
Abstract
Abstract
Background:
The initial periods of increased flow inside lymphatic vessels demonstrate specific temporary patterns of self-tuning of lymphatic vessel contractility that are heterogeneous across regional lymphatic networks. The current literature primarily refers to the immediate and fast reactions of the lymphangions to increases in basal flow. Until now, there were no available data on how the lymphatic vessels react to comparatively longer periods of imposed flow.
Methods and Results:
In this study, we measured and analyzed the contractility of the rat thoracic duct segments, isolated, cannulated, and pressurized at 3 cm H2O at no imposed flow conditions and during 4 hours of imposed flow (constant transaxial pressure gradient of 2 cm H2O). We found the development of a progressing lymphatic tonic relaxation and inhibition of the lymphatic contraction frequency over 4 hours of imposed flow. After a short initial decrease, lymphatic phasic contraction amplitude rose significantly during the first hour of imposed flow, and it demonstrated a trend to return toward control levels after 3 hours of imposed flow. As a result, the fractional pump flow (active lymph pumping per minute) of isolated thoracic duct segments reached and maintained a statistically significant decrease (from control no-flow conditions) at the end of the third hour of imposed flow.
Conclusions:
Our new findings provide a better understanding of how lymphatic contractility changes during the development of prolonged periods of steady lymph flow. The latter may occur during the initial phases of development of an inflammatory-related tissue edema.
Introduction
L
In the thoracic duct, such short-lasting phasic pulses of shear stress-dependent NO release are shown to play an important regulatory role to ease lymphatic diastolic filling and to keep lymphatic vessels in an efficient mode of pumping during periods with low physiological levels of basal lymph flow. 6 These brisk releases of NO molecule synchronized with each lymphatic pumping cycle, which peak during diastole, and enhanced lymphatic filling and the subsequent contraction.6,8,14 At the same time, the increases in overall lymph production and thus lymph flow (imposed flow) induce fast and relatively large increases in the levels of NO, the latter causing fast relaxation and inhibition of lymphatic contractions.3,15–17
The inhibitory influences of large, rapid releases of NO (not dependent on the phasic contractions) are believed to prevent unnecessary expenditures of energy to pump lymph, whereas passive driving forces (often related to increased lymph formation) are sufficient to maintain lymph flow. 3 The inhibition of phasic lymphatic contractions in such cases helps to avoid increases in the resistance of the phasically contracting lymphatic vessels to flow with an existing passive extrinsic lymph flow.3,18,19 These large increases in steady-state NO will overwhelm the contractile efficiency-supportive effects of the smaller-magnitude NO spikes that are associated with the phasic lymph flow during the active lymphatic pumping.
Such complex relationships between different patterns and magnitudes of lymph flow with variable lymph NO make interpreting the effects of the genetic removal of basal NO in lymphatics difficult. Indeed, it appears that the effects of genetic ablation of endothelial nitric oxide synthase (eNOS)-derived NO in eNOS knockout mice have been interpreted without in-depth consideration of the duality of physiological roles of NO in collecting lymphatics. 20 Moreover, the lymph flow-dependent roles of NO in the regulation of lymphatic contractility appears to be even more complicated if regional 5 and interspecies21,22 differences in lymphatic contractility are taken into account. An additional level of complexity of the flow-induced adaptive reactions of contracting lymphatic vessels was brought by recent studies on the regional roles of lymphatic endothelium-derived relaxing factors (EDRFs) other than NO.10,11
The initial periods of imposed flow inside collecting lymphatic vessels demonstrate specific temporary patterns of self-tuning of lymphatic vessel contractility 3 and are heterogeneous in different regional lymphatic networks. 5 However, currently, all of these effects of increased flow in the isolated lymphatic vessels were studied in experiments with imposed flow kept for up to about 15 minutes. Therefore, the data and conclusions obtained in those studies and studies performed more recently 23 reflect the immediate and fast reactivity of the lymphangions to increases in imposed flow. Those experimental conditions imitate the initial phase of periods of increased lymph flow during different physiological events (increased gut adsorption, skeletal muscle activity, etc.) as well as during the increases of lymph formation seen in pathologic processes with the corresponding development of an acute tissue swelling.
However, until now, there are no data available that describe how the lymphatic vessels react to comparatively longer periods of imposed flow. Careful evaluations of lymphatic contractility and pumping in such conditions have never been published. We believe that studies with conditions of steady imposed flow created over several hours would provide important information on how lymphatic vessels behave in situations when the flow inside of them is permanently increased. Thus, we performed experiments with 4 hour periods of steady, moderate imposed flow within isolated segments of rat thoracic duct, therefore imitating the initial periods of long-term elevations of basal lymph flow.
Materials and Methods
Animals and surgery
We examined the contractile activity of thoracic ducts from 16 male Sprague-Dawley rats (weighing between 300 and 400 g). All animal procedures for the current studies were reviewed and approved by our Institutional Animal Care and Use Committee and were in accordance with federal and local regulations.
To isolate the thoracic duct, rats were euthanized with pentobarbital (120 mg/kg body weight intraperitoneally). Then, the animal was positioned on its back; the ventral chest wall was opened by lateral incision; and the sternum and approximately half of the ribs were excised. The inferior vena cavae was ligated and cut close to the diaphragm. The lungs and heart were set to the left side of the animal so as to expose the thoracic duct between the aorta and vertebral column. The thoracic duct was then carefully cleared of all surrounding tissues by using a dissecting microscope. Extreme caution was used to not grab or pinch the thoracic duct at any time, thereby reducing the likelihood of damage. The area of interest was kept moist for the period of dissection by using the standard Dulbecco's phosphate-buffered saline (catalog No. 14040-133; Invitrogen Corp., Carlsbad, CA).
Sections of thoracic duct 1–2 cm long containing one valve were dissected and used for experiments. Throughout the data collection periods of the experiments, we measured and recorded the diameters of the thoracic duct sections used for these studies. At a transmural pressure of 3 cm H2O and absence of the imposed flow, the average thoracic duct diastolic diameters were 620 ± 35 μm.
Isolated thoracic duct procedures, experimental techniques, and protocol for functional tests
Once the segment of the thoracic duct was exteriorized, it was transferred to an isolated vessel chamber (modified Living Systems Instrumentation single-vessel chamber model CH/1). The chamber was filled with albumin-physiological salt solution (APSS; in mM: 145.0 NaCl, 4.7 KCl, 2.0 CaCl2, 1.2 MgSO4, 1.2 NaH2PO4, 5.0 Dextrose, 2.0 Sodium Pyruvate, 0.02 EDTA, 3.0 MOPS, and 10 g/L bovine serum albumin) pH adjusted to 7.36 at 38°C. The isolated thoracic duct segment was cannulated and tied onto two carefully matched glass pipettes (500–550 μm). Great care was used to prepare and select pairs of resistance-matched pipettes for these experiments as described in our previous studies.5,6
The inflow and outflow pipettes were connected to independently adjustable pressure reservoirs filled with APSS. Care was taken to ensure that there were no air bubbles in the tubing or the pipettes. Once the vessel was cannulated, a slight positive transmural pressure (2–3 cm H2O) was applied to detect leaks and to ensure that the vessel was undamaged and untwisted. The vessel was set to its approximate in situ length and positioned just above the glass coverslip chamber bottom.
The chamber was transferred to the stage of an inverted microscope. The vessel was set to an equilibration transmural pressure of 3 cm H2O and warmed to 38°C over 15–20 minutes. Once tone alone or tone and spontaneous contractions were observed, the vessel was allowed to equilibrate at 3 cm H2O for another 30 minutes. A video camera, hi-resolution monitor, and DVD/HDD recorder were used to observe and record the lymphatic segments continuously in all experiments.
At the beginning of every experiment, we evaluated the pressure-induced contractile responses of the thoracic duct. Lymphatic segments were exposed to a standard range of transmural pressures: 1, 3, and 5 cm H2O for 5 minutes at each pressure. We chose this set of transmural pressures as a control test to confirm that existing contractile activity in isolated thoracic duct segments used for the study was normal and matched our previously obtained average parameters of contractile activity of such preparations.5,6,24 Only lymphatic segments displaying normal stretch-dependent contractile responses (data not shown) were used to collect data.
The rat thoracic duct displays maximal active pumping at 3 cm H2O,5,6,24 and a modest imposed flow (2 cm H2O axial pressure gradient) inhibits its contractions moderately.5,24 Therefore, we selected these conditions—transmural pressure of 3 cm H2O and imposed flow gradient of 2 cm H2O—for our current experiments to investigate the influence of steady imposed flow on thoracic duct contractility during 4 hours of imposed flow. After the control pressure tests were completed, we set a transmural pressure of 3 cm H2O on the thoracic duct segments and recorded their contractile activity for 5 minutes. The average parameters of lymphatic contractions for the first 5 minutes were used as the control/“no flow” data set.
Immediately after completion of the control/“no flow” period, the thoracic duct segments were exposed to an imposed flow axial gradient of 2 cm H2O for 4 hours by setting the inflow pressure to 4 cm H2O and outflow pressure to 2 cm H2O. This technique to create an imposed flow was previously developed in isolated lymphatic preparations.3,5,6,9,24,25 In the 4 hour experiments, we carefully monitored levels of fluid in input and output reservoirs to keep these levels unchanged over the whole 4 hours of the experiment.
We measured the contractile activity in specimens for 5-minute intervals through the duration of the 4 hour experiment: initially—after 5 minutes of imposed flow (averages between minutes 5 and 10 of existence of an imposed flow—labeled as point “5 min” in Figures), then for the last 5 minutes every half hour of the imposed flow over 4 hours (i.e., 5-minute interval started at time point 25 minutes, 5-minute interval started at time point 55 minutes, and so on until the last 5-minute interval started at time point 3 hours 55 minutes). At the end of each experiment in every segment, the passive (relaxed) diameter was measured at level of transmural pressure of 3 cm H2O after the vessels were exposed to a nominally calcium-free, EDTA (3.0 mM) supplemented APSS for 15 minutes.
Experimental conditions in our studies were typical to those similarly used for tests of thoracic duct segments that were cultured for 3–12 days with confirmed absence of the negative effects on lymphatic contractility induced by experimental conditions. 21
Data analysis and statistics for isolated vessel experiments
The lymphatic diameters were tracked from the DVD records of experiments by using “Vessel Track” software developed and generously provided by Professor Michael J. Davis.26,27 Briefly, the outer lymphatic diameters were measured from the DVD record at a tracking frequency of 30/s. We used cardiac pump analogies to define systole and diastole in reference to the lymphatic contractile cycle,5,28,29 and the end-diastolic and end-systolic points in the diameter tracings were recorded for each selected 5-minute interval of “no flow” and imposed flow gradient of 2 cm H2O.
From the lymphatic end-diastolic and end-systolic diameters (EDD and ESD), the following lymph pump parameters were calculated: lymphatic tone index (the difference between the passive lymphatic diameter in Ca-free APSS and EDD, expressed as a percentage of the passive lymphatic diameter in Ca-free APSS), contraction amplitude (the difference between the diastolic and systolic diameters, expressed as a percentage of difference between EDD and ESD normalized to passive lymphatic diameter), contraction frequency, ejection fraction (EF, the fraction of end-diastolic volume ejected during the single lymphatic contraction, calculated by using formula EF = (EDD2 − ESD2)/EDD2), 28 and fractional pump flow (FPF) (an index of lymph pump flow, calculated as the EF times the contraction frequency).
To compare the changes in diameters during the lymphatic contractile cycle, the diastolic and systolic diameters were normalized to the passive lymphatic diameters in Ca-free APSS at the corresponding transmural pressure because of the anatomical variations between segments of thoracic duct. In all experiments, only one thoracic duct segment was used from each animal for the experiment (totaling n = 16).
Data were analyzed with a linear mixed-model approach, with time as a fixed factor and experiment as a random factor. REML (residual maximum likelihood estimation) was used to estimate the covariance parameters. Models with random intercepts, random effects, and heterogeneous residual variance were evaluated hierarchically for each variable, and the most appropriate was chosen based on likelihood ratio testing of the nested models. All variables tested were found to vary significantly over time, with p-values ranging from <0.0001 to 0.0064. The significance of changes in each variable at later times compared with time 0 was tested directly by rerunning the model with time as a categorical fixed factor and time 0 as the reference category. Statistical analysis was performed with Stata 14 for Windows (StataCorp LP, College Station, TX).
Results
Figure 1 demonstrates the progressing relaxation in rat thoracic duct over the 4 hours of imposed flow gradient of 2 cm H2O. This decrease in thoracic duct tone became statistically different from tone at no flow conditions at the end of the second hour of the presence of imposed flow. Finally, at the end of the fourth hour of imposed flow, the average tone was 61% lower than in the thoracic duct during control conditions (no imposed flow). These values were 1.9% ± 0.4% (4 hours of flow) and 4.7% ± 0.7% (no flow) tone correspondingly.
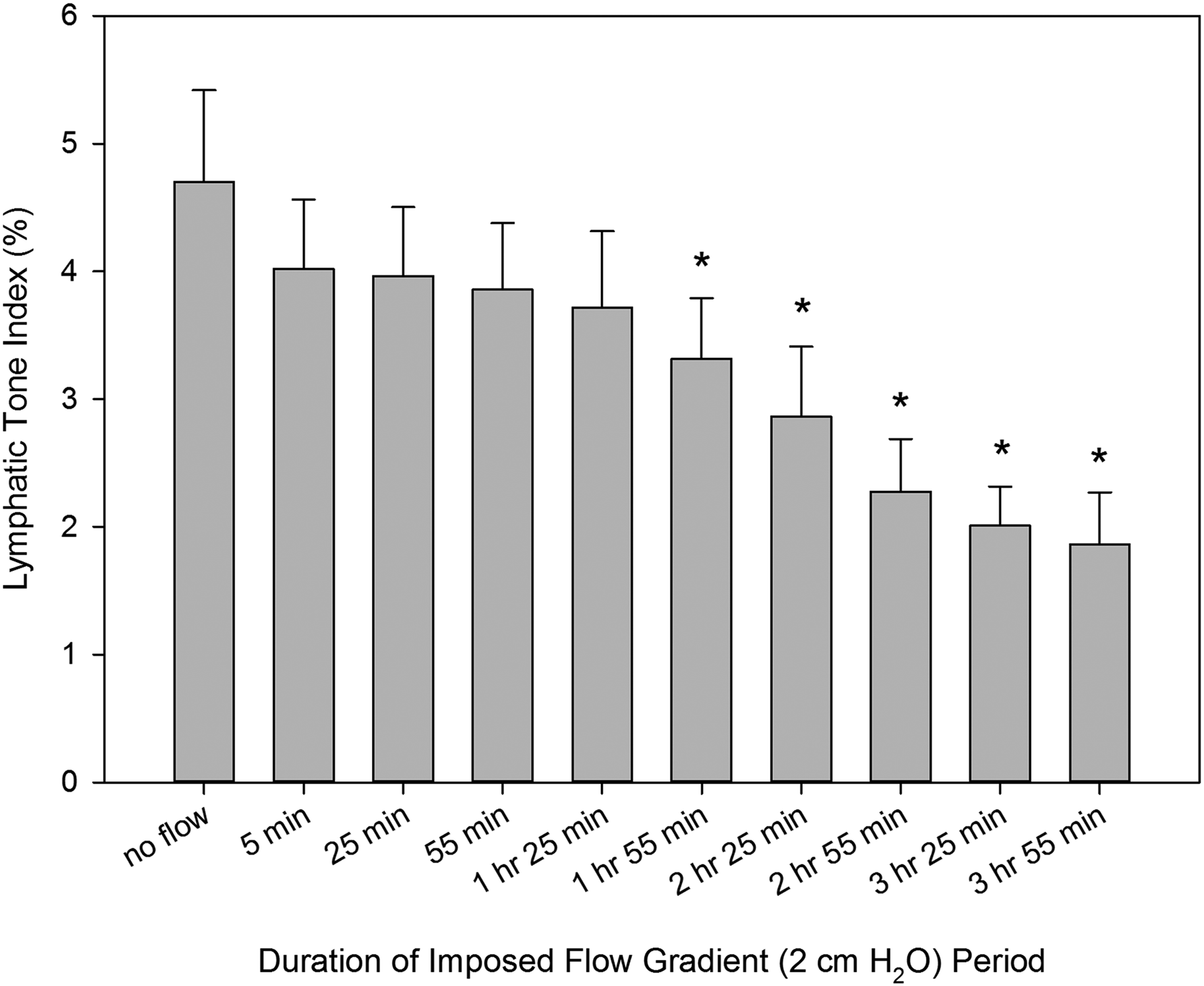
Changes in lymphatic tone of the rat thoracic duct during imposed flow applied for 4 hours. *Indicates significant differences (p values ranging from <0.0001 to 0.0064) between lymphatic tone index at control conditions (no imposed flow) and at corresponding time points after creation of an imposed flow.
Changes in the phasic contraction amplitude over the 4 hours of the imposed flow were more complicated and are illustrated in Figure 2. After an initial period of slight but not-significant decrease of contraction amplitude from 13.3% ± 1.7% at no flow conditions to 11.4% ± 1.4% at the end of the initial 5 minutes of imposed flow, we observed the development of a prolonged positive inotropic effect over the initial 3 hours of imposed flow. Contraction amplitude maximally increased to 20.1% ± 2.5% at the end of the second hour of the presence of these conditions. During the fourth hour of imposed flow, the contraction amplitude demonstrated a trend to return toward control levels.
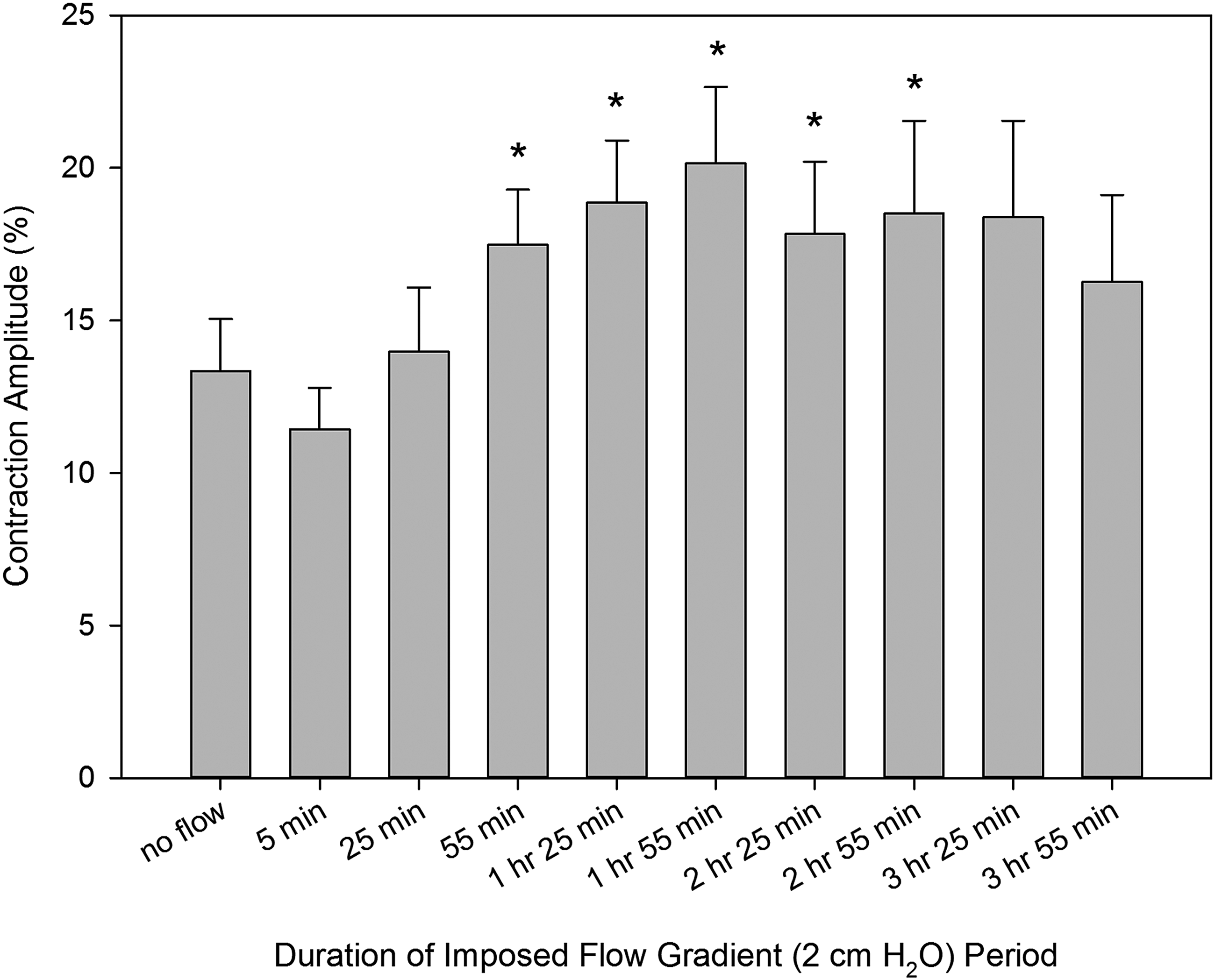
Changes in contraction amplitude of the rat thoracic duct during imposed flow applied for 4 hours. *Indicates significant differences (p values ranging from <0.0001 to 0.0064) between contraction amplitude at control conditions (no imposed flow) and at corresponding time points after creation of an imposed flow.
At the same time, a robust, unidirectional decrease in thoracic duct contraction frequency occurred over the duration of the 4-hour period of imposed flow. Figure 3 demonstrates the consistent development of the negative chronotropic effect of imposed flow during the whole 4 hours of imposed flow. This decreased contraction frequency reached and maintained statistically significant differences (from control no-flow conditions) at the end of the first hour of imposed flow. At the end of the fourth hour of imposed flow, the average contraction frequency was 37% lower than during control conditions (no imposed flow). These values were 6.4 ± 1.1/min and 10.1 ± 1.3% correspondingly.
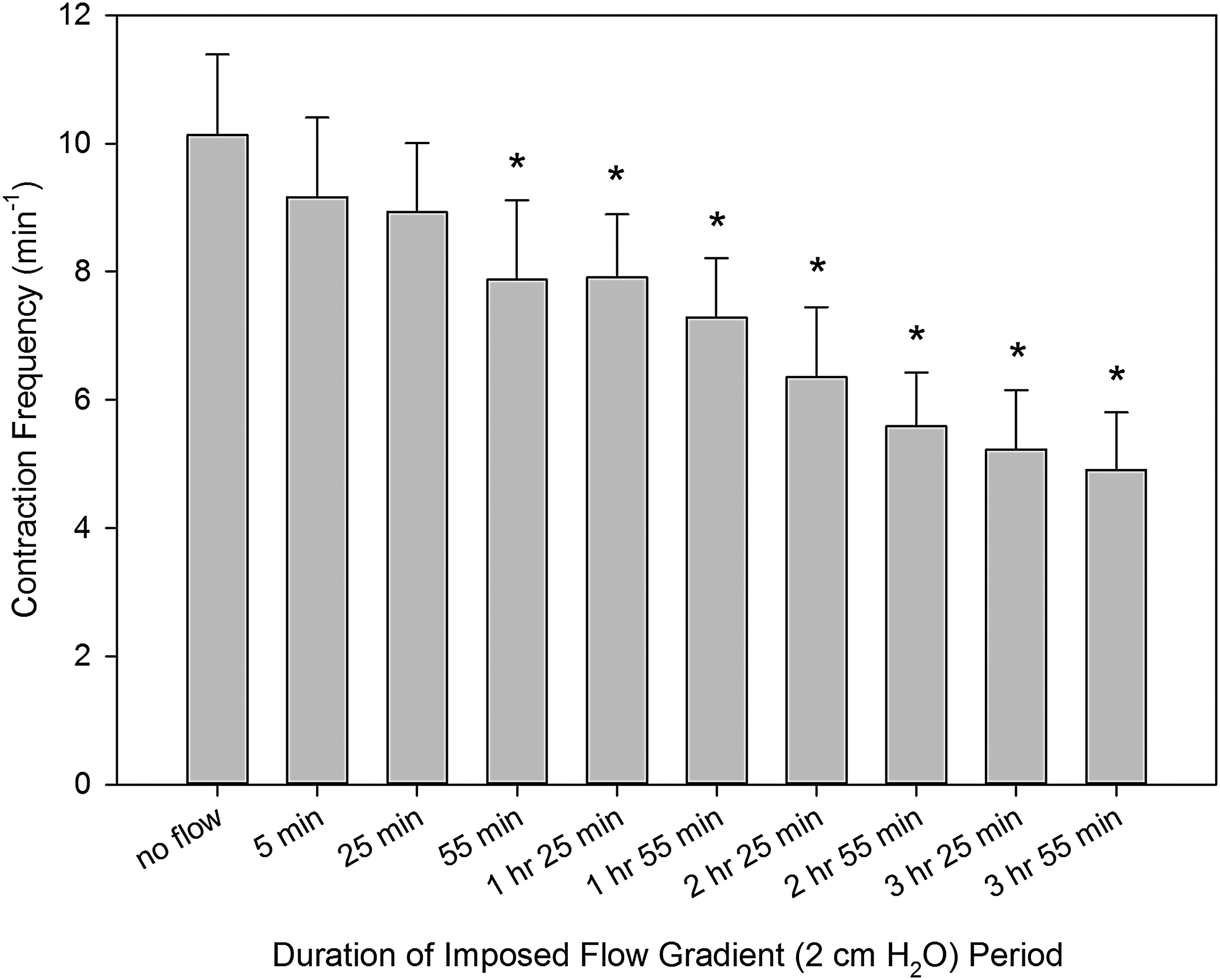
Changes in contraction frequency of the rat thoracic duct during imposed flow applied for 4 hours. *Indicates significant differences (p values ranging from <0.0001 to 0.0064) between contraction frequency at control conditions (no imposed flow) and at corresponding time points after creation of an imposed flow.
As a result of the specificity in inotropic and chronotropic effects of prolonged imposed flow on the isolated thoracic duct segments described earlier, their FPF (active lymph pumping per minute) reached and maintained a statistically significant decrease (from control no-flow conditions) at the end of the third hour of imposed flow. Figure 4 illustrates these findings; it demonstrates that after an initial slight decrease in FPF, then thoracic duct segments tended to maintain the same level of pumping over the first two first hours of the imposed flow. During the third and fourth hours, FPF started to fall. As a result, FPF in control no-flow conditions on average was 2.7 ± 0.4; then, after 4 hours of imposed flow, it dropped to 1.7 ± 0.4.
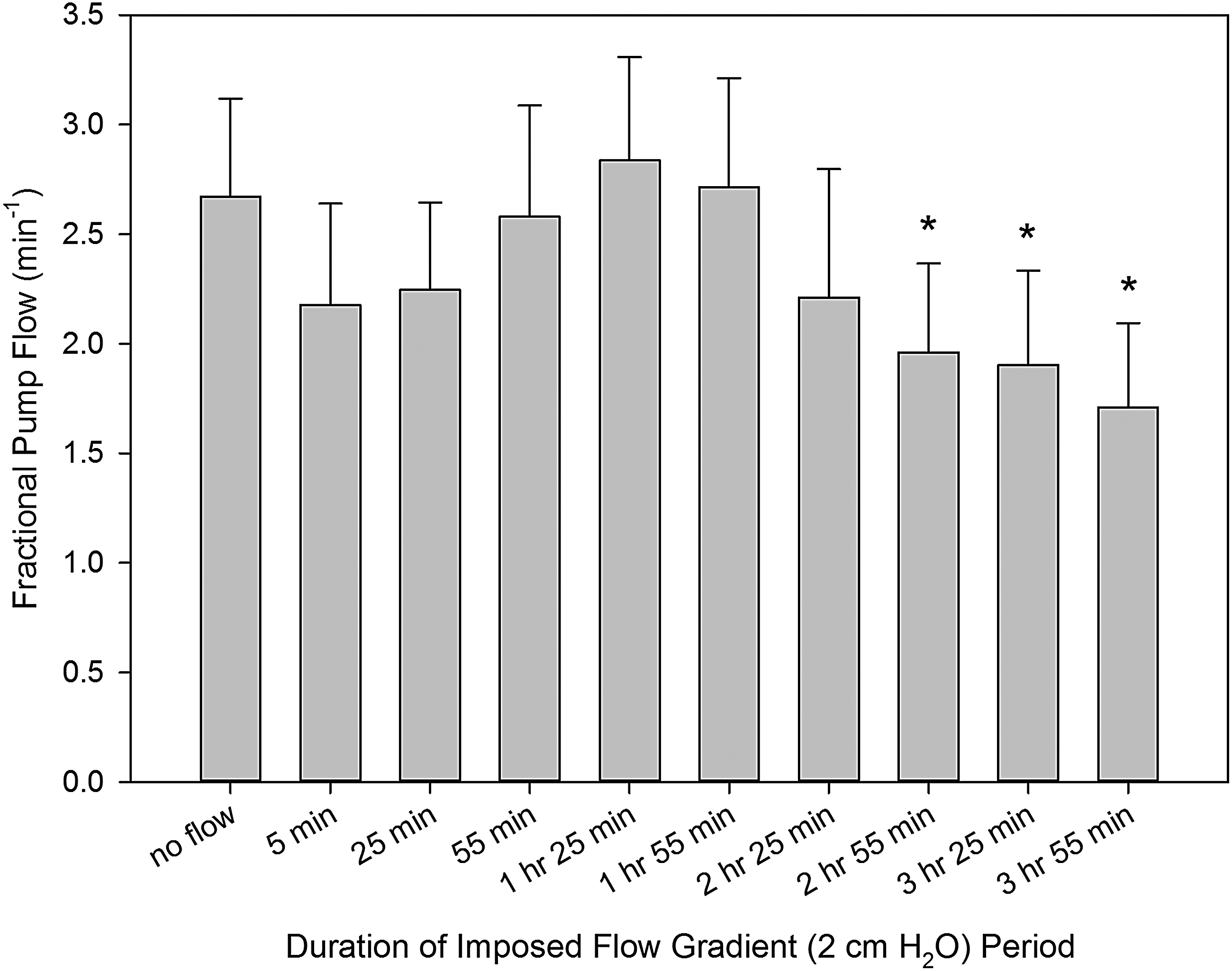
Changes in fractional pump flow of the rat thoracic duct during imposed flow applied for 4 hours. *Indicates significant differences (p values ranging from <0.0001 to 0.0064) between fractional pump flow at control conditions (no imposed flow) and at corresponding time points after creation of an imposed flow.
Table 1 demonstrates the changes in all parameters of the thoracic duct contractility over the duration of the 4-hour period of imposed flow compared with no-flow conditions.
Time represents the duration of imposed flow. p-Values for each parameter demonstrate significant differences between its value at control conditions (no imposed flow) and at corresponding time points after creation of an imposed flow.
AMP, contractile amplitude (%); FPF, fractional pump flow (/min); Freq., contraction frequency (/min); LTI, lymphatic tone index (%).
Discussion
In this article, we for the first time characterized the contractile response of segments of the thoracic duct to imposed flow (2 cm H2O) for a comparatively long period (4 hours). We found that the steady imposed flow gradient induced relaxation of the thoracic duct together with development of the negative chronotropy (decrease of the contraction frequency). Both these effects of imposed flow were strengthened over time.
Contrary to unidirectional decreases in lymphatic tone and contraction frequency, changes in contraction amplitude in response to the prolonged imposed flow demonstrated a more complicated pattern. After a fast but slight initial drop, contraction amplitude increased during the first hour of the presence of flow. Later during the fourth hour of the imposed flow, the contraction amplitude demonstrated a trend to decrease. As a result, during the first 3 hours of the imposed flow, thoracic ducts segments were able to compensate the decrease in contraction frequency by the development of stronger contractions, therefore keeping their active lymph pumping (FPF) at essentially the same level. Only after 3 hours of imposed flow, we observed the decrease of the active lymph pumping of the thoracic duct segments. We considered this as a sign that the positive inotropic compensation of the flow-induced negative chronotropy had reached its limits after 3 hours of the presence of an imposed flow.
While analyzing these patterns of responses of the thoracic duct to prolonged flow, we saw that the very initial response to the imposed flow conditions in the thoracic duct segments, in general, matched those we had observed earlier.5,17 In particular, the NO/cGMP/PKG-mediated regulatory pathway in the thoracic duct is responsible for the reduction of lymphatic tone and for the self-regulatory adjustments of contractility to the initial increase in lymph flow. 17
However, the data we present here on more extended effects of flow on thoracic duct contractility create several potential explanations of the bidirectional nature of the observed changes in contraction amplitude and active lymph pumping of the thoracic duct. Such changes may result from decreases in the sensitivity of lymphatic endothelial cells to permanent wall shear stress over the 4-hour period and/or a decreased reaction of lymphatic muscle to the long-term existence of increased levels of NO. However, the potential reversal of effects of flow due to the action of other non-NO EDRFs on lymphatic muscle due to their local accumulation under the conditions of the prolonged flow can also be considered. For example, it is known that histamine, recently confirmed for its role as EDRF in mesenteric lymphatic vessels,10,11 has a concentration-dependent duality of its effects on lymphatic contractility.30,31
In conclusion, currently, the mechanisms responsible for observed reactions of the rat thoracic duct to the existence of steady imposed flow are not clear. However, the understanding of lymphatic function in different lymphatic networks still suffers from the absence of detailed physiological information obtained in predominantly discovery-driven studies. Therefore, we believe that our new findings are important for a better understanding of the temporal dynamics of how lymphatic vessels behave during the development of prolonged periods of steady flow, such as that which may occur during the initial phases of development of an acute tissue edema. We believe that the fundamental physiological information described in this article creates a solid basis for future hypothesis-driven research on the regulatory mechanisms of lymphatic function during prolonged increases in lymph flow.
Footnotes
Acknowledgment
This work was supported by the National Institutes of Health (NIH RO1 HL-070308, AG030578, HL094269, DK099221, and HL123420).
Author Disclosure Statement
No competing financial interests exist.