
Abstract
Abstract
Background:
Metastatic tumor cells spread through lymphatic vessels and colonize draining lymph nodes (LNs). It is known that tumors induce lymphangiogenesis to enhance lymphatic metastasis and that metastatic cancer cells are carried by lymph flow to LNs.
Methods and Results:
Here, we investigated the molecular and cellular regulation of collecting lymphatic vessel contraction in vessels draining a metastatic tumor using intravital microscopy. In tumor-draining collecting lymphatic vessels, we found vessel contraction was suppressed. The infiltration of peritumor tissue by inducible nitric oxide synthase positive and CD11b+Gr1+ myeloid cells played a critical role in the suppression of lymphatic contraction. Depletion of Gr1+ cells with an anti-Gr1 antibody improved contraction of tumor-draining lymphatic vessels. In addition, inducing tumor cell death restored lymphatic contraction in nude mice.
Conclusions:
These findings indicate that tumors contribute to regulation of lymphatic transport in a reversible manner, warranting further investigation into the role of impaired lymphatic transport in cancer progression.
Introduction
D
Unlike initial lymphatic vessels, collecting lymphatic vessels are invested in organized lymphatic muscle cells (LMCs) that drive lymphatic contraction and force lymph toward LNs. 6 Lymph transport is driven by both passive forces from tissue movement and contractions from LMCs. 6 However, proper LMC coverage is required to maintain lymphatic function, demonstrating the importance of these cells. 7 Nitric oxide (NO) is a key modulator of LMC activity and, therefore, lymph transport.8,9
Previously, it was demonstrated that tumor cell supernatant reduces lymphatic contraction ex vivo through an NO-mediated mechanism. 10 Recently, noninvasive methods to study lymphatic pumping with near infrared fluorescence imaging showed reduced tumor-draining lymphatic contraction. 11 However, how tumors regulate lymphatic contraction is not clear. In this study, we investigate alterations of tumor-draining collecting lymphatic vessel contraction through intravital imaging to assess the molecular and cellular regulation of this process.
In previous studies, we found that CD11b+Gr1+ cells suppressed lymphatic contraction by expression of inducible nitric oxide synthase (iNOS) during sterile inflammation. 12 CD11b+Gr1+ cells are a heterogeneous population of immature myeloid cells—including precursors of granulocytes, macrophages/monocytes, and dendritic cells—with powerful immune suppressive function in cancer progression.13,14 CD11b+Gr1+ cells also suppress T cell responses through multiple mechanisms, including inhibiting CD3-zeta chain expression as well as producing arginase 1 and NO (through iNOS).15–17 In this study, we investigate whether tumor-associated CD11b+Gr1+ cells impair lymphatic contraction during cancer progression.
Materials and Methods
Mice
In all experiments, 8- to 10-week-old mice were used. Before imaging, mice were anesthetized using a ketamine–xylazine mixture at 100 mg:10 mg per kg body weight. The protocol for the animal experiments was reviewed and approved by Institutional Animal Care and Use Committee of the Massachusetts General Hospital. The procedures were performed in accordance with the approved guidelines.
Statistical analysis
Significance was determined by a t-test with a Welch's correction for comparison between two groups or an analysis of variance with a Tukey's post hoc test for comparison between more than two groups. p-Values <0.05 were considered significant. Data analysis was performed in Prism (GraphPad Software, La Jolla, CA).
Tumor implantation
A total of 2 × 10 6 tumor cells were implanted in the lower leg close to the dorsal side of the footpad. The lymphatic contraction was assessed when paw thickness reaches 6–8 mm. At this stage of B16F10 tumor growth, there was spontaneous LN metastasis in ∼40% of the mice. We used four different tumor cell lines in their syngeneic mouse strains: B16F1 and B16F10 (melanoma in C57BL/6 mice), P0008 (breast cancer in FVB mice), and SCCVII (squamous cell carcinoma in C3H mice) to study alterations of tumor-draining lymphatic vessel contraction.
Imaging collecting lymphatic vessels
In brief, mice were anesthetized and 2 μL of 2% FITC-Dextran (2 × 10 6 kDa) was injected into the footpad, as previously described. 18 The leg skin and underlying connective tissue near the popliteal lymphatic vessel were carefully removed using sterile microsurgical dissection, thereby exposing the collecting lymphatic vessels. Time-lapse images were taken with an inverted fluorescence microscope. Images of the lymphatic vessels were analyzed using in-house Matlab code. Ejection fraction—the percentage of total volume ejected per contraction—was used to quantify the strength of lymphatic contraction. 12
Gr1+ cell depletion
Control immunoglobulin G or anti-Gr1 antibody (clone RB6-8C5) 100 μg/injection was injected 7 days after tumor implantation and every 3 days until the time of imaging.
HSTS26T implantation and diphtheria toxin treatment
HSTS26T human sarcoma tumor chunks (1–2 mm) were implanted on dorsal side of the footpad in nude mice, and when the paw thickness reached 6 mm, mice were randomly divided into two groups [phosphate-buffered saline (PBS) or diphtheria toxin (DT)]. DT (2 μg/mouse) or PBS was injected i.p. once after randomization and again 3 days later. Five days after the first injection, lymphatic contraction was measured.
Imaging LN metastases
Hematoxylin and eosin (H&E) staining was performed on multiple 10 μm frozen sections from individual TDLNs. TDLNs were imaged and examined by two independent reviewers to determine the presence of metastatic tumor cells.
Results
Multiple tumor types suppress tumor-draining lymphatic vessel contraction
Cancer-induced lymphangiogenesis is correlated with increased dissemination of tumor cells to regional LNs and poor prognosis in a variety of human cancers.19–27 Although tumor-draining lymphatic vessels have been shown to have impaired contraction. 11 it is not clear how cancer modulates lymphatic contraction strength. We measured the lymphatic contraction strength when hind paws bearing tumors reached a thickness of 6–8 mm. At this stage, there were only sporadic cancer cells or very small LN metastases in the TDLN. Lymphatic vessels were dilated, and there was a reduction in contraction strength across all tumor types tested (Fig. 1, Table 1, Supplementary Movies 1 and 2. Interestingly, lymphatic ejection fraction and frequency under normal conditions in nontumor-bearing mice are dependent on the mouse strain (Table 1).
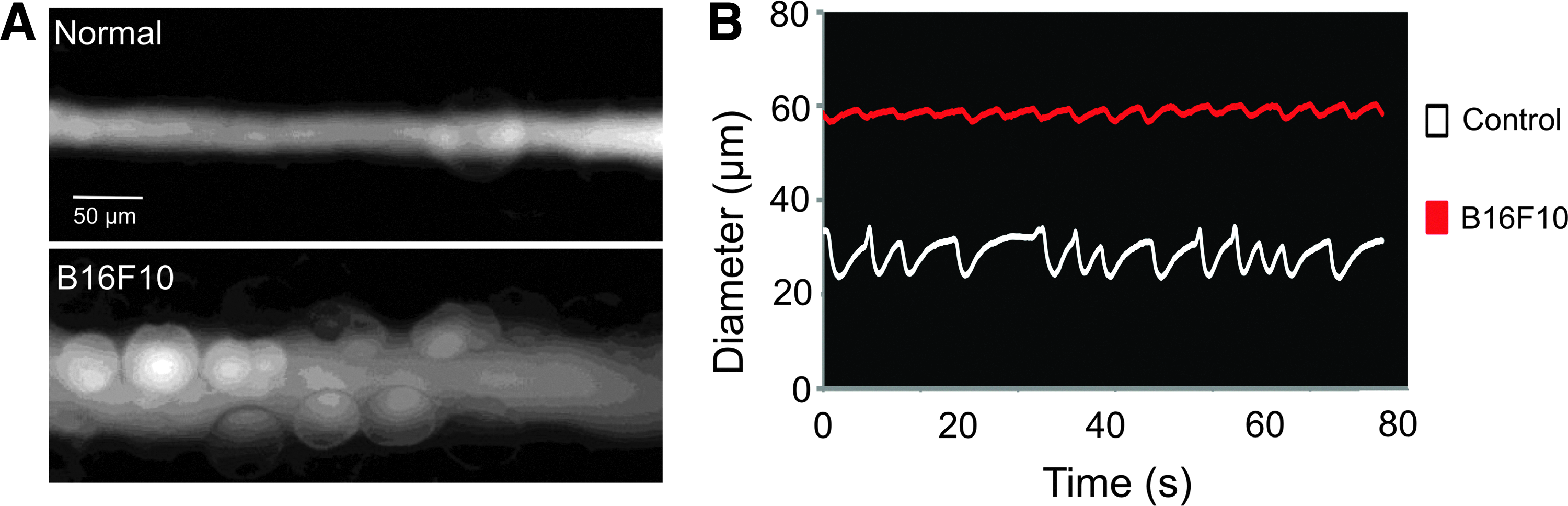
Lymphatic vessels are dilated and lymphatic contraction is inhibited in vessels draining B16F10 melanomas. B16F10 melanoma was implanted into the hind footpad of C57Bl/6 mice. Lymphatic contraction analysis of the afferent lymphatic vessel to the popliteal lymph node was performed in nontumor-bearing control animals
Mean ± standard error of the mean.
p < 0.05 of tumor compared with nontumor-implanted control tested with an unpaired t-test with a Welch's correction.
p < 0.01 of tumor compared with nontumor control tested with an unpaired t-test with a Welch's correction.
p < 0.01 compared with C57BL/6 by an ANOVA with a Tukey's post hoc test.
p < 0.01 compared with nude by an ANOVA with a Tukey's post hoc test.
p < 0.01 compared with HST26T by an ANOVA with a Tukey's post hoc test.
ANOVA, analysis of variance; DT, diphtheria toxin.
iNOS suppresses lymphatic contraction during tumor metastasis
Previously, it was found that iNOS-expressing cells suppress lymphatic contraction during sterile inflammation through NO. 12 Therefore, we tested whether iNOS is involved in suppressing lymphatic contraction during cancer progression by using iNOS−/− mice available on the C57Bl6 background. We observed maintenance of lymphatic contraction strength (ejection fraction) in iNOS−/− mice bearing syngeneic B16F1 and B16F10 tumors (Fig. 2A, B), comparable with nontumor-bearing controls. To determine whether altered lymphatic contraction affects LN metastasis, H&E staining was performed on the TDLNs from wild-type (WT) and iNOS−/− mice implanted with B16F1 and B16F10 (Fig. 2C, D). We found that the LN metastasis rate was not different in WT versus iNOS−/− in both B16F1 and B16F10 models/tumors (Fig. 2E, F), even though lymphatic contraction was maintained in iNOS−/− mice compared with WT mice.
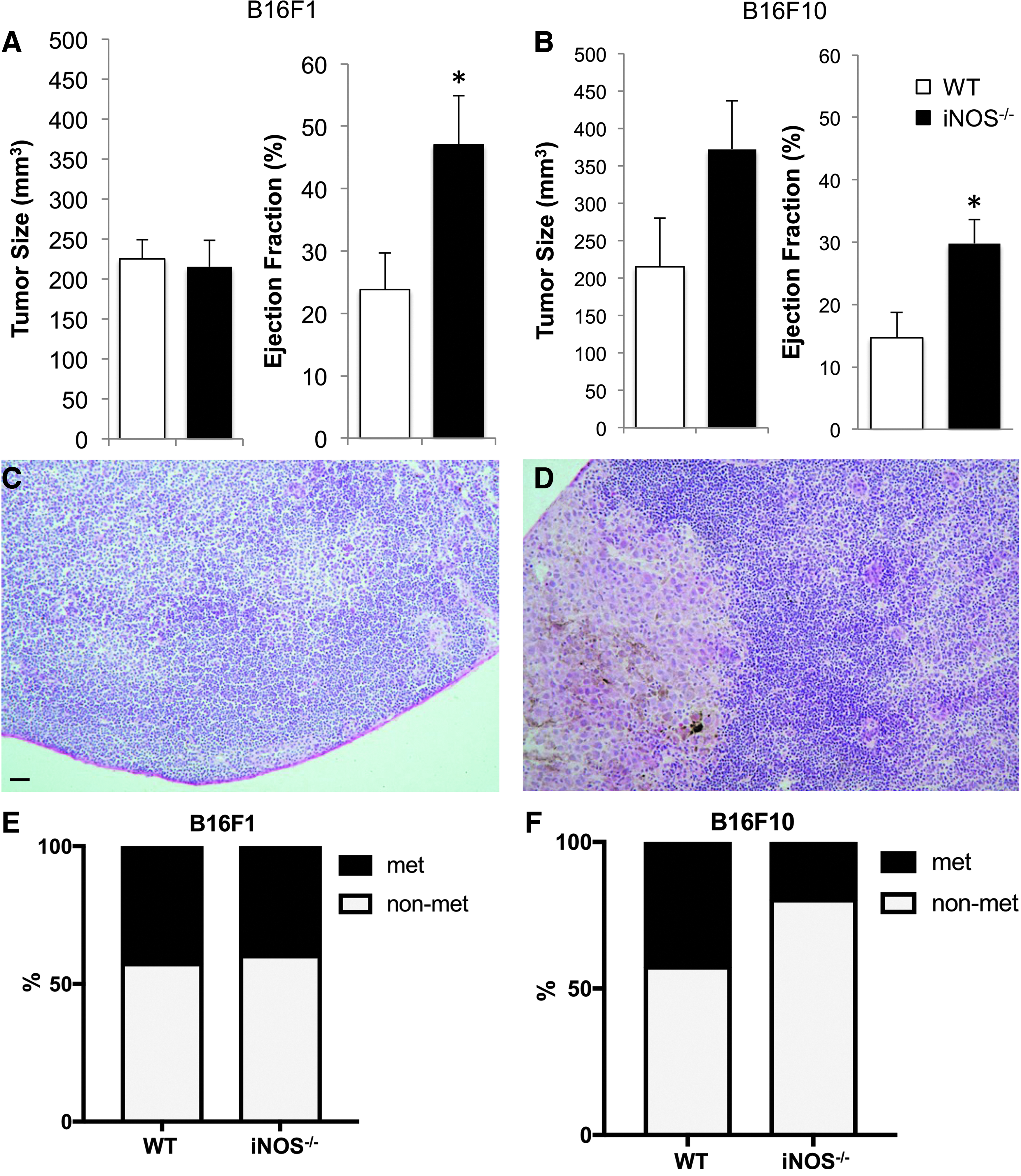
Lymphatic contraction is maintained in iNOS−/− mice. B16F1 or B16F10 melanoma was implanted into the hind footpad of C57Bl/6 WT or iNOS−/− mice before undergoing lymphatic contraction analysis. There is no difference in tumor size between WT and iNOS−/− mice implanted with B16F1 or B16F10 melanoma
Myeloid-derived cells suppress lymphatic contraction in cancer-draining lymphatic vessels
Previously, it was found that CD11b+Gr1+ cells are the major contributors of iNOS-mediated suppression of lymphatic function in sterile inflammation. 12 To confirm the presence of this cell type in tumor models, flow cytometry was performed on B16F10 tumor tissue and it was found that 82.8 ± 2.1% of infiltrated CD45+ cells were CD11b+. Within this population, 40 ± 7% of CD45+CD11b+ were also Gr1+ cells. We further showed that a fraction of these CD11b+ cells were also expressing iNOS and could be responsible for inhibiting lymphatic contraction (Supplementary Fig. S1). Next, we assessed whether depleting Gr1+ cells with an anti-Gr1 antibody restored lymphatic contraction after B16F1 or B16F10 tumor implantation. We found that depleting Gr1+ cells had no effect on tumor growth (Fig. 3A, B), but the strength of lymphatic contraction was greater than that in placebo-treated animals (Fig. 3C). These data show that Gr1+ cells inhibit lymphatic contraction in tumor-bearing animals.
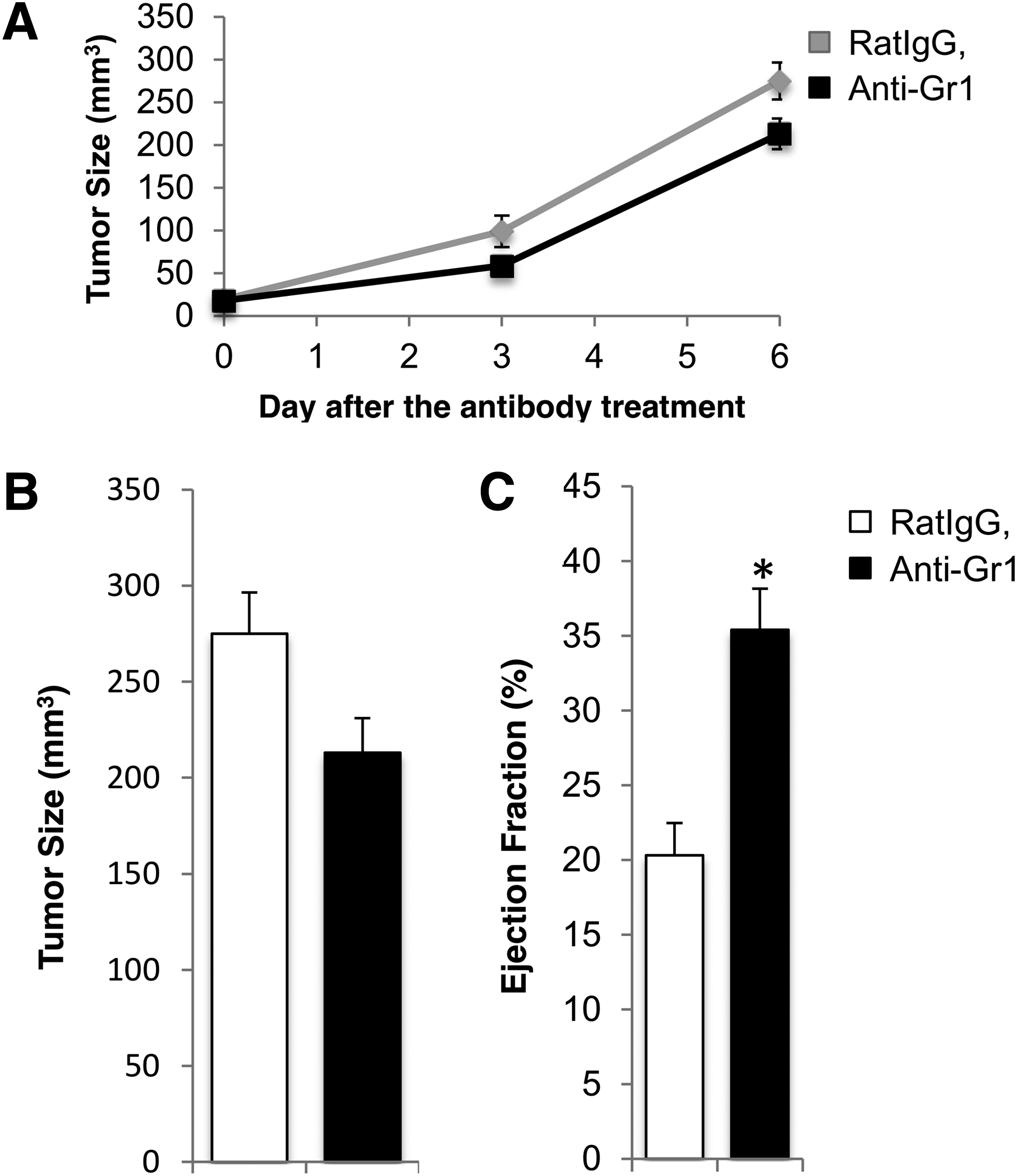
Lymphatic contraction is increased by depleting Gr1 cells. WT C57Bl/6 mice were implanted with B16F10 melanoma and underwent treatment with anti-Gr1 IgG or nonspecific rat IgG (control) to determine whether depletion of Gr1+ cells would restore lymphatic contraction. Anti-Gr1 IgG treatment had no significant effect on B16F10 melanoma growth or tumor size
Induced cancer cell death restores lymphatic contraction
Previously, we found that intratumor lymphatic vessels are collapsed. 28 Therefore, we tested the hypothesis that inducing tumor cell death may open these compressed vessels within the tumor and stimulate lymphatic contraction. To prevent any off-target effects on murine host cells, we took advantage of the differential sensitivity of human and murine cells to DT. 29 HSTS26T human sarcoma tumor chunks were implanted into the footpad in nude mice and were treated with DT to kill the human cancer cells. DT does not cause death of murine cells. 29 PBS injections were used as control. Mice treated with DT had a reduced maximum lymphatic diameter and restored lymphatic ejection fraction (Fig. 4A–C, Table 1), demonstrating that tumor burden itself plays a role in inhibiting lymphatic contraction. This is further confirmed by a correlation between tumor size and ejection fraction (Fig. 4D). Furthermore, these data show that the impairment in lymphatic contraction induced by tumors is reversible; however, studying the contribution of immune cell infiltrates in this model may be compromised because of the use of an immunodeficient mouse strain.
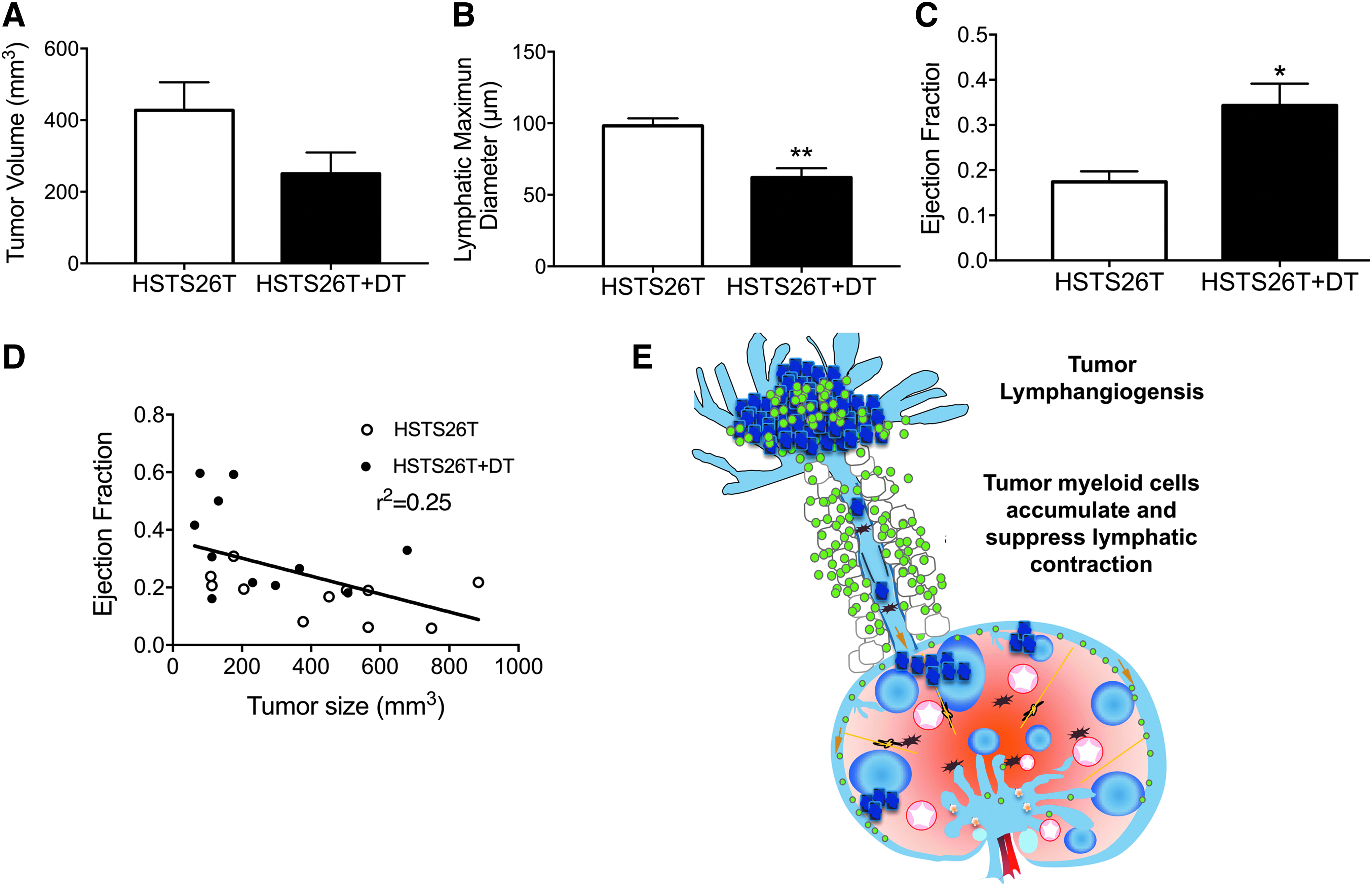
Depleting tumor cells increase lymphatic contraction in HSTS26T human sarcoma. HSTS26T was implanted in the footpad of nude mice and DT was used to reduce tumor burden. DT treatment directly killed tumor cells, reducing tumor size
Conclusion
In this study, we demonstrate that iNOS is a regulator of tumor-draining lymphatic vessels. We found that multiple tumor types reduce contraction in tumor-draining collecting lymphatic vessels (Fig. 1, Table 1), and that iNOS−/− mice were able to maintain a higher ejection fraction than WT mice in melanoma models (Fig. 2). This finding suggests that NO produced by iNOS expressing cells suppresses lymphatic contraction in both sterile inflammation 12 and the tumor setting.
It is well known that tumors recruit various myeloid cells, including Gr1+ cells, that support tumor growth by inducing angiogenesis, lymphangiogenesis, and immune suppression.13,30,31 Although iNOS is expressed in many cells, Gr1+ cells commonly express iNOS in the tumor setting. Therefore, we determined whether depleting Gr1+ cells had an effect on lymphatic contraction. We found that, although Gr1+ cell depletion did not have an effect on tumor growth or size, lymphatic contraction was improved (Fig. 3C). This finding suggests that Gr1+ cells reduce lymphatic contraction during tumor growth, likely through an iNOS-mediated mechanism.
Previously, it was found that tumors induce lymphatic hyperplasia.32,33 We used DT treatment of a human cell line to demonstrate that direct tumor cell death can restore lymphatic contraction (Fig. 4B, C). A correlation was found between tumor size and lymphatic ejection fraction, establishing that tumor burden contributes to a reduction in lymphatic pumping (Fig. 4D). This cumulative data lead to the proposed mechanism (Fig. 4E), wherein both the tumor and the tumor-associated myeloid cells suppress lymphatic contraction. Furthermore, these data show that lymphatic dysfunction in tumor-draining lymphatic vessels is reversible, in contrast to lymphatic vessel dysfunction during and after bacterial infections. 34 Whether the reduction in tumor-draining lymphatic contraction leads to reduced lymph flow and lymphatic drainage requires further study, as no change was measured in lymphatic drainage from perilesion regions of melanomas compared with normal skin in patients. 35
Lymphatic vessels deliver not only tumor cells to the LN but also tumor antigen tumor-derived regulatory factors and dendritic cells. Thus, lymphatic vessels are critical for generating anticancer immune responses in TDLNs. This research highlights the need to further study the relationship between lymphatic function and immune regulation during cancer progression. 36 Future work is needed to understand the complex relationship between lymphatic contraction, metastasis, and antitumor immunity.
Footnotes
Acknowledgments
This work was supported by NIH DP2 OD008780 and NIH R01 HL128168. S.L. was supported by Canadian Institutes of Health Research Bridge fund 371997. E.M.B. was supported by the LE&RN/FDRS Lipedema Postdoctoral Fellowship.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.