs move preferentially toward particular lymph nodes remain poorly understood. Research has shown that the entry of cancer cells from the lymphatics into the lymph nodes consists of at least two stages, (1) cancer cell entry into the sinus and (2) migration across the sinus floor into the lymph node cortex. 78 It is not yet known whether other biological characteristics of breast cancers affect the risk of nodal involvement.
There is huge potential for concentrating chemotherapeutics to particular lymph nodes, as opposed to the circulatory system, so as to avoid systemic dilution and dose-limiting side effects. 3 It has been shown that lymphatics alter their function in response to various mechanical stimuli 45 ; therefore, it is essential that the impact of diseases on the mechanical environment be elucidated. The exact relationship between the biomechanical factors and the biochemical responses of BCCs remains to be identified.74,96 The interplay of several factors is required for BCCs to metastasize in the lymphatics; however, the information available on this topic is scarce at best and a large number of uncertainties remain.
The biomechanical environment in the lymph system plays a key role in the progression of breast cancer; therefore, exposing the effect BCCs have on the flow field is of crucial importance. This review emphasizes the importance of future work focused on investigating BCC deformability and size effects in the lymphatics. One of the most important factors that determines the behavior of cells is the surface forces the cell membranes are exposed to. Recently, a study concluded that the mechanical forces generated by fluid flow regulate cellular behavior in terms of metastatic potential and proliferation. 97 Therefore, establishing the flow induced surface forces BCC membranes are subjected to in the lymphatics is critical. Combining this information with knowledge of the BCCs response to chemotactic signals 78 will improve our understanding of the behavior of BCCs in lymph flow.
The response of BCCs to the physical microenvironment in the lymphatics may be exploitable for the treatment of breast cancer. Recapitulation of the key aspects of the environment, such as quantifying the flow-induced surface forces BCC membranes are exposed to, may lead to improved cell culture experiments. Translation of these findings to biological characterization experiments offers the potential to create more accurate test beds for breast cancer metastasis studies.
Modeling the Lymphatic System
Analytical modeling
The lymphatic system represents a challenging system for modeling, which led to several groups utilizing analytical methods as a first step toward characterizing its behavior. The lymphatic system is vastly underresearched when compared with the cardiovascular system. The first model of the lymphatic system is attributed to Reddy et al. and currently remains the only model of the entire system. 98 It is a simplified one-dimensional model of the entire lymphatic network that is based on the Navier–Stokes equations. Many ensuing studies have focused on specific areas of the lymphatic system, rather than the entire system as a whole.40–44
Analytical models of lymphangions in series (1–6 lymphangions) were developed to explore the optimal coordination of lymphangion contraction in both normal physiological conditions (input pressure less than output pressure) and edematous conditions (input pressure greater than output pressure). 40 Lymphedema is a condition that often occurs in breast cancer patients after an axillary node clearance, which results in altered fluidic conditions within the lymphatics. The single lymphangion model predicted high flow when subjected to edematous conditions and low flow when subjected to normal conditions. In the six lymphangion models it was found that the maximum flow occurs when the lymphangions pump almost synchronously (slight time delay between contractions, up to 3 seconds) and the flow reduces when the lymphangions pump simultaneously (contract at the same time).
A lumped parameter model for a chain of lymphangions (1–5 lymphangions) was created, which accounted for both the passive behavior of the vessels and the active vessel contractions. 41 The concept used in this investigation provided an insight closer to the physiological behavior of the lymphatics in vivo than any previous model. Sequential contraction of lymphangions was found to be much more efficient than synchronized contraction with regard to mean flow rate and adverse pressure overcome, in agreement with Venugopal et al.
Bertram et al. found that both intrinsic pumping and increased impedance add to the metabolic load of pumping, and the flow rate is greatly reduced if the lymphangion is not within range of its optimal operating point. When the model was subjected to a high pressure gradient, the pressure waveforms were sinusoidal in shape, and the pressure gradient in each lymphangion was small. However, interestingly, at higher flow rates these pressure waveforms took on more complex shapes as intermediate pressures exceeded downstream pressures. The advancements made by Bertram et al. have been improved upon further in ensuing studies.44,47,48
The model, initially developed in 2011, was modified to incorporate new physiological values, and with all the model parameters linked to experimental measurements, the group was able to elucidate whether all the experimental measurements were consistent. Bertram and colleagues found they were not, and then employed dimensional analysis to restore effective pumping in the model. The adverse pressure gradient across a series of lymphangions was maintained, whereas both the contraction timing and the lymphangion filling time were varied systematically to investigate valve function. 47 The properties of the valves were found to have a significant effect on system function.
Jamalian et al. identified which parameters have the greatest effects on system outcomes by carrying out a parameter sensitivity analysis based on the model developed by Bertram et al. 42 The authors were able to identify the number of lymphangions that resulted in the highest flow rate at each lymphangion length and the optimum number of lymphangions per vessel length based on a particular pressure difference. The first study to analyze a branching network of lymphatics since Reddy et al. in the 1970s is also attributed to this research group. 43 They built upon their previous lumped parameter model to create a bifurcating lymph network structure which consisted of multiple lymphangions (Fig. 2a).

Illustration of some of the various lymphatic system geometries modeled using numerical techniques.
They studied the effect of pumping coordination, external pressure and axial pressure difference on lymphatic pumping and what they found was that the effects of these functional parameters are highly interconnected. Their results demonstrated that effective pumping was highly influenced by both transaxial and transmural pressure differences. Thus, efficiency of the lymphatic pump is highly dependent on the coordination of the contraction wave. Additionally, the response of lymphatics to applied pressure conditions and signaling pathway manipulations have been modeled using mathematical simulations (Fig. 2b). 26 The mechanobiological control mechanisms described in this study adapt to varying fluidic environments to optimize lymph transport. For further information, a recent review summarizes the mechanobiological control mechanisms involved in lymph vessel function. 99
To date very few groups have modeled the initial lymphatics.35,51,100 The first estimates of the pressure drop required for lymph flow in the initial lymphatic networks (lymphatic capillaries) have only recently been obtained. 50 Anatomical measurements of rat mesenteric initial lymphatic networks were acquired and using a segmental Poiseuille flow model the pressure drops across both real and theoretical networks (approximately 1000 models) were calculated.
There are several aspects of lymphatic behavior that have not yet been addressed by the aforementioned lumped network models. Despite the assumptions and simplified modeling techniques used, the lumped models revealed very complicated behavior. The complexity of the lymph system is such that using discrete models as a first step allows a wide range of parameters to be investigated in a controlled manner. The complexity of these models can be increased over time to replicate in vivo conditions using more advanced computational methods to solve the space-dependant flow characteristics.
Computational modeling
Experimental characterization of lymphatic flow is extremely difficult; therefore, using computational fluid dynamics (CFD) may be the only feasible way to study certain aspects of it. To date, only a handful of studies have attempted to model the system using computational methods; however, the number of studies being published recently is increasing as summarized in Table 1. The lack of computational models developed presents a wealth of opportunities for researchers. A major concern with modeling flow in the lymphatics is the assumption of Poiseuille flow as the lymphatics have been shown to undergo large rapid contractions. 9 As a result, the validity of assuming Poiseuille flow to estimate wall shear stress (WSS) in a lymphangion was investigated. 34
A three-dimensional (3D) model of a radially expanding and contracting lymphangion was generated, and the WSS was analyzed throughout a contractile cycle. The applied wall motions resulted in high radial velocities; however, the WSS values recorded for each wall motion remained within 4% of quasistatic Poiseuille values. Thus, the study concluded that the assumption of Poiseuille flow is valid for the estimation of WSS in a lymphangion contractile cycle. Rahbar and Moore's model was the first 3D model published and the results provided valuable information; however, one limitation of the study is that it neglected to consider the potential effects lymphocytes and other cells have on the flow. The role the lymphatics play in spreading BCCs to distant locations within the body is by large unknown.
Several studies have identified the important role nitric oxide (NO) plays in controlling the lymphatic contractile cycle16,30,101 and advancements in imaging techniques have allowed more physiologically relevant models to be created. A realistic 3D model of a lymph vessel, created from confocal images, modeled the transport of NO within a rat mesenteric lymphatic vessel, again, this model was the first of its kind, (Fig. 2c). 28 Flow stagnation areas were found near the valve leaflets and NO concentration levels were shown to be sensitive to both imposed flow conditions and endothelial cell sensitivity.
The effect of sinus geometry and valve leaflets on the flow around lymphatic valves was investigated in a 3D parametric model based on images of isolated rat mesenteric lymphatics. 46 Similarly to their previous study, recirculation regions were recorded near the leaflets, and velocity values were found to be much lower than central lymph flow. Stagnant flow was observed, which may have an effect on cell transport in the region. The authors recommend that future studies should incorporate fluid–structure interaction (FSI) coupled with particle transport models. Additionally, the use of multiphase flow models could explore this microenvironment in even further detail.
Lymph node models have been created in which analysis of the flow patterns identified the parameters involved in lymph transport and exchange within the node.11,29,102 The first 3D model of a lymph node, (Fig. 2d), revealed that over 90% of the afferent lymph flow was directed around the periphery, through the subcapsular sinus and medulla. This behavior was also found in a subsequent study. 29 What is very interesting to note is that the WSS along the subcapsular sinus are within an order of magnitude of the lymphatics that lead to the afferent lymphatics of the lymph node. 11
Knowledge of these stresses and their magnitudes could be used in future experiments, where the various immune cells and pathogens contained within a lymph node are exposed to realistic conditions and their corresponding behavior investigated. This could potentially lead to more accurate in vitro cell culture and lymphatic metastasis models. The knowledge obtained from the studies discussed in this section has proved useful with regard to determining lymphatic function, but the models discussed contain a number of assumptions which warrant acknowledgement.
For example, many studies assumed the lymphatics were cylindrical in diameter when calculating the volume changes and uniform behavior was assumed for each lymphangion, which is not the case in vivo. As Table 1 illustrates, none of these studies considered the multiphase effects of cell flow on the fluidic environment. Hence the complexity of these models could be increased by employing multiphase modeling techniques such as Lagrangian tracking, the Discrete Element Method (DEM), or the Dynamic Fluid Body Interaction (DFBI) method. These types of models may provide an insight into the lymphatic conditions BCCs experience and how they alter normal flow conditions.
Recently, a study concluded that the mechanical forces generated by fluid flow regulate cellular behavior in terms of metastatic potential and proliferation. 97 The metabolic response of cells is associated with the shear stress magnitudes and gradients they are exposed to,7,93 therefore, identifying the local velocity gradients and shear stresses that BCCs are exposed to is crucial in determining their behavior under these specific flow conditions. Given the experimental difficulties of obtaining shear stress profiles on the surface of BCCs, in silico models offer several advantages with regard to obtaining detailed information on flow conditions and cell surface forces.
Numerical simulation of BCC motion in response to the hydrodynamic forces imposed by the lymphatics may facilitate in obtaining the shear stresses experienced by the cell membranes in this fluidic environment. To the author's knowledge, no experimental data are available on the response of BCCs to lymphatic flow conditions; however, numerous studies have investigated particle flow in microdevices, which could be leveraged to validate future numerical modeling studies in this area.
In situ studies
Information on lymphatic transport mechanisms is vital if the role lymphatics play in transporting BCCs is to be elucidated. High-speed imaging systems have measured flow velocities and shear stresses in lymphatics throughout an entire contraction cycle, which found that lymph velocity varies with phasic contractions in both magnitude and direction. 9 The work carried out by Dixon et al. provided the foundation for many subsequent studies.15,34,103
Lymphatics can contract up to ∼40%, over a characteristic period of 2–3 seconds, which results in contraction velocities of approximately 0.04 mm/s in rat mesenteric lymphatics. In these studies, lymphocytes were used as tracers to determine the lymph velocity and estimate WSS, and this approach assumes that lymphocytes faithfully follow the flow; however, this may not be the case. Lymphocytes are relatively large (5–8 μm) with regard to the vessels they travel in, and research has shown that large particles (>10% the channel width) tend to lag behind the fluid velocity. 104 Spatial resolution in these types of experiments also depends on the lymphocyte density, which can be quite low.
Recently, both the immediate and longer term response of lymphatics to edemagenic stress were investigated in rats that were infused with saline to induce edema. 105 Increases in flow rate, lymphatic contraction frequency, and fractional pump flow, which resulted in increased lymphatic contractile activity, were recorded. However, despite these large variations, statistically insignificant changes in vessel diameter were recorded. The increase in contraction frequency and the unchanged vessel diameter accounts for the increased WSS levels that were seen post infusion.
Average shear stresses in collecting lymphatics have been reported to be 0.64 ± 0.14 dyn/cm2, with peaks of 4–12 dyn/cm2, 103 and shear stresses on valve leaflets were up to three times greater than downstream in the lymphatic vessels. 106 However, in this study, shear stresses increased up to 40 dyn/cm2 due to the imposed edemagenic conditions. 105 Vessel contraction increased by 50% at 1 minute postinfusion and gradually decreased with time.
Lymphatic function has been shown to alter in response to induced inflammation in mesenteric lymphatics. 45 What is unique about this study is that the cellularity of the lymph in response to the inflammation was also measured. It was found that the relative density of cells in the lymph increased as the lymph flow decreased. The authors question whether changes in the hydrodynamics and resistance of the lymph nodes affect the work load that is required to move lymph through it. The results of the above studies suggest that lymphatics are particularly sensitive to mechanical stimuli, which allows them to adapt to altered fluid conditions.
Lymphatic contractility varies depending on the region of the lymphatic system. The strength and sensitivity of lymph pumps, in various regions of isolated rat lymphatics (thoracic duct, mesenteric, cervical, femoral lymphatics), were found to be predetermined by different hydrodynamic factors and outflow resistances. 107 The first study to investigate the timing of secondary lymphatic valve movements, in response to the lymphatic contraction cycle, is attributed to Davis et al. 10 Their results found that lymphatic vessel distention has a significant influence on the pressure gradient required for opening/closing the valves. Further work by this research group has also characterized the electrophysiological properties of rat mesenteric lymphatics. 30 Results of these experimental investigations have provided valuable inputs to many of the analytical models discussed previously.
In vivo imaging of lymphatics
Imaging of the lymphatics is challenging due to their size and location, as a result obtaining information on their velocities and contraction frequencies in vivo is very difficult. The first continuous in vivo measurement of lymph flow with temporal resolutions sufficient to quantify pulsatile dynamics was obtained using Doppler optical coherence tomography (OCT). 108 This work successfully recorded lymph volumetric flow rates and lymphatic contractions simultaneously in mice models and the accuracy of the method was compared with both in vitro and in vivo methods.
Real-time fluorescence imaging of MDA-MB-231 cells travelling in the lymphatics of immunodeficient mice was carried out to investigate the lymphogenic metastasis of breast cancer. 109 Migration of the BCCs in the lymphatics and hence, into the lymph nodes, where metastases were formed, was observed noninvasively, thus the entire architecture of the network was preserved, in contrast to other studies that investigate lymphatic flow.12,15,103 The events recorded in this study provide a more accurate indication of the physiology of breast cancer metastasis than previous work.
The metastasizing BCCs were monitored weekly to record their dissemination and the average BCC velocity was found to be 23 ± 11 μm/s, typical of lymphatic flow rates in mice. Before implantation into the mouse models, the BCCs were fluorescently labeled. In vivo fluorescent images of the BCCs were then captured to determine their velocity as they migrated through the lymphatics. The sequential monitoring discovered local metastases in the lymphatics after 8 weeks of implantation, which consequently resulted in reduced flow rates in the vessels.
Recently BCCs were injected into lymph nodes in rat models and the transport kinetics of a florescent dye were monitored so that the lymphatic flow from afferent lymphatics to lymph nodes could be quantified. 110 A significant difference between cancer-free and cancer-bearing lymph nodes was found, with the flow rates in cancer-bearing nodes being far lower than normal nodes. By using whole-organ imaging technology, a 3D representation of lymph nodes, with cellular resolution, have been obtained. 111 The benefit of these models includes a clearer picture of the node architecture and organization of the cells within the structure.
The emerging imaging techniques mentioned enable quantification of lymphatic function using high-resolution imaging, which will progress both research and clinical applications. While these experiments reveal important physiologically relevant results, the use of in vitro methods allow more control over factors that affect lymphatic function, which can lead to more detailed descriptions of the fluidic environment.
In vitro studies
Micro Particle Image Velocimetry (μPIV) techniques were employed to measure flow in lymphatics isolated from rat mesenteries. 8 This was the first study to use this measurement technique in lymphatics and its potential use in future experiments is very promising. The use of smaller tracer particles and μPIV processing techniques allowed the spatial resolution of the experiments to be significantly increased. The authors were able to record the flow around lymphatic valves and areas of flow disturbance were apparent (Fig. 3). The temporal resolution (2 μs) allowed for much greater detail than any other measurement technique to date.
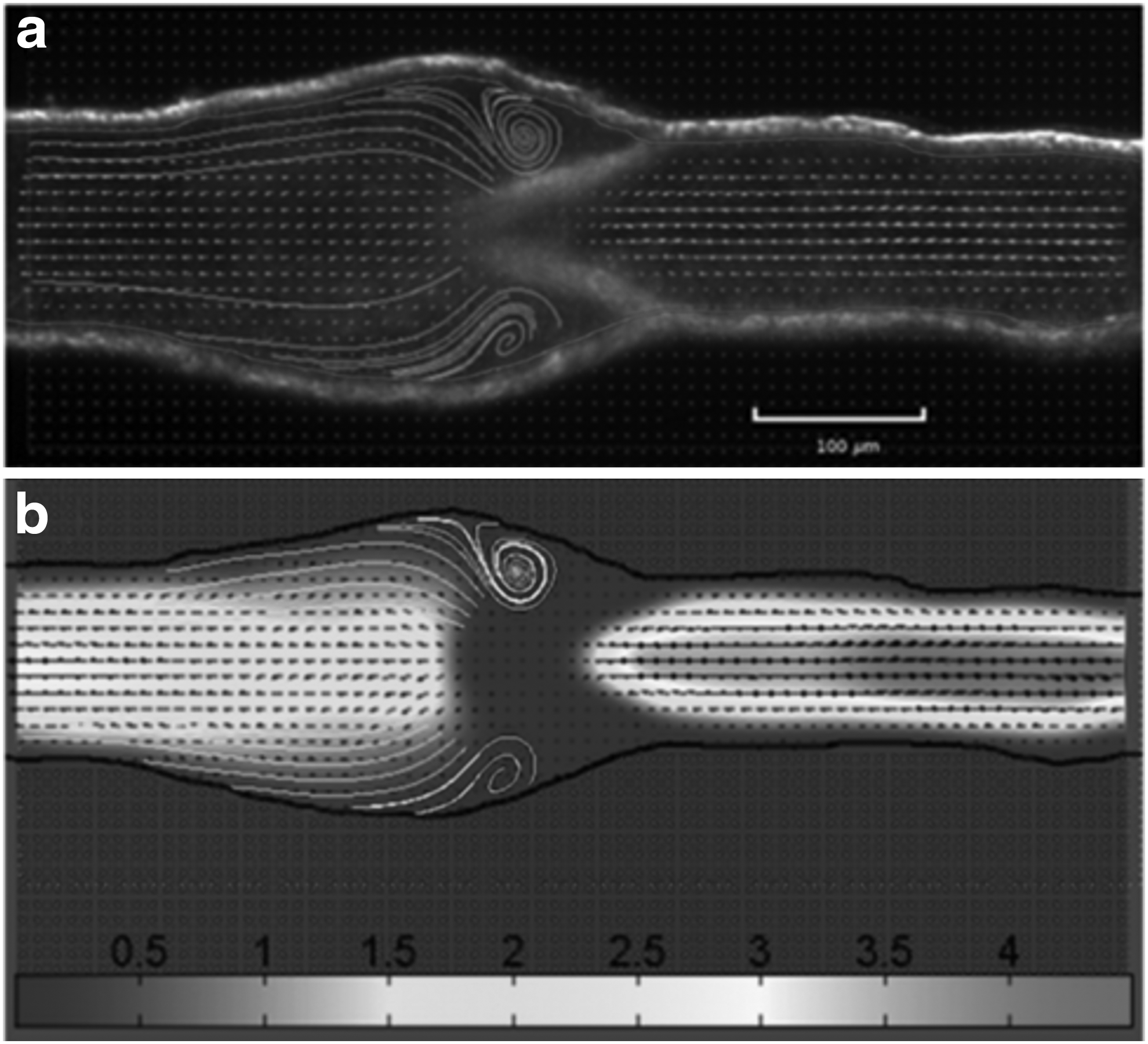
Flow around valve leaflets of a noncontracting rat lymphangion at an axial pressure gradient of −1 cm H2O at a transluminal pressure of 1 cm H2O obtained from μPIV measurements.
An in vitro model of the tumor–lymphatic environment, which allowed quantification of cell invasion and transmigration dynamics across lymphatic endothelium under varied biomechanical conditions, was created. 32 The focus of this study was to recapitulate the fluid forces acting on interstitial cells and the endothelium in the lymphatic environment since both of these are involved in cell trafficking. This model demonstrated that both luminal and transmural flows enhance the invasion and transmigration of MDA-MB-231 BCCs through the extracellular matrix. The authors show that luminal shear stress indirectly modulates tumor cell migration dynamics through its action on the endothelium. This model demonstrates the need for further research in this particular area by highlighting the complex relationship that exists between flow and BCC behavior.
As demonstrated throughout this review, the function of the lymphatic system, and the role it plays in cancer metastasis, is complicated. Most of our knowledge is derived from animal studies,10,15,30,47,53,112,113 therefore, it would not be unexpected if lymph transport in humans differs in some aspects from what has been found to date. Accurate measurement of lymph flow in vivo is very challenging; however, continuous advancements in imaging technologies will provide insights into the fluidic environment and flow-induced stresses BCCs are exposed to in this environment. With this knowledge, improvements can be made to current research modalities, exposing BCCs to a more realistic environment, leading to improved diagnostic techniques.
The response of BCCs to the physical environment in the lymphatics may be exploitable for the treatment of breast cancer metastasis. Preclinical and clinical studies show that lymph node metastases and primary tumors can respond differently to the same therapeutic regimen. 76 Drug development needs to account for the various microenvironments in which BCCs reside to improve efficacy of therapy. Recapitulation of the key aspects of the environment, such as quantifying the flow-induced surface forces BCC membranes are exposed to, may lead to improved cell culture experiments. Interdisciplinary research, in areas such as cell biology and microfluidics, is required to increase the knowledge base surrounding BCC metastasis before the end goal of discovering methods to control or manipulate the mechanisms governing BCC metastasis is achieved.
Dynamics of Particles in Confined Microenvironments
Microfluidic systems are regularly used to transport, sort, and analyze biological cells. Consequently, there is a considerable wealth of research that examines transport properties of particles and biological cells in microfluidic devices.58–70,114–122 Particles flowing in microchannels experience drag and lift forces that result in complex behavior, where particle velocities may lag or exceed the undisturbed flow velocity. Fluid forces due to shear, wall effects, and particle rotation scale differently depending on the local flow conditions with the effect that low concentration particles suspended in micro-flows move to equilibrium positions.
Segré and Silberberg were the first to report particle migration to focus on an annulus with a radius of ∼0.6 times the pipe radius. 117 This unexpected migration of particles to a stable dynamic equilibrium suggested that different types of forces acted on particles in Poiseuille flow. Later, theoretical analyses discovered that there are two opposing forces that dominate neutrally buoyant particles; the wall-induced lift force Fwall, and the shear gradient-induced lift force Fsheer gradient, (Fig. 4). The wall repulsion force pushes particles away from the channel walls as a result of the asymmetry of the corresponding wake vorticity distribution. The shear gradient lift force originates from the curvature of the velocity profile and causes the particles to migrate away from the central axis. The equilibrium position of particles is determined by the balance of these two forces as well as the particle and channel size.
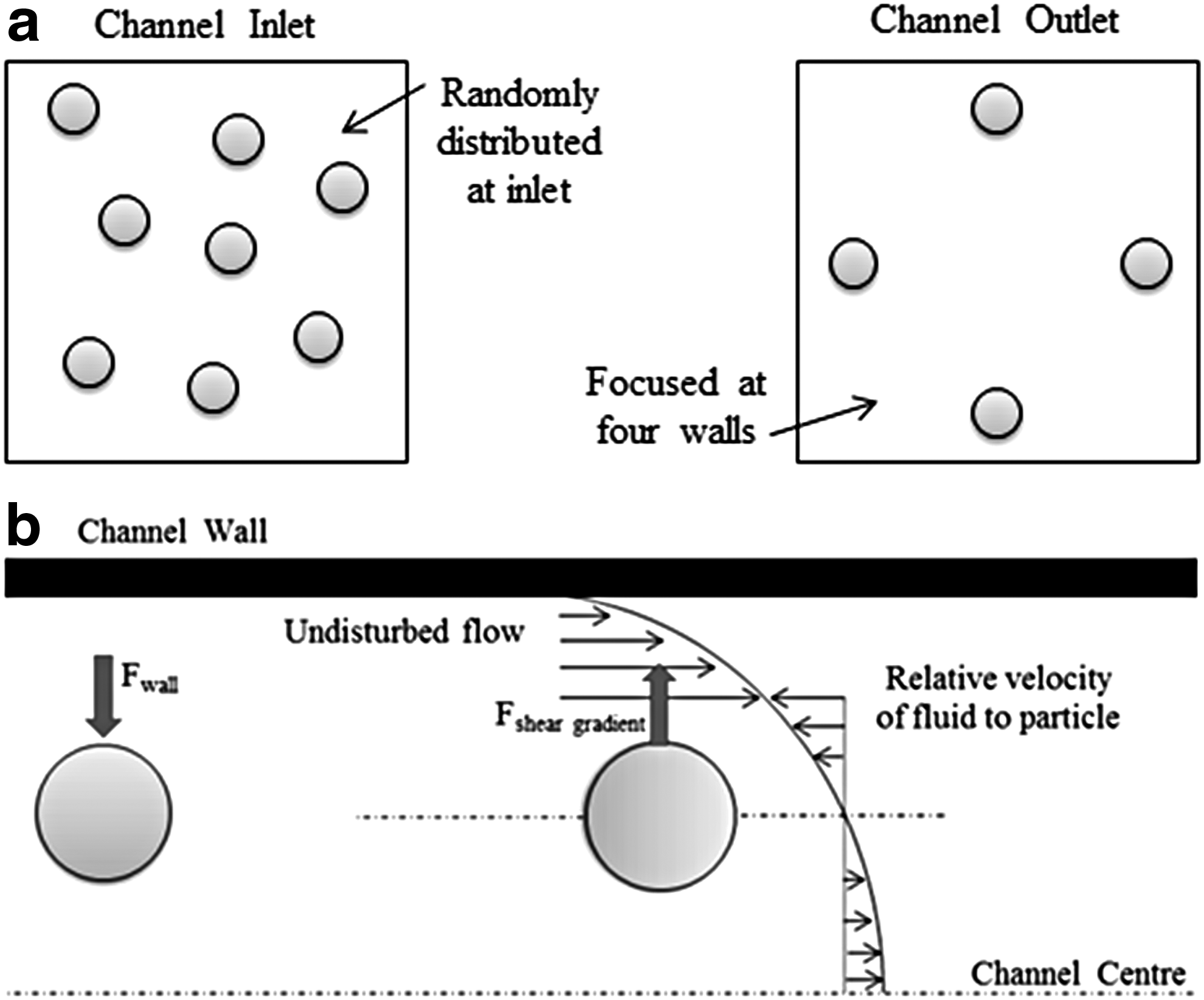
Despite the extensive body of experimental, theoretical, and computational studies that Segré–Silberberg's findings initiated, a complete insight into the inertial migration of particles is not fully understood.123,124 Table 2 summarizes a number of studies that have investigated particle/cell behavior under low flow rates (Re < 500) in microchannels in an attempt to elucidate their behavior. A number of studies summarized in Table 2 are discussed to provide an overview of the events that occur during particle/cell advection in microflows.
A description of the particles/cells that were tested is provided along with information on the flow rates and a description of the channel geometry.
It has been shown that migration does not occur when the particle size is small compared with the channel height. 63 The critical Re at which the equilibrium positions start to form is estimated to be 20–30.125,126 However, when the channel height approaches the same order of magnitude as the particle diameter, the walls influence their advection and assuming that the particles travel with the local fluid velocity is no longer valid. Further research is required to fully characterize the motion of single particles in Poiseuille flows. 127 The smallest dimensions of the lymphatics are not much larger than the BCCs that are being transported in them; as a result, there is a highly dynamical and strongly interdependant relationship between them.
Dilute suspensions of large (∼10% the channel width) particles adhere to the following sequence of events when flowing in a square microchannel; at low Re (Re ∼100), the particles migrate toward the channel walls, where they are evenly spaced in the flow direction, whereas at higher Re (Re >700) values, the uniform spacing no longer remains and particles tend to cluster together at additional equilibrium positions located near the channel center. 128
Staben et al. formulated a boundary integral solution technique for a particle flowing between two parallel plane walls in Poiseuille flow. They found that the average particle velocity for a uniform distribution of particles over the channel depth was greater than the average fluid velocity for the range dp/H < 0.82, where dp is the particle diameter and H is the channel height. However, this result can be explained by the exclusion of particles from the near wall regions, where the fluid velocity is slowest. When dp/H > 0.82, the particle velocities are significantly affected by wall interactions at all depths such that the average particle velocity is less than the average fluid velocity. 104 When the particle size is large compared with the channel size, the particle velocity decreases due to the retarding effects of the nearby walls.
In the case of BCCs, which are large when compared with the lymphatics, the effect the BCCs have on the local fluid velocity at the flow rates experienced in vivo has not been established. In a later study, particle tracking experiments were carried out to validate the boundary integral simulations. 127 Again they found that the measured and predicted velocities show that a particle's velocity lags behind the main fluid velocity when the particle is larger and/or closer to the channel walls.
Figure 5a shows that small particles travel at the same velocity as the local fluid, except in regions closest to the walls. Increasing particle size results in a decrease in the particle velocity due to the retarding effects of the nearby walls, as seen by the shifting of the velocity curves to the left with increasing particle size. When the ratio of dp/H is large, the general assumption that the particle velocity equals the undisturbed flow velocity does not hold and they recommend it should be re-examined.

For dp/H << 1, Di Carlo et al. observed that the particle equilibrium positions approached ∼0.6 times the channel radius, in agreement with Segré and Silberberg, and they also observed that the particle velocities were not the same as the unperturbed fluid mean at their center point. 129 The low Re values that were investigated in these studies remain considerably larger than the values present in the lymphatics (Re < 1). To the author's knowledge, an understanding of the behavior of large particles/cells under these flow conditions has yet to be elucidated. Recent work carried out by Liu et al. proposes a two-stage particle migration based on numerical predictions which is in agreement with previous experimental observations.60,130 The results reveal the general dependence of equilibrium positions on dp/H and Re.
Kim and Yoo investigated the lateral migration of neutrally buoyant particles in square channels (0.06 ≤ Re ≤ 58.65). At low Re, they observed that the particles (6 μm and 10 μm) were distributed somewhat uniformly across the square microchannels (W = 87.5 μm and 140.9 μm), concluding that particle migration is not seen at such small Re. 118 However, as they increased Re they observed particle migration away from the channel center, as illustrated in Fig. 5b. These results are in direct agreement with previous work. 126
As demonstrated from the above literature, a complete understanding of the mechanisms behind particle migration has yet to be exposed, despite the increased research in this area. The majority of the studies examine the migratory behavior of very small particles (<10% the channel width) under typical inertial focusing flow rates (30< Re <300). However, very limited data are available on the response of large particles (>10% the channel width) to the low Re flow present in the lymphatics. A more complete understanding of real-world particle migration (e.g., cancer cells, bacteria) requires additional knowledge of how particle shape, deformability, and fluid properties contribute to the overall motion.
Cancer Cell Advection in Microflows
The theoretical understanding of the inertial migration of cancer cells is poor when compared with rigid particles. Cancer cells vary in size, shape, and morphological properties and have been shown to be much more deformable compared with the normal cells of the same tissue.116,131,132 Deformable cells moving in confined flow introduce nonlinearities in the system, which causes cross-stream migration. The direction of the deformability-induced lift force is toward the channel center, owing to the parabolic nature of the Poiseuille velocity profile. 124 This deformability-induced lift results in equilibrium positions closer to the center of the channel in comparison with rigid particles. 133
At low Re (Re = 21–42), Hur et al. describe the potential for migration due to deformation as a potential means to sort cells. 116 This enables similar sized cells to be separated based on their morphological response to the flow. The lateral migration of droplets, particles, and a number of cell types, including MCF-7 cells were compared. The results revealed that BCCs with increased metastatic potential and larger BCCs migrated closer to the channel centerline than that of benign BCCs despite their similar cell size. They found that in general, increasing cell size or deformability causes the equilibrium positions to shift away from the wall, similarly to previous work. 134 Bacteria and cancer cells can be filtered using these separation techniques.70,114,121
Additionally, the migration behavior of nonspherical particles has been shown to be largely influenced by the rotational diameter.114,135 Tanaka et al. examined the inertial migration of MDA-MB-231 cells and particles as a means of identifying CTCs in high hematocrit bloodflow. 68 It was found that the BCCs required a longer channel length (almost double) to reach an equilibrium state compared with the rigid particles. These differences were attributed to the distribution of BCC size and interestingly, cell deformation was deemed not be a major contributing factor. Small variations in BCC size resulted in large differences in the migration length.
Lim et al. used Particle Tracking Velocimetry techniques to observe the behavior of particles, white blood cells (WBCs), and prostate cancer cells in physiological saline and blood. 119 Inertial focusing behavior of the particles in diluted and whole blood solutions (HCT = 45%) was observed. They found that despite the similarity in diameter between the WBCs and the particles, WBC focusing was visibly weaker. The decrease in WBC focusing was attributed to the smaller WBCs found unfocused at vertical positions near the channel wall. This is a very interesting finding, if one considers that BCCs can vary in size from 10 μm to 40 μm, one would expect that the cells behavior in the lymphatics would depend on their size; however, there is little or no evidence in the literature on how BCCs affect lymph flow or what the consequences with regard to cancer metastasis are.
The mechanical properties of cells can change significantly in the context of disease. The stiffness of cancer cells is drastically reduced when compared with cells of the same origin. Furthermore, increasing metastatic potential is associated with decreasing single cell stiffness. To date, the effect these properties have on the flow conditions in the vessels transporting them has not been investigated. Taking all of the above factors into account suggests that the response of BCCs to the fluidic forces they are subjected to depends largely on the cell's geometrical and morphological properties.
Future Perspectives
Despite the amount of literature available on the biological/signaling aspect of how BCCs enter the lymphatics, the events that occur subsequently in transporting them have been relatively unexplored. Thus, a prominent area for future research will be the mechanics that govern BCC advection in the lymphatics. The use of in vitro models will provide controlled environments in which variables can be systematically analyzed. The outputs can then be used to create more biologically relevant models in the future. The intersection between biological modeling of lymphatic metastasis and the mechanics of BCC transport will provide significant advancements in the area of lymphatic research and biology.
More comprehensive theoretical and experimental techniques are needed to quantify the interaction between cells/particles and lymph flow, under various physiological conditions, resulting in a more concrete basis for numerical simulation, with applications lending themselves to numerous types of biological cell flows. A major challenge that remains is the successful translation of experimental measurements to an in vivo setting. The lymphatics are not easily visualized and this technical limitation has greatly hindered research advancements. The majority of the studies discussed lack of the spatial resolution needed to investigate specific flow patterns within the lymphatics. New investigative techniques need to be developed or current imaging modalities modified so that the flow velocity in small lymphatics can be accurately recorded.
Numerous reviews have highlighted the need for further research in these areas.5,136,137 More sophisticated imaging techniques are now becoming available and μPIV show great promise with regard to accurately capturing flow patterns in the lymphatics. The ideal investigation would incorporate a combination of microfluidic and imaging technologies to analyze the behavior of multiphase flow in physiologically relevant models, where all experimental inputs have been obtained from in vivo measurements. The recent advancements in imaging capabilities will lead to an increase in the amount of physiologically relevant models generated, in which CFD analysis can predict in vivo conditions.
As evidenced by the scarcity of studies devoted to the topic, numerical modeling of large cells in microvessels presents a number of difficulties, future efforts are required to focus on research needs such as biomicrofluidics. All of the computational models to date have assumed that lymph is a continuous, single-component fluid, and the multiphase effects of cells suspended in the flow have been ignored. The introduction of cells into the current models would provide a valuable comparison. The use of multiphase modeling techniques, such as the DFBI method, or employing FSI methods, could potentially lead to some interesting findings with regard to BCC flow in the lymphatics. The results of this review serve to motivate experimental work focused on linking fluidic conditions to BCC dynamics associated with metastasis.
Conclusions
This review highlights the significant advances that have been made over the past two decades to improve our understanding of the lymph system, with special attention paid to the lymphatic fluidic environment, which metastasizing BCCs are exposed to. To date, the dynamics of BCC transport within the lymphatics has not been quantified. The fundamental fluid dynamics that control the transport of these cells has yet to be established, at least partially, due to the lack of biomechanically relevant models. Numerous studies have been carried out to characterize lymphatic flow; however, none of these studies addresses the physical effect of BCCs.
One major conclusion that can be drawn as a result of reviewing this literature is that, despite the increase in the number of studies being carried out in this area, there remains a vast wealth of information that has yet to be discovered. There is a critical need to develop more complex models to accurately capture the flow phenomena present. Establishing comprehensive connections between the experimental and numerical studies will provide novel and powerful developments for breast cancer diagnostics in the future.
Footnotes
Acknowledgments
This study was supported by the Irish Research Council and the Mid-Western Cancer Foundation.
Author Disclosure Statement
No competing financial interests exist.