
Abstract
Abstract
Background:
Lymphatic filariasis (LF) is a global health problem, with a peculiar nature of parasite-specific immunosuppression that promotes long-term pathology and disability. Immune modulation in the host by parasitic antigens is an integral part of this disease. The current study attempts to dissect the immune responses of aminoacyl-tRNA synthetases (AARS) with emphasis on Brugia malayi asparaginyl-tRNA synthetase (BmAsnRS), since it is one among the highly expressed excretory/secretory proteins expressed in all stages of the parasite life cycle, whereas its role in filarial pathology has not been elaborately studied.
Methods and Results:
In this study, recombinant BmAsnRS (rBmAsnRS) immunological effects were studied in semipermissive filarial animal model Balb/c mice and on clinically defined human samples for LF. In mice study, humoral responses showed considerable titer levels with IgG2a isotype followed by IgG2b and IgG1. Immunoreactivity studies with clinical samples showed significant humoral responses especially in endemic normal with marked levels of IgG1 and IgG2 followed by IgG3. The cell-mediated immune response, evaluated by splenocytes and peripheral blood mononuclear cells proliferation, did not yield significant difference when compared with control groups. Cytokine profiling and qRT-PCR analysis of mice samples immunized with rBmAsnRS showed elevated levels of IFN-γ, IL-10, inhibitory factor—cytotoxic T lymphocyte-associated protein-A (CTLA-4) and Treg cell marker—Forkhead Box P3 (FoxP3).
Conclusions:
These observations suggest that rBmAsnRS has immunomodulatory effects with modified Th2 response along with suppressed cellular proliferation indicating the essence of this molecule for immune evasion by the parasite.
Introduction
Lymphatic Filariasis (LF), also known as elephantiasis, is a mosquito-borne debilitating parasitic disease that afflicts more than 200 million people worldwide. 1 It is caused by filarial nematodes such as Wuchereria bancrofti, Brugia malayi, and Brugia timori. To chronically dwell and breed in human host, filarial nematodes formulated a range of strategies so that it can evade and downregulate the host's immune system. 2 One among the key features of filarial infection is the significant increase in the number of downregulatory cytokines such as IL-10 and TGF-β3,4, which induce regulatory T cells and alternatively activated macrophages, that in turn modulate both Th1 and Th2 responses toward lymphatic pathology. 5
Previous studies conducted using filarial antigens have indicated that these antigens selectively induce naive human T cell differentiation in vitro away from a Th1 phenotype and serve as a model to understand the immune responses during the early exposure of filarial antigens and their subsequent activities in eliciting immune response. 6 This study attempts to explore the host immune responses against filarial asparaginyl-tRNA synthetase (AsnRS), a member of aminoacyl-tRNA synthetases (AARS). AARS are heterogeneous enzymes that play a fundamental role in protein synthesis, protein translation mechanism, and also have other secondary functions. These enzymes are found to be essential for viability in both eukaryotes and prokaryotes.7,8 In helminthic parasites, the critical role of AARS in protein synthesis has been realized and acknowledged by WHO as rational targets for new antiparasitic drugs.7,9
Brugian AsnRS (BmAsnRS) is one of the class IIb AARS enzymes that possesses a distinct N-terminal-beta-barrel domain (OB fold) that binds the tRNA anticodon stem loop. 10 It is a highly expressed excretory/secretory protein (10 folds higher than any other filarial tRNA synthetase) in both the genders of the nematode at all the stages in its life cycle.11,12 It has been reported that BmASnRS has the ability to activate interleukin 8 (IL-8) receptors and can display a selective chemokine-like activity toward the human cells that may play a role in the massive inflammatory response associated with LF.13,14 The recent findings of our research group have demonstrated the significant role of rBmAsnRS in angiogenesis process of LF. 15
To study the complex immune regulatory network involved in filarial pathology, the characterization of filarial antigens becomes obligatory. At the same time, the uninfected individuals (endemic normal [EN]) residing in endemic regions exhibit acquired protective immunity (resistance against filarial parasites), which shows a balanced immune Th-dichotomy that has paved the way to identify novel protective antigens for LF. 16
In the current study, Brugian AsnRS was expressed as recombinant protein to investigate its role in provoking the humoral and cellular immune response in experimental Balb/c mice filarial model using alum as an adjuvant. Furthermore, the immunogenic properties of rBmAsnRS were examined on sera and peripheral blood mononuclear cells (PBMCs) from endemic population.
Materials and Methods
Recombinant BmAsnRS expression and purification
The BmAsnRS full-length gene, cloned in pET15B, was transformed into Escherichia coli BL21 (DE3) cells and the expression of recombinant protein was induced with final concentration of 0.5 mM IPTG. Soluble rBmAsnRS protein, with hexa-histidine tag at the N-terminal, was purified by Immobilized Metal-Affinity Chromatography (IMAC) on chelating Sepharose Fast Flow matrix (GE Healthcare Life Sciences). The expression pattern and the purity were analyzed and visualized on 12% sodium dodecyl sulfate polyacrylamide gels (SDS-PAGE) and western blot analysis. Correct folding, dimerization, and enzymatic activity of rBmAsnRS were confirmed with the help of a published and pretransfer editing assay by malachite green reagent. 17
Animals and immunization
The use of animals in the experimental protocols for this study was approved by the Institutional Animal Ethical Committee (IAEC), Anna University under the guidelines of the Committee for the Purpose of Control and Supervision of Experiments on Animals (CPCSEA), Chennai, India. 6–8 weeks old female Balb/c (H-2d) mice were purchased from Tamil Nadu Veterinary and Animal Sciences University, Chennai and housed in the animal house facility at Center for Biotechnology, Anna University, Chennai.
Before immunization, the animals were bled to obtain preimmune serum. A group of five mice were immunized intraperitoneally with 15 μg of rBmAsnRS in 100 μL of PBS mixed with alum (Thermo Fisher Rockford) at 1:1 ratio. The control group was immunized with alum adjuvant only. Each group was vaccinated at day 0, and four booster doses of the antigen were administered in a periodic manner on day 14, 21, 28, and 35. Mice were tail bled to determine the humoral response elicited by rBmAsnRS. The sera from both the groups, at different immunization times, were stored at −20°C.
Analysis of antibody titer levels
The 96-well microtiter plates (NUNC, Germany) were coated with 100 ng/well of purified rBmAsnRS in 0.06 M carbonate buffer of pH 9.6 and were incubated at 4°C overnight. The plates were washed thrice in PBS containing 0.05% Tween-20 (PBS/T) blocked with 100 μL of 5% skimmed milk and incubated at 37°C for 1 hour. After further washes with PBS/T, 100 μL of sera collected at diverge intervals from the immunized mice was added serially in twofold dilution (1:1000–1:64,000) and incubated for 1 hour at 37°C. The plates were washed again and incubated with goat anti-mouse IgG-ALP conjugate (Santa Cruz biotechnology) (1:1000) at 37°C for 1 hour. Then, p-nitrophenyl phosphate substrate (pNPP) (Amresco, OH) (1 mg/mL) in substrate buffer (100 mM Tris-Cl, pH 9.5, 100 mM NaCl, and 5 mM MgCl2) was added after the final wash. The developed color was quantitatively measured as absorbance at 405 nm using a micro plate ELISA reader. Antibody titers were assessed, and the mean absorbance (±2SD) of alum control sera was taken as the cutoff value.
Estimation of mice IgG isotypes
Mice isotype ELISA was performed using mice sera with maximum IgG titer. In brief, rBmAsnRS antigen was coated (100 ng) in ELISA plate and incubated overnight. After washing, sera samples (1:100 dilution) from different immunized groups were added and incubated at 37°C for 1 hour. Then, goat anti-mouse isotypes such as IgG1, IgG2a, IgG2b, and IgG3-specific antibodies ALP-conjugated were used as per manufacturer's instruction (Sigma-Aldrich, St. Louis, MO) and incubated at 37°C for 1 hour. After incubation and washing, pNPP substrate was sequentially added to the respective wells of isotype antibodies and the absorbance was read at 405 nm.
Splenocyte proliferation assay
To evaluate the possible induction of T cells in Balb/c mice, splenocyte proliferation assay was carried out. The rBmAsnRS protein-immunized mice were sacrificed on day 42 and the spleens were removed aseptically. The splenocytes were separated and washed twice with fresh culture medium (RPMI 1640) (HiMedia Laboratories, Mumbai, India). Lysis buffer (0.8% ammonium chloride) was added to the pellet to remove RBCs. Later, the cells were counted by Trypan blue dye exclusion method and the cells that showed 95% viability were chosen for performing the assay. The single-cell suspension was cultured in triplicate in 96-well plates at 0.2 × 10 6 cells/mL in RPMI 1640 medium (200 μL/well) supplemented with gentamycin (80 μg/mL), 25 mM HEPES, 2 mM glutamine, and 10% fetal bovine serum (Gibco Life Technologies). The cells were then stimulated in vitro with rBmAsnRS (0.1, 0.5, 1, and 2 μg/mL), or positive control Concanavalin A (Con A) (Sigma-Aldrich) (0.1 μg/mL) and incubated at 37°C at 5% CO2 for 72 hours. Wells that contain only the medium were used as unstimulated controls. The proliferation was measured using Alamar Blue (Invitrogen, Paisley, United Kingdom), which is a colorimetric method for determining the number of viable cells (20 μL/well) at 570 nm absorbance subtracted from absorbance of (unstimulated) control. The stimulation index (SI) was calculated as the optical density (OD) of cells stimulated with antigen divided by the OD of unstimulated cells, whereas the results were expressed in the form of mean, SI ± SD.
Cytokine assay
Splenocytes were plated in 24-well flat bottomed culture plates at 4 × 10 6 cells/mL, stimulated with 0.5 μg/mL of rBmAsnRS and incubated for 72 hours. The culture supernatants were removed and filtered through 0.22 μm pores and assayed for cytokine levels using sandwich ELISA for anti-mouse IL-17, IL-4, IL-5, IL-10, and IFN-γ according to the manufacturer's instructions (Ray Bio, Norcross). All the concentrations were derived from standard curves, and the data were expressed in pg/mL after deducting the values of unstimulated cultures through three independent experiments ±SD.
Analysis of cytotoxic T lymphocyte-associated protein-A and FoxP3 by quantitative real time-PCR
TRIzol (Sigma-Aldrich) was added to the splenocytes (4 × 10 6 cells/mL), which were harvested after the removal of supernatant and stored at −80°C. Then the cells were lysed (QIAshredder; Qiagen, Spoorstraat, Venlo, Netherland), after which the total RNA was extracted according to the manufacturer's protocol (RNeasy minikit; Qiagen, Spoorstraat, Venlo, Netherland) and the RNA was dissolved in 20 μL RNase-free water. Reverse transcription was performed at 42°C for 60 minutes using Revert Aid First-Strand cDNA Synthesis Kit (Thermo Fisher Scientific, Pittsburgh, PA). Primer-BLAST software was used to design the primers, and the sequences of mouse-specific primers used were as follows: glyceraldehyde 3-phosphate dehydrogenase (GAPDH) forward primer GGGTCCCAGCTTAGGTTCAT and reverse primer GTTCACACCGACCTTCACCA, cytotoxic T lymphocyte-associated protein-A (CTLA-4) forward primer AGAACCATGCCCGGATTCTG and reverse primer GCTCTGTTGGGGGCATTTTC, and FoxP3 forward primer GCTAGAACAGGTGCCTCAGC and reverse GAGGAGTTCAGTAGAGGGGC (Sigma-Aldrich). Real-time quantitative PCR was performed in Applied Biosystems Step One Real-Time PCR instrument using KAPA SYBR® FAST qPCR Master Mix Kit (KAPA bio systems, MA). The relative transcripts were determined according to the manufacturer's protocol and data were analyzed for a fold change in expression using the formula 2−ΔΔCT.
Human sera samples
The samples were collected in accordance to the U.S. Department of Health and Human Services Human Experimentation Guidelines. All the procedures followed were in accordance with the guidelines issued by the Department of Public Health, Government of Tamil Nadu, India for the studies that deal with human subjects. The protocols adopted were also approved by the Institutional Review Board at the Centre for Biotechnology, Anna University, Chennai, Tamil Nadu, India. Clinical groups were classified into microfilariae (MF)-positive patients and chronic pathology (CP) patients or EN individuals, and the blood samples were collected after obtaining informed consents. The samples were categorized as MF based on the detection of circulating MF through microscopy and/or presence of circulating antigen using ELISA. 18 Similarly, the subjects showing lymphedema and other visible clinical symptoms of filariasis were grouped under CP.19,20 Preimmune or nonendemic normal (NEN) control samples were obtained from Bint Al-huda Teaching Hospital, Thi-Qar, Iraq.
Screening of human sera for BmAsnRS antibodies and IgG isotypes
Sera from four different clinical groups such as EN, MF, CP (n = 20), and NEN (n = 10) were screened for anti-BmAsnRS IgG antibodies using rBmAsnRS as antigen in indirect ELISA. The microtiter plates were coated with 100 ng/well of purified rBmAsnRS and incubated overnight at 4°C. After washing and blocking, the plates were incubated with 1:100 dilution of human sera at 37°C for 1 hour. It was further washed and incubated with goat anti-human IgG-ALP conjugate (1: 1000) (Sigma-Aldrich) at 37°C for 1 hour. After further washes, the plates were developed with pNPP substrate (Amresco) (1 mg/mL) in substrate buffer, and absorbance was measured at 405 nm.
For estimating IgG isotypes of anti-BmAsnRS, the human sera samples (n = 10) (1:100 dilution) from various study groups were analyzed by indirect ELISA using mouse monoclonal antihuman IgG1, IgG2, IgG3, and IgG4 (Sigma-Aldrich) and probed with goat anti-mouse ALP conjugate (Santa Cruz) as a secondary antibody. Then, pNPP substrate was added to detect the levels of IgG isotypes in the sera.
PBMCs proliferation assay
PBMCs were obtained from heparinized blood, purified by sedimentation on a Ficoll gradient (Sigma-Aldrich) from EN, CP, and NEN (n = 10). The cells were washed and their viability was determined by Trypan Blue dye exclusion method and those cells that showed 95% viability were utilized for performing the assay. PBMCs were placed in flat-bottomed wells of micro titer plates at a concentration of 0.2 × 10 6 cells/mL in RPMI-1640 supplemented with gentamicin (80 μg/mL), 25 mM HEPES, 2 mM glutamine, and 10% fetal bovine serum. The cells were stimulated with rBmAsnRS protein (0.1, 0.5, 1, and 2 μg/well) or with Con A (0.1 μg/well) as a positive control incubated at 37°C at 5% CO2 for 72 hours. Wells loaded with only medium were used as unstimulated controls. The proliferation was measured by Alamar Blue. All cultures were prepared in triplicates and the results were expressed as mean absorbance at 570 nm subtracted from absorbance of (unstimulated) control. The proliferative responses were calculated as mentioned earlier.
Statistical analyses
All statistical analyses were performed using GraphPad Prism software version 5 (GraphPad Software, Inc., CA). The difference between two means was compared using nonparametrical analysis of Student's t-test. For multiple comparisons, nonparametric Kruskal–Wallis test and Dunn's posttest were used. A probability (p) value ≤0.05 was considered statistically significant. Each experiment was repeated three times independently.
Results
rBmAsnRS purification
The recombinant BmAsnRS was expressed in E. coli host as soluble fraction and purified by histidine tag present in the plasmid construct. The purification was performed by IMAC with metal binding ability of histidine tag. The protein fraction, eluting as a major peak, gave an intense band with a molecular weight of 63 kDa in SDS-PAGE, confirming its purity (Fig. 1a). The protein identity was further confirmed by western blot analysis with anti-His antibody (Fig. 1b). Each new lot of protein was assessed for endotoxin using limulus amoebocyte lysate (LAL) assay to rule out the presence of endotoxins above acceptable limits (<0.01 ng/μg of protein). The retention of enzyme activity in the purified protein was confirmed by Malachite green assay before immunization.

Mice antibody titer and isotype profile
The total IgG antibodies in sera were measured by ELISA and the antibodies' production results were expressed in terms of peak titers (Fig. 2). rBmAsnRS elicited potent humoral responses with peak titer of 32,000 in mice on day 21 after immunization. The antigen-specific IgG levels for rBmAsnRS were significantly high (p < 0.001) compared to alum control, from which we can derive that one booster is sufficient for rBmAsnRS to elevate the antibody response in semipermissive animals.
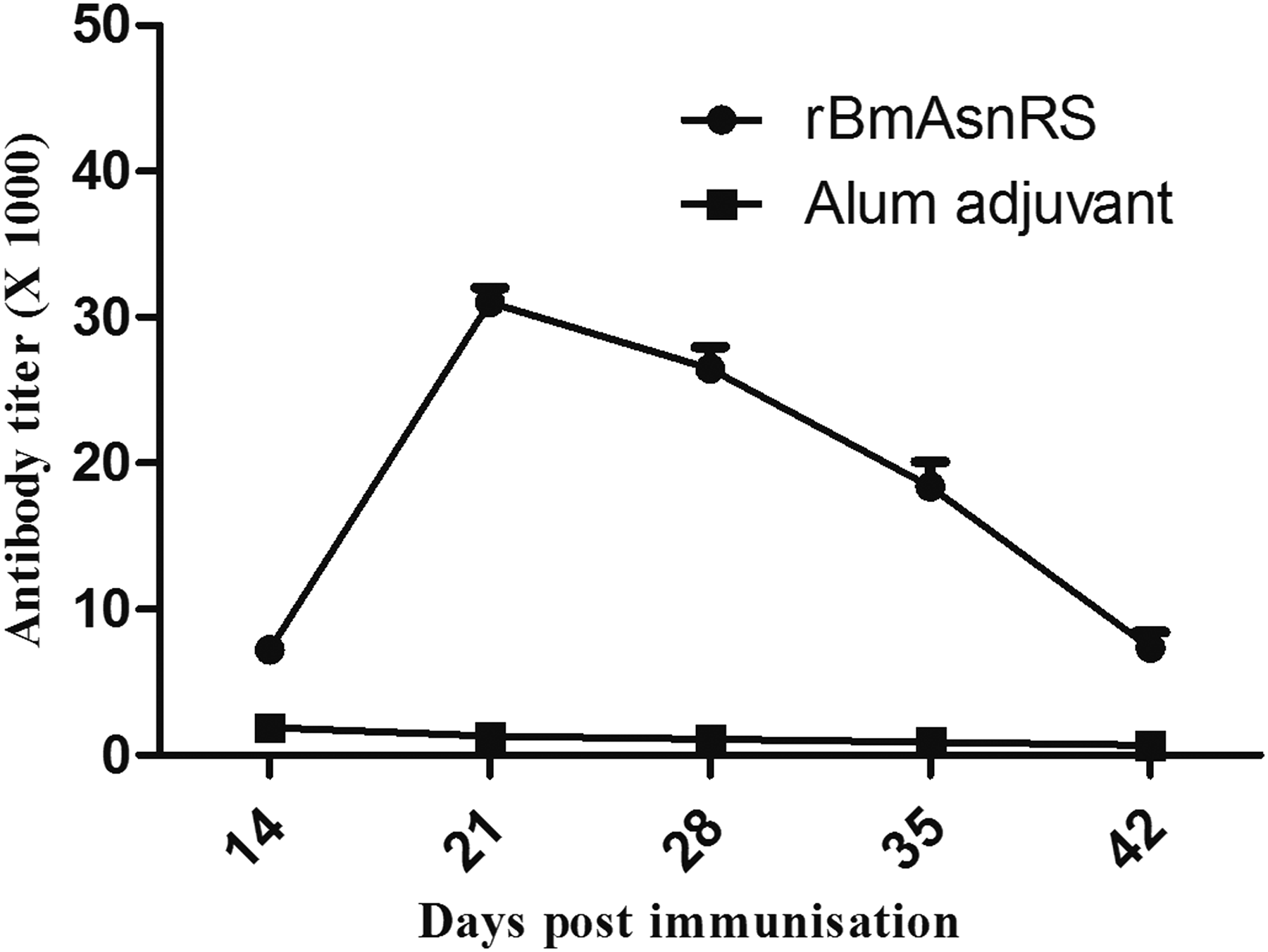
Humoral response by ELISA in mice immunized with rBmAsnRS. Specific antibody titer (in thousands) in mice at different intervals postimmunization. The data represent the mean titer ± SD of five mice in each group.
Mouse anti-rBmAsnRS IgG1, IgG2a, IgG2b, and IgG3 isotype profiles, measured on day 21 postimmunization sera had shown the maximum total titer levels. The profile showed a significant (p < 0.001) elevation in the levels of IgG2a, followed by IgG2b and IgG1 isotypes (Fig. 3).
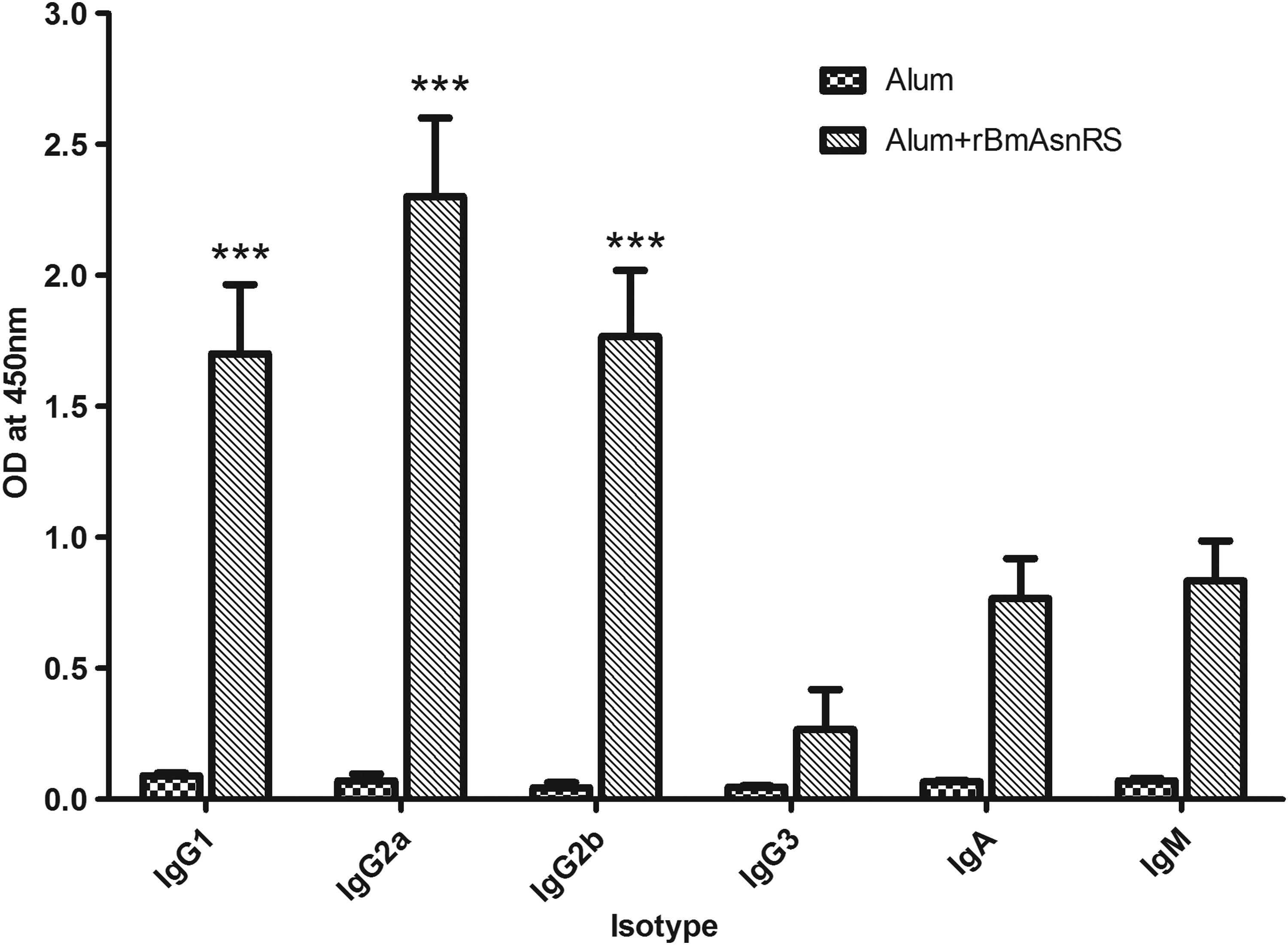
The antibody isotypes in Balb/c mice measured in 1:100 diluted sera by ELISA. Data represent the mean absorbance of antibody isotypes from five mice in each group ± SD given as bars. The asterisks on top of the bars indicate a significantly-high OD value (***represents p < 0.001) compared with other antibody isotypes. The statistical significance was calculated by Kruskal–Wallis test and Dunn's posttest. OD, optical density.
Cellular immune response of rBmAsnRS in mice model
To evaluate the induction of T cells in Balb/c mice, splenocyte proliferation assay was carried out. The splenocytes showed the maximum stimulation index of proliferation when pretreated with 0.5 μg/mL of rBmAsnRS. Hence, this concentration was used for further experiments. When splenocytes of rBmAsnRS immunized mice were stimulated with the same antigen, the stimulation index (mean SI = 11 ± 3) was not significant enough (p = 0.4) compared with control group immunized with alum (SI = 10.33 ± 1.53) (Fig. 4). Under stimulation with Concanavalin A (ConA) (0.1 μg/mL), mitogen induced significant proliferation in both adjuvant control and antigen-immunized mice splenocytes (mean SI = 35.67 ± 2.08 and 36 ± 3.61 respectively), thus confirming the proliferation of T cells.
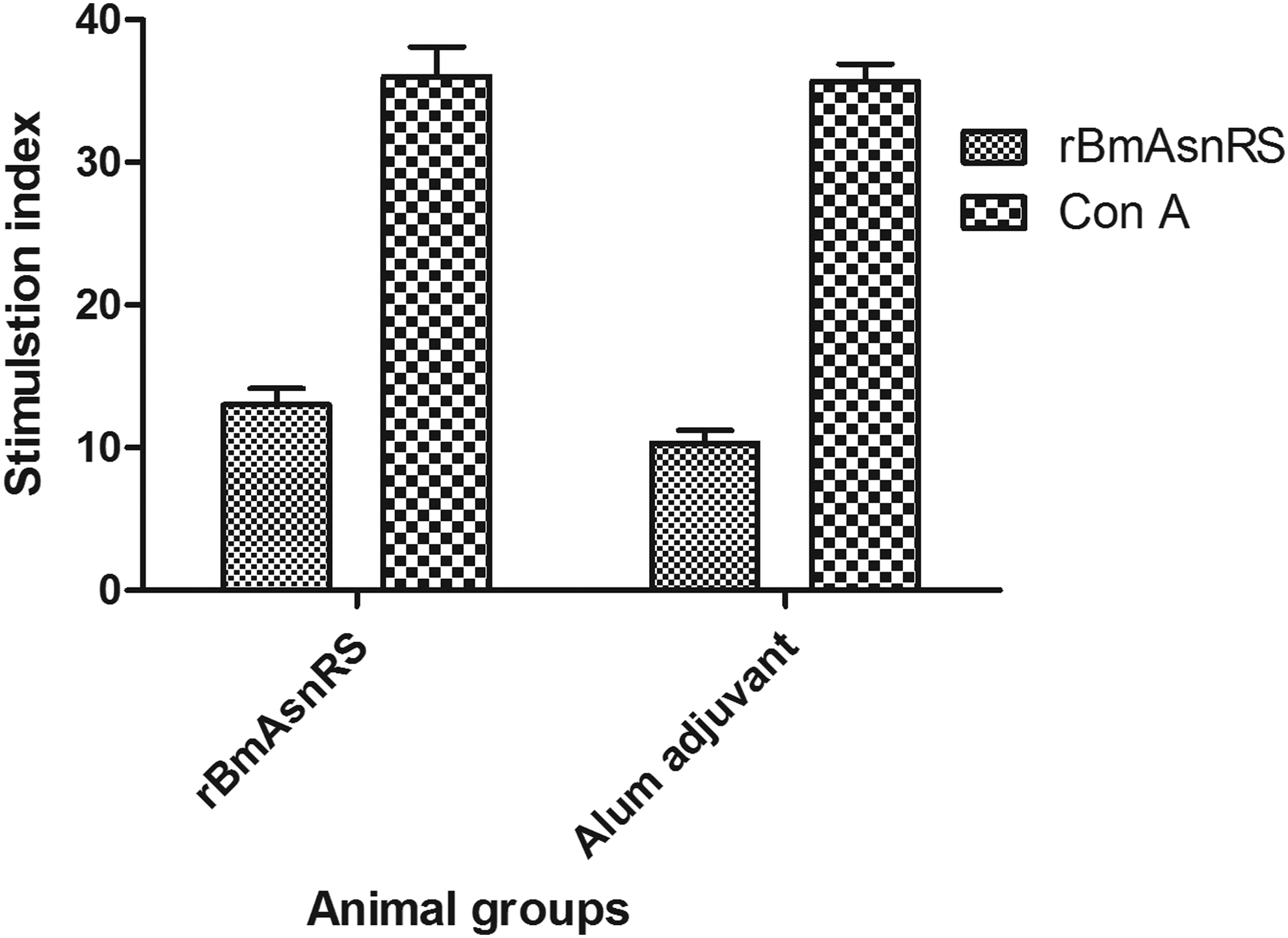
Splenocyte proliferation of alum control and alum+rBmAsnRS immunized mice stimulated with rBmAsnRS and positive control (ConA). Data are represented as mean SI ± SD of splenocyte cells stimulated with 0.5 μg/well of rBmAsnRS and 0.1 μg/well of ConA from five mice in each. SI, stimulation index.
Filarial AsnRS antigen showed a significant elevation in the levels of IFN-γ (p < 0.001) followed by IL-10 (p < 0.05), whereas no significant difference was found in the levels of IL-4, IL-5, and IL-17 (Table 1).
Cytokine Levels (pg/ml) in Culture Supernatants of Spleen Cells from Immunized Mice Stimulated In Vitro with rBmAsnRS Antigen at 0.5 μg/ml
All concentrations were derived from standard curves and data are expressed after deducting the values of unstimulated cultures. Experiments were done in triplicate and represented as mean concentration ± SD.
rBmAsnRS, recombinant Brugia malayi asparaginyl-tRNA synthetase.
Quantitative real time-PCR analysis of FoxP3 and CTLA-4
As the humoral and cellular response data in this study showed elevated levels of Treg antibodies (IgG2b), cytokine IL-10, and rBmAsnRS antigen-specific suppressed T cell proliferation, the expression levels of Treg cell transcription factor (FoxP3) and marker CTLA-4 were analyzed. Splenocytes from both alum control and vaccinated with alum+rBmAsnRS (five mice in each group) were stimulated in triplicates with 0.5 μg/mL of rBmAsnRS. The expression levels of FoxP3 and CTLA-4 were normalized against endogenous gene (GAPDH) as control. Both FoxP3 and CTLA-4 transcription were significantly (p < 0.001) stimulated in splenocytes that were already primed with rBmAsnRS antigen (Fig. 5). FoxP3 was elevated to the levels of 5.324 ± 0.972 (mean ± SD) compared with 1.269 ± 0.179 in alum control. In case of CTLA-4 mRNA levels, 4.324 ± 0.646-fold increase was found relative to 1.758 ± 0.186 in adjuvant control.
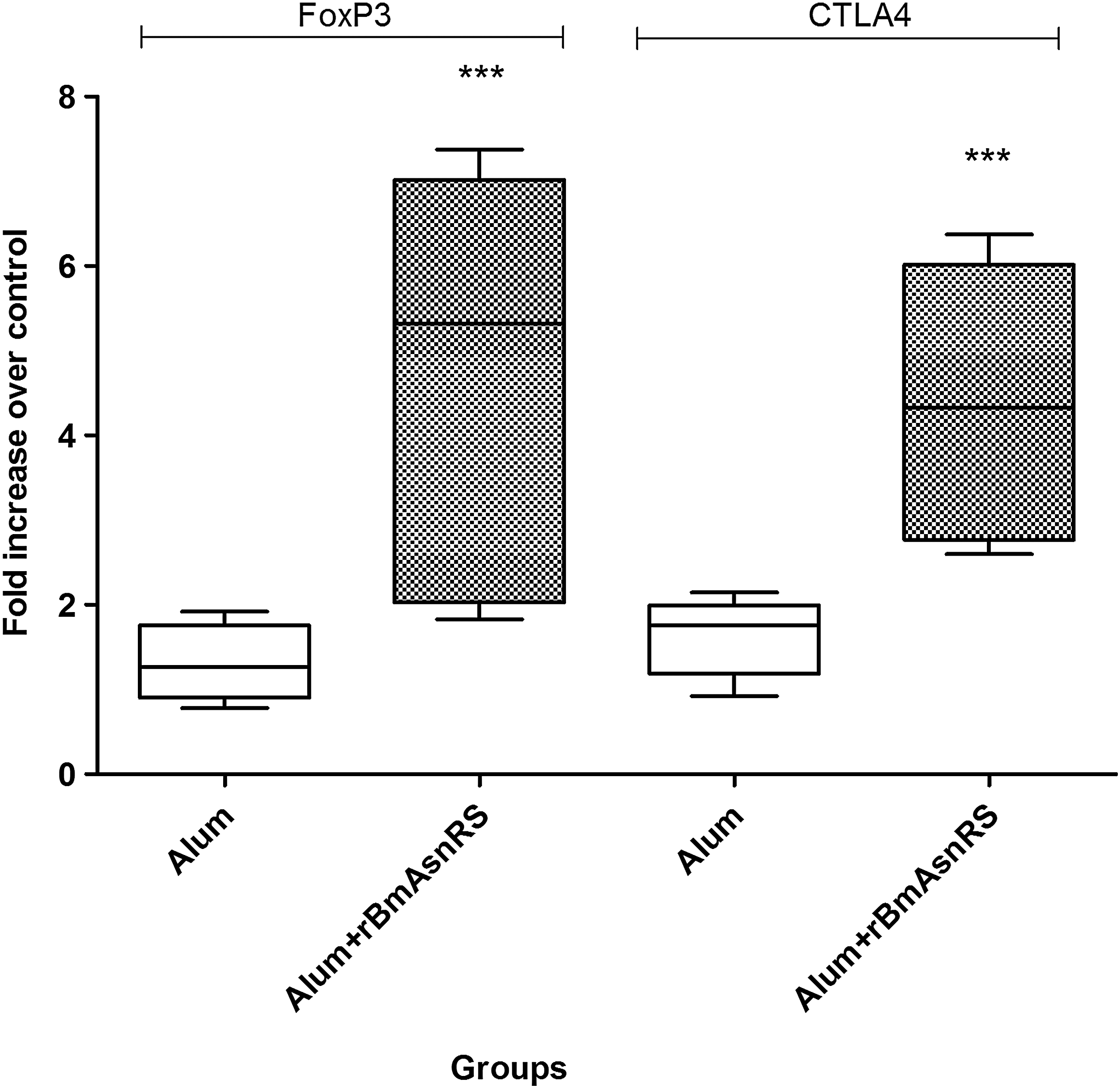
Expression profile of FoxP3 and CTLA-4 genes when stimulated with rBmAsnRS in mice model. Data generated from alum control and alum+rBmAsnRS mice groups (n = 5) were articulated in box and whisker plot. In the plot, horizontal lines represent 25th, 50th, and 75th percentiles and vertical lines represent maximum and minimum value. The asterisks on top indicate a significantly high fold increase (***represents p < 0.001) compared with other antigens within the animal groups. CTLA-4, cytotoxic T lymphocyte-associated protein-A.
Serological responses to rBmAsnRS from different clinical groups of LF
With regard to immunoglobulin responses in NEN sera, subjects living in a filariasis endemic region of southern India showed significant reactivity to rBmAsnRS. The EN group had the highest level of reactivity (p < 0.001) compared with MF and CP individuals (Fig. 6). The mean absorbance of the negative control NEN sera mean ± 3SD was taken as the cutoff value. EN subjects had the highest levels of anti-BmAsnRS antibodies (i.e., mean OD = 1.205 ± 0.288) in their sera. There was no significant difference found between MF (mean OD = 0.524 ± 0.138) and CP (mean OD = 0.598 ± 0.144) groups. Earlier reports with filarial antigens such as ALT-2, GST, TGA, VAH, and TRX showed significantly high recognition of naturally immune sera (EN), whereas the MF and CP had showed higher protection efficacy.

Reactivity of rBmAsnRS with human sera samples from different clinical groups. EN, MF, CP (n = 20 for each group), and NEN sera (n = 10) were used as negative control. The data are represented as scatter plot where each dot represents absorbance of individual sera. The line drawn parallel to the X-axis represents the cutoff value (mean OD of NEN ±3 SD). CP, chronic pathology; EN, endemic normal; MF, microfilariae; NEN, nonendemic normal.
Clinical isotype profile
The BmAsnRS antibody IgG isotype profiles in EN, MF, CP, and NEN (n = 10) were examined. The analysis showed that the anti-BmAsnRS IgG antibodies in the sera of EN and CP individuals were occupied by IgG1 isotype (p < 0.001), followed by IgG2 and IgG3, while IgG4 isotype was least detectable, whereas the IgG response from MF individuals showed significantly (p < 0.005) high levels of IgG2 isotype (Fig. 7).
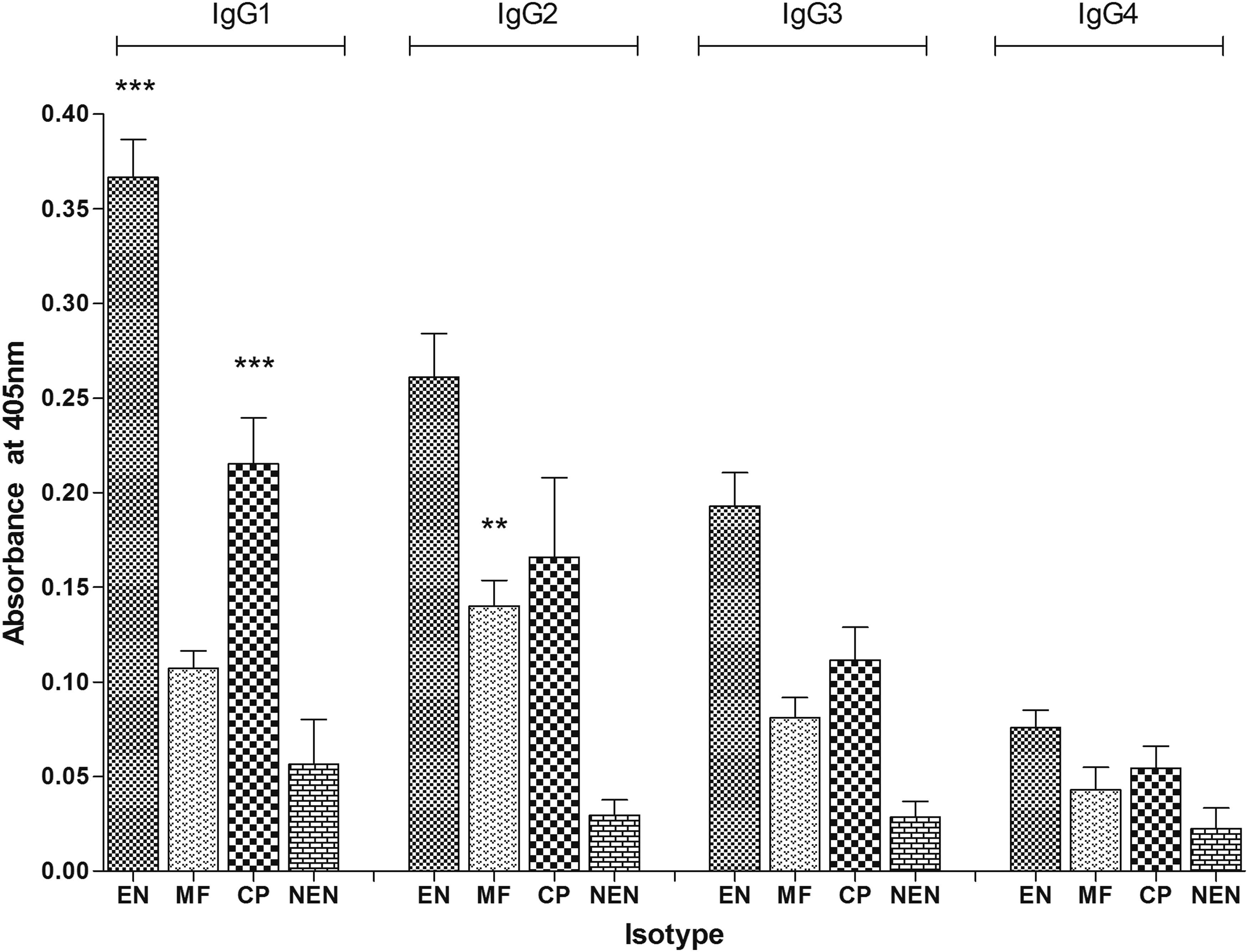
BmAsnRS antibody isotype profiles in EN, MF, CP, and NEN evaluated using an isotype-specific ELISA. Results showed significant levels of IgG1 (***represents p < 0.001) in EN and CP groups, IgG2 was highly significant (**represents p < 0.005) in MF than the others. Data represent mean absorbance of 10 samples mean ± SD in each group.
PBMCs proliferation assay
The lymphoproliferative response of PBMCs separated from EN, CP, and NEN subjects (n = 10) was examined in vitro with rBmAsnRS protein and concanavalin A (ConA). The putatively immune PBMC samples (EN) showed a stimulation of mean (SI = 3.007 ± 0.38) and diseased individuals (CP) with elicited stimulation of mean (SI = 2.576 ± 0.543) for rBmAsnRS, which was not significantly different between the two endemic regional samples. Furthermore, there was no significance in stimulation between endemic region samples and NEN samples (mean SI = 2.204 ± 0.377) with the antigen. In all the three groups, PBMCs had illustrated good stimulation index for the positive control (ConA) (Fig. 8). These data indicate the hyporesponsiveness state of BmAsnRS-specific T cell response in endemic population.
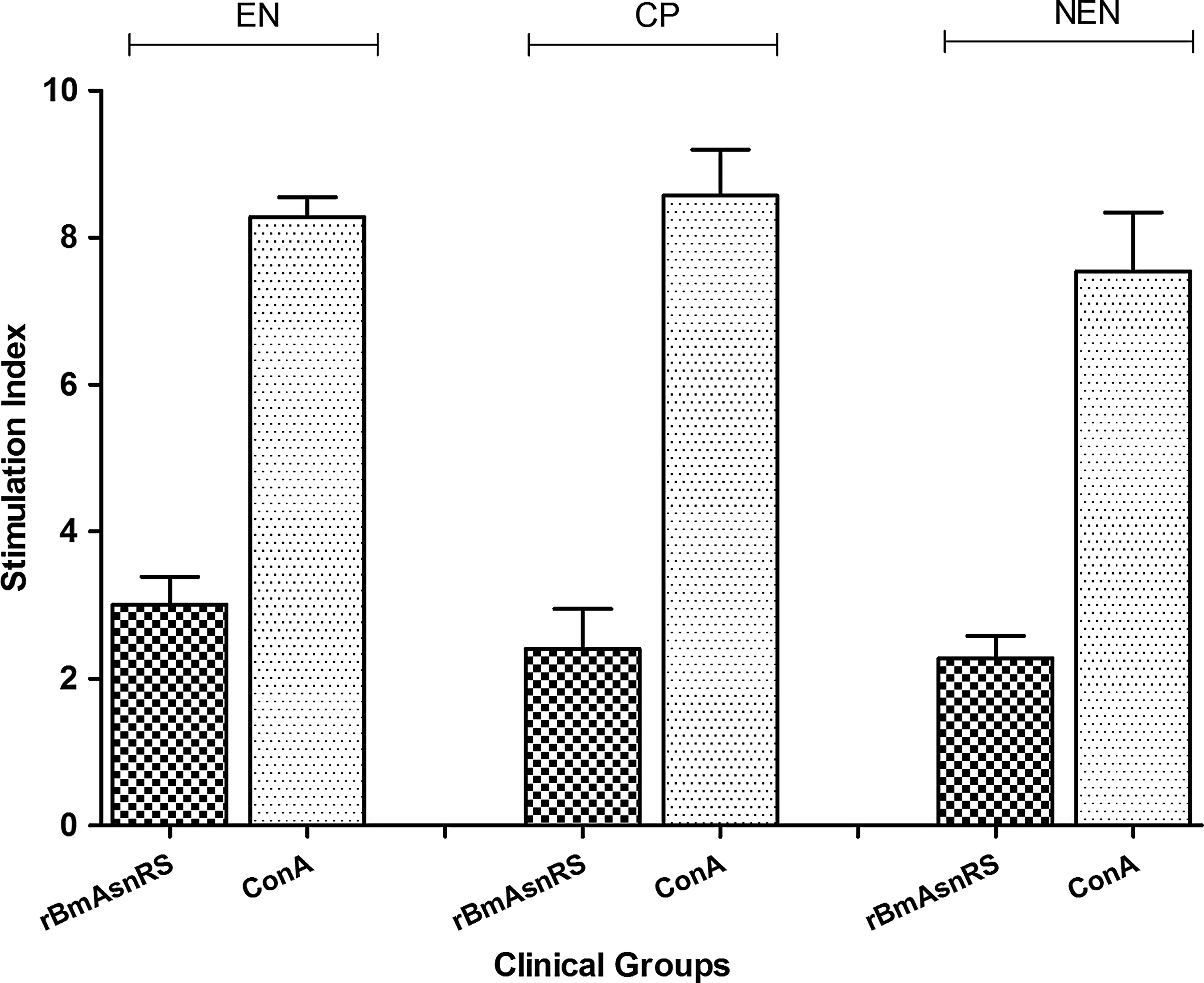
Proliferation of human PBMCs stimulated with 0.5 μg rBmAsnRS. ConA was used as positive control. The data are represented as mean SI ± SD of EN, CP and NEN individuals (n = 10). The data indicate nonsignificant proliferation in all groups for the antigen. PBMCs, peripheral blood mononuclear cells.
Discussion
Immune evasion of filarial parasites exerts profound immunoregulatory effects on the host immune system with both parasite antigen-specific responses and more generalized levels of immune modulation. 20 These features suggest that LF is the most immunologically complex infections of human beings, despite the existence of protective immunity in humans. 21 The antigenicity and immunological role of AARS enzymes were not well explored in most species except humans, where their role in association with human diseases had been explored in the context of noninfectious diseases. 22 The recent studies from our laboratory on implications of Brugian AsnRS in triggering angiogenesis prompted us to investigate the immune response elicited by it. Hence, this work was carried out to define the immunological role of filarial-derived AARS enzyme, BmAsnRS as a whole protein with alum as an adjuvant in mice model, and humoral and cellular response in human samples.
Immunization with rBmAsnRS-elicited high titers in Balb/c mice suggested the presence of epitopes to stimulate B-cells and immunogenicity. Higher antibody titers could be directly correlated with protective efficacy of vaccine candidates where antibodies play a pivotal role in the clearance of L3 and microfilaria. 23 Although many filarial antigens can show reactivity, when antisera is raised in mice, not all the antigens were recognized in natural infection in the specific host. 24 Analysis of IgG isotype profile in mice showed elevated levels of IgG2a and IgG2b followed by IgG1 against rBmAsnRS, which indicate the propensity toward both, Th1 and Th2 response. The recent studies to evaluate immune responses against Leishmania tropica infection in Balb/c mice have also shown the correlation of IgG2a and IgG1 in association with protective responses. 25
In the context of cellular response in mice, the stimulation index results suggest no significant proliferation response to rBmAsnRS in splenocytes prestimulated with the antigen compared to adjuvant control (alum) group. These findings imply antigen-specific immune suppression which could be due to low expression of costimulatory signals on antigen-presenting cells, low production of proinflammatory cytokines, and insufficient production of memory cells resulting in impaired T cell proliferation, rendering T cells tolerance.21,26 In the natural course of infection with B. malayi, the parasites are capable of inducing high levels of regulatory cytokines such as IL-10 and TGF-β, causing low proliferative response of T-cells. 27
Cytokines' analysis showed increased production of IFN-γ and IL-10 in culture supernatants stimulated with rBmAsnRS, but no significant change in IL-5, IL-4, and IL-17 levels over control. Although there is a strong humoral response, it is not a classical Th2 response, which might be due to elevated immunomodulatory IL-10 levels. A similar finding has been reported by Specht et al., in which an antagonist activity was proved between IL-4 and IL-10 in filariasis infection. 28 Although IL-10 appears to be involved in B cell stimulation, it has inhibitory effect on Th2 response predominantly during allergic studies in animal models.29,30 While, in infections with helminth parasites, which induces Th2 response as in Trichuris muris, it has been found that IL-10 promotes Th2 response. 31 This study suggests that rBmAsnRS-specific IL-10 elevation could have prevented class switching of antibodies induced by IL-4 and IL-5 and could be responsible for the humoral response. Despite less proliferation response to rBmAsnRS, IFN-γ levels were found to be elevated, which is consistent with Taylor et al. study, which established that although there was hyporesponsiveness, production of IFN-γ was high during filarial infection. 32 Elevation of IFN-γ levels cause activation of macrophage and produce nitric oxide that is directly responsible for parasite clearance in many parasitic diseases, including LF.33,34 Thus, rBmAsnRS could lead to a mixed type of immune response skewed toward Th1/Treg with possible modulation of Th2 arm.
To investigate the effects on regulatory T cells by BmAsnRS, we analyzed the expression of critical transcription factor of Treg cells, FoxP3, and Treg marker, CTLA-4, 35 where both showed approximately four- to five-fold augmentation. In this study, we have the evidence that BmAsnRS provokes Treg-mediated response through CTLA-4, and earlier studies proved that blocking CTLA-4 can partially restore the immunological responsiveness in cells from infected individuals and thus mount Th1/Th2 response.36,37 Similarly, there are evidences on major role of FoxP3 high expression levels during filarial infections in suppression of effector T cell and Th2 immunity.24,38 Thus, this could explain the T cell hyporesponsiveness in this study and the expression of CTLA-4 on T cell, which was found to suppress the IL-5 production and is concurrent with our cytokines profile. 37
Naturally immune EN clinical samples showed considerably high reactivity against rBmAsnRS, which correlates with mice humoral response and thus confirmed the presence of immunodominant epitopes in the protein. Isotype profile of EN sera illustrated elevated levels of IgG1, which is known as one of the major immunoglobulins involved in antifilarial immunity. 39 IgG1 and IgG3 are strong activators of complement C1 compared to IgG2, whereas IgG4 does not activate complement. 40 The high levels of IgG1, followed by IgG2 and IgG3, elicited by rBmAsnRS could hypothesize the role of innate immunity in protective immunity through activation of complement cascade and also in antibody-dependent cellular cytotoxicity. There is concurrence to the pattern of isotype profile observed in all categories except MF samples (maximum IgG2), which can be related to the stage of infection. 41
In vitro human PBMC stimulation with rBmAsnRS showed no significant proliferative difference between clinically diverse groups (EN, CP, and NEN). The diminished levels of proliferation can be attributed to the immunomodulatory regions of BmAsnRS that could reduce the antigen-specific proliferation and might have also contributed to elevated IL-10 levels, thereby dampening the T cell-mediated response. Many studies have established that lymphocyte hyporesponsiveness, correlated with crude filarial antigens, might be due to immunomodulatory regions involved in whole proteins.23,42 rBmAsnRS, by itself, elucidated suppressive T cell response, which is not uncommon for filarial antigens that could result in an overall parasite-specific immunosuppressive environment to establish the parasitic helminthes infections in human hosts. 21
To conclude, our analysis of mice cytokine profile signified the immunosuppressive role of rBmAsnRS with altered splenocyte cell proliferation. rBmAsnRS induced a modified Th2 response with shut down of IL-5 along with reinforcement of IL-10 switching toward regulatory cells phenotype with upregulation in FoxP3 and CTLA-4 expression. Our results from animal model and human clinical samples demonstrated that immune response against rBmAsnRS is predominantly immunomodulatory indicating the importance of this molecule for the parasite to evade the host immune system. These findings suggest that BmAsnRS promote Treg-mediated response, which might help in future to find new therapeutic strategies for controlling LF.
Footnotes
Acknowledgments
The research work was conducted with the help of equipment procured under MHRD-CEMA F.NO-5-3/2015-TS VII and BUILDER BT/PR12153/INF/22/200/2014 programs. A.H. acknowledges the fellowship support provided through the ICCR (Indian Council of Cultural Relationships) and the ministry of Iraqi higher education and scientific research.
Authors' Contributions
Conceptualization, supervision, and visualization of the work was done by A.D., the experiments were designed by A.D., M.K., M.N., and A.H., and performed by A.H., M.N., S.J., J.J., K.K., and S.S. Statistical data analysis was performed by A.H. and M.N., and draft was prepared by A.H., M.N., S.S., M.K., and A.D. The resources required to conduct this research work were kindly provided by A.D. and M.K.
Author Disclosure Statement
No competing financial interests exist.