
Abstract
Abstract
Background:
Mesenteric lymph (ML) has been implicated in the development of multiple organ dysfunction syndrome in critical illness. Extracellular RNAs play a role in cell-to-cell communication during physiological and disease processes but they are rarely studied in ML. We aimed at examining the RNA profiles of peripheral plasma, ML, and ML's extracellular vesicle (ML-EV) and triglyceride-rich lipoprotein (ML-TRL) fractions, obtained from rodent models of critical illness.
Methods and Results:
We collected ML for 5 hours from rodent models of critical illness [Acute Pancreatitis, Cecal Ligation and Incision (CLI), Gut Ischemia-Reperfusion (IR)] and matching Sham control rats. ML-EV and ML-TRL fractions were also isolated. RNA sequencing was performed on the RNA extracted from ML, ML-EV, ML-TRL, and plasma by using the Ion Torrent Personal Genome Machine platform. RNA sequences were searched using the Basic Local Alignment Search Tool against rat genome and RefSeq, microRNA (miRNA), genomic tRNA, functional RNA, and Genbank nucleotide databases, and the read counts were analyzed. Each sample type had a distinct RNA profile. ML contained more RNA per volume and a larger proportion of tRNA fragments than plasma. ML-EVs were enriched with miRNA, whereas ML-TRLs contained low absolute amounts of RNA. The RNA size profiles for CLI and Gut IR were different from Sham. ML carried intestinal RNAs and in a CLI model it was significantly enriched with bacterial RNA sequences.
Conclusions:
We found the distinct but diverse RNA profiles of ML and its compartments, and their different profiles in critical illness. Intestinal-derived small RNAs in ML may have a direct role in critical illness and utility as potential biomarkers.
Introduction
Various regulatory RNA species 1 are stable outside the cell 2 and play roles in cell-to-cell communication 3 and other biological or pathological processes.2,3 Such extracellular RNAs, 1 with the most studied being microRNAs (miRNA),2,3 have been found in a variety of biofluids,4,5 where they can be packaged with extracellular vesicles (EV), 6 lipoproteins, 7 and other ribonucleoprotein complexes. 3
The lymphatic system is an organized network of lymphoid tissue; it transports tissue fluid/lymph and lymphoid cells, 8 and various pathological conditions such as inflammation, cancer, and infection can cause alterations in the lymphatic function. 9 Mesenteric lymph (ML) continuously drains from the gut and re-enters the circulation at the subclavian vein just before the heart and lungs. ML follows a direct anatomical path between the gut and peripheral bloodstream without passing through the liver. In health, ML is essential in fluid homeostasis and dietary lipid absorption. Triglyceride-rich lipoproteins (TRL) are the major lipoproteins within ML that transport intestinal lipids to the bloodstream. In disease, ML is emerging as an important factor, 10 and it has been implicated in the development of multiple organ dysfunction syndrome in critical illness. 10 Although miRNAs have been detected within pancreatitis ML of the rodent model and patients using microarray approaches, 11 other extracellular RNAs or other types of critical illness are yet to be studied.
Here, we examined the small RNA profiles of plasma and ML collected from different rodent models of critical illness [Acute Pancreatitis, Cecal Ligation and Incision (CLI), Gut Ischemia-Reperfusion (IR)] for the first time by using Ion Torrent sequencing.
Materials and Methods
Refer to Supplementary Data for complete details. Briefly, Acute Pancreatitis, CLI, and Gut IR were induced in the adult male Sprague-Dawley rats. Sham control rats underwent the same intervention, but without any disease induction. For the drainage models, the superior ML duct was cannulated to collect ML continuously on ice over 5 hours from rats. A peripheral plasma sample was collected from all the study rats at a single time point on completion of the 5 hours lymph collection. For CLI and Gut IR, EV- and TRL-enriched fractions were isolated from ML (ML-EV; ML-TRL) by using Macherey-Nagel exosome precipitation solution and differential ultracentrifugation, respectively. Small RNA sequencing was performed on RNA extracted from pooled samples of plasma (N = 3), ML (N = 3), ML-EV (N = 4), and ML-TRL (N = 5) on the Ion Torrent Personal Genome Machine platform. RNA sequences were filtered and searched using the Basic Local Alignment Search Tool (BLAST) against various databases, including rat genome and RefSeq, miRNA, genomic tRNA, functional RNA, and Genbank nucleotide databases. RNA sequences were classified, and the read counts were analyzed.
Results and Discussion
There were distinct differences in the RNA profiles between ML and plasma. Recently, differences in small RNA composition between 12 human biofluids were characterized by Godoy et al., 5 but ML was not investigated. ML contained more RNA per volume than plasma with both total input and BLAST-matched read counts being >20-fold higher (Fig. 1A; Supplementary Table S1). Although >90% of usable reads in both ML and plasma were identified as being of “rat” origin, their RNA subtypes occurred in different proportions (Fig. 1B). The most notable difference was in the tRNA proportion: >90% occurred in ML relative to ∼45% in plasma for Sham rats, with the majority being tRNA-halves (∼32 nt; half of full-length tRNA). Among various tRNA-halves, the most abundant tRNA-half detected in this study was the one derived from 5′ end of Gly-GCC (tRNA with GCC anticodon recognize the GCC codon, which codes for glycine): mean 48% of total rat tRNA in ML and 69% in plasma of Sham rats (Fig. 1C). Over-representation of tRNA fragments could be an artefact of conventional RNA-sequencing methods,12–14 but other studies15,16 have shown that various stress conditions can induce tRNA cleavage, and these tRNA fragments serve as small interfering RNAs that regulate the activity of translation factor.
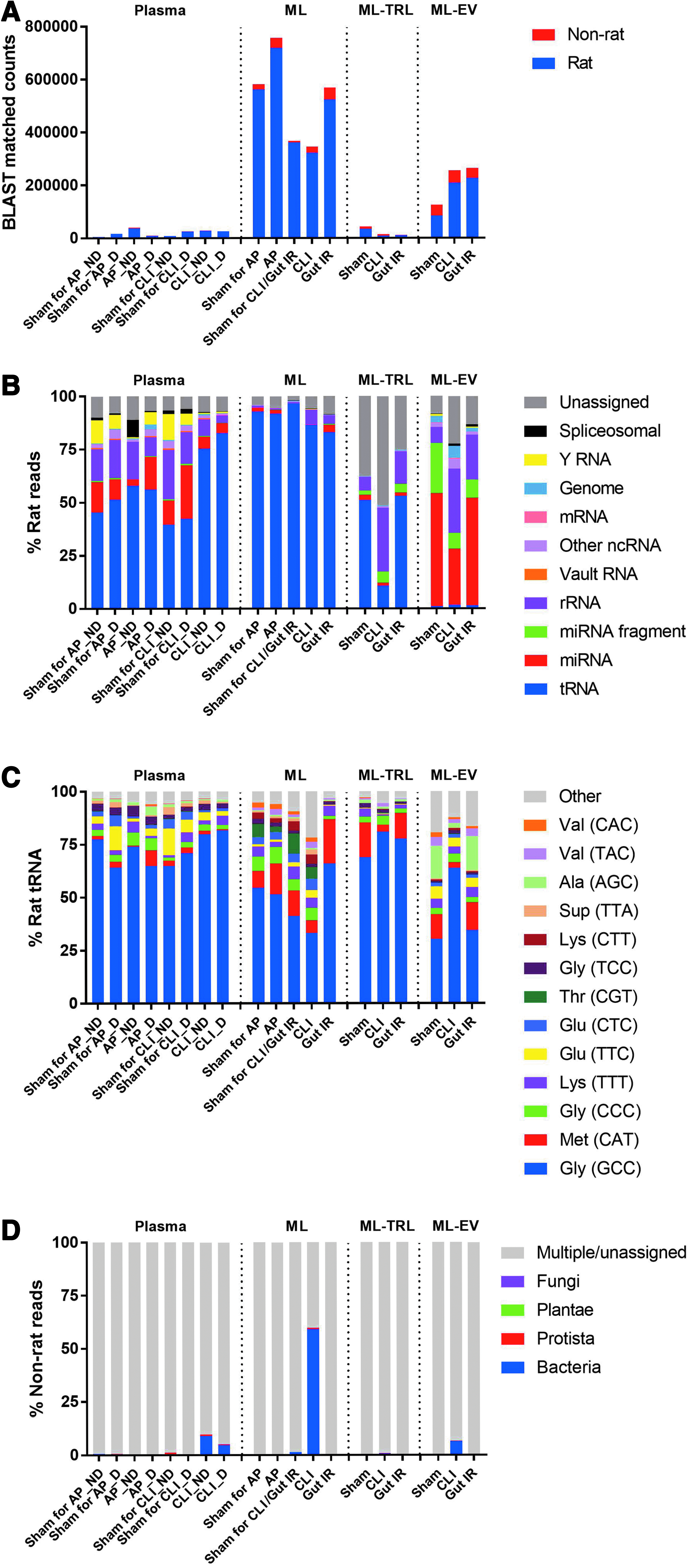
Extracellular RNA profiles of plasma, ML, ML-TRL, and ML-EV in the rodent models of critical illness. Rodent models of AP, CLI, and Gut IR, and their Sham controls were compared. For AP and CLI, plasma RNA profiles, ML ND, and D models were also compared.
In contrast to the tRNAs, plasma contained a greater proportion of miRNAs, rRNA fragments, and Y RNAs than the ML (Fig. 1B). Of the top 10 mature miRNAs identified (Supplementary Table S2), the most predominant in plasma and ML were miR-451-5p and miR-145-5p, respectively; whereas miR-16-5p was abundant in all plasma and ML samples. miR-143-3p and miR-3473 were highly abundant in ML but very low in plasma. miR-143/miR-145 are co-transcribed miRNAs that have enriched expression in the intestinal mesenchyme and are shown to have a role in intestinal wound repair, 17 and miR-3473 had an increased expression in inflamed large intestinal epithelial cells in a mouse model of colitis, 18 thus confirming that ML carries intestinal origin miRNAs.
Comparison between ML, ML-TRL, and ML-EV from Sham controls detected differences in RNA profiles between different ML compartments. tRNA-halves, which occupied >90% of total rat reads in unfractionated ML, were present as only 1% of the ML-EV reads (Fig. 1B). Instead, ML-EV was highly enriched with miRNAs (76% of total rat reads), of which 30% were short fragments (mean 17 nt) of the annotated mature miRNAs (mean 22 nt) (Fig. 1B). ML-EV also contained a higher proportion of rRNA- and genome-matched reads than the unfractionated ML (Fig. 1B). In comparison to ML-EV, Sham ML-TRL contained less miRNAs (2.4% of total rat reads) and their fragments (2%), but more tRNA-halves (51%) (Fig. 1B). However, overall ML-TRL samples contained low absolute amounts of RNA, suggesting that TRLs are not a major RNA carrier in ML.
ML and its TRL- and EV-enriched fractions also showed differences in their miRNA contents (Supplementary Table S2). miR-22219 and miR-15019,20 previously identified in EVs were enriched in both ML-TRL and ML-EV in comparison to the unfractionated ML, whereas miR-145-5p was consistently abundant in all ML-derived samples. miR-3473, found to be enriched in unfractionated ML, was ranked either #1 or #2 in ML-TRL samples, but low in ML-EV. miR-125a-5p that has a role in intestinal epithelial cell fate 21 was distinctly enriched in ML-EV. Of interest, the most abundant miRNA fragment in both ML-TRL and ML-EV was matched antisense to miR-222-3p and we hypothesize that it may represent a new miRNA and/or have a role to bind and inhibit the function of miR-222-3p.
Lastly, we compared the RNA profiles of critical illness models. There was no substantial difference in the RNA profiles of both plasma and ML between Acute Pancreatitis and its respective Sham (Fig. 1B), but we could detect increased abundance of miR-217-5p (112 vs. 0 reads per million), miR-375-3p (12 vs. 0), miR-122-5p (99 vs. 29), and miR-148a (126 vs. 43) in ML of Acute Pancreatitis than its Sham, confirming the previous miRNA findings by Blenkiron et al. 11 In contrast, we detected more changes in the RNA profiles for CLI and Gut IR compared with Sham. In CLI, there was a substantial increase of ∼32 nt reads in the plasma, irrespective of ML drainage status (Fig. 2A), and this correlated well with increased tRNA-halves (Fig. 1B). In particular, Gly-GCC tRNA-halves were found to be more abundant in the CLI plasma than Sham plasma (614,868 vs. 267,526 reads per million). The Gly-GCC tRNA-halves were previously shown to increase in ischemia and inhibited the function of endothelial cells, 22 and tRNA type-specific biogenesis and/or release of 5′ tRNA halves has been reported, 23 but the field of tRNA fragment biology remains an interesting new research area and warrants further studies.
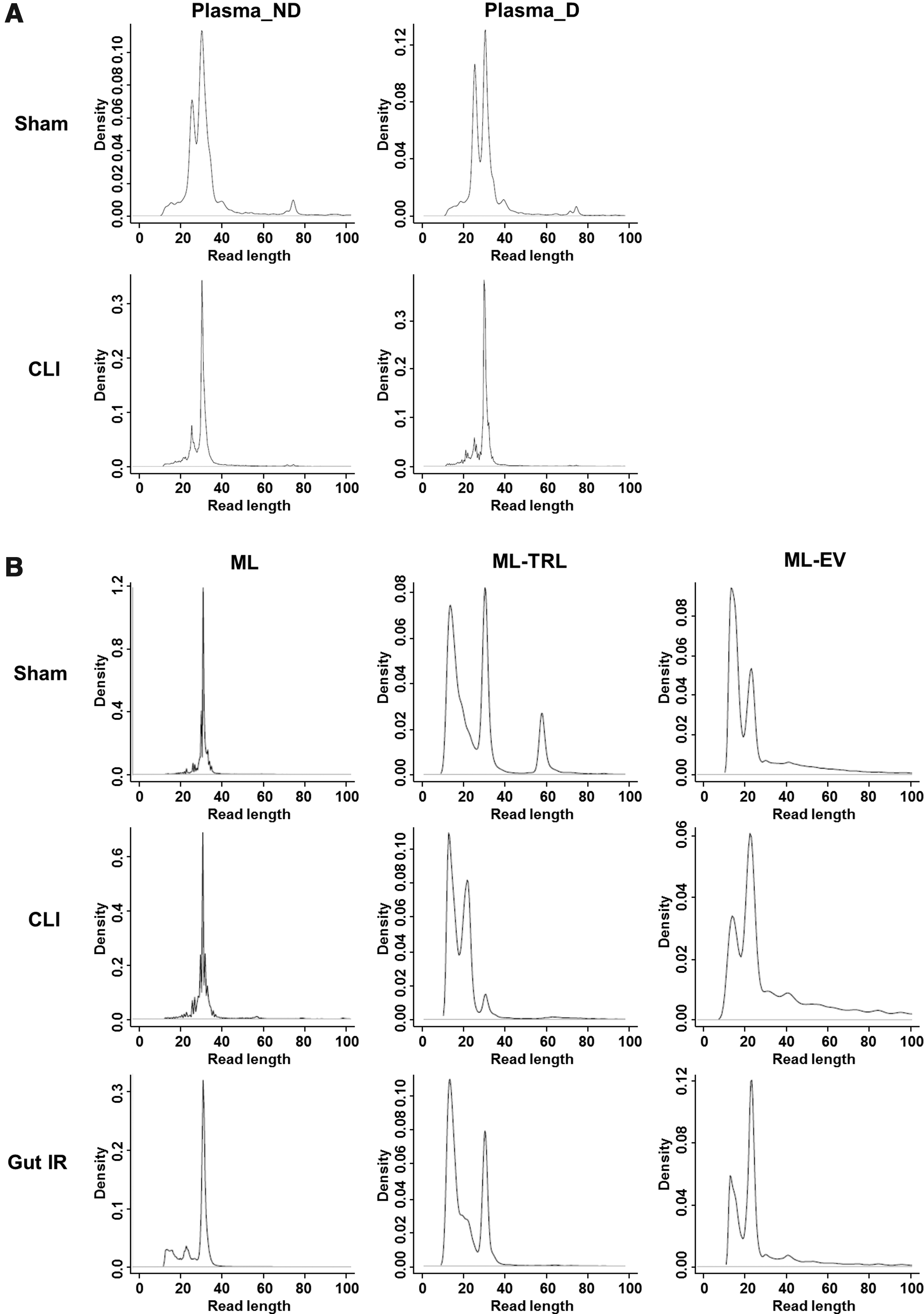
Comparison of size profile of RNA reads between Sham, CLI, and Gut IR
Read length size profiles of ML were not substantively different between Sham, CLI, and Gut IR (Fig. 2B), and the majority of RNA were constantly matched to tRNA-halves (Fig. 1B). However, the RNA size profiles of ML-TRL and ML-EV did differ between Sham, CLI, and Gut IR (Fig. 2B). BLAST-matched results showed more distinct RNA profile changes in the CLI than Gut IR, compared with the Sham (Fig. 1B, C). In all ML-derived samples (ML, ML-TRL, and ML-EV), the % rRNA-matched short reads were consistently the highest in CLI, lower in Gut IR, and the lowest in Sham (Fig. 1B). Previous studies24,25 have considered rRNA-matching short reads as degraded products and excluded them from further analysis. This is in contrary to other studies that have suggested their possible roles as DNA damage-induced qiRNAs, 26 small guide RNAs, 27 or diRNAs; 28 as well as proposed their interaction with translation factors, 29 mRNA and tRNA analogues, or antibiotics. 30 In addition, our finding may support a potential role of rRNA-derived small RNAs in critical disease processes, as observed in diabetes. 31 Small rRNAs are now deserving further investigation for these pathological roles.
The other interesting observation was the detection of bacterial sequences in CLI (Fig. 1D; Supplementary Table S3). Overall, 59% of total “non-rat” sequences in CLI ML matched bacteria, 6% in ML-EV, and 9% in the plasma of the non-ML drainage model, which reduced to 5% in the plasma of the ML drainage model. In contrast, minimal levels of bacterial sequences (0%–1.3%) were detected in all the samples from other disease models. BLAST-searches showed that 61% of bacterial sequences found in CLI ML matched the Enterobacteriaceae family; a normal component of the gut microbiota in rats, but only accounting for 9% of typical abundance. 32 The dominance of Enterobacteriaceae in CLI ML suggests preferential transfer of this taxa's RNAs from the intraperitoneal feces (dispersed within the abdomen as part of the CLI induction) into the visceral lymphatic channels of the peritoneum. The prevention of bacterial RNA transfer from ML to systemic circulation may represent a method to reduce disease severity.
Conclusion
This is the first study to report the distinct but diverse RNA profiles of ML and its compartments (TRL and EV) and their different profiles in critical illness. Our results show carriage of intestinal small RNAs in all ML, enrichment of miRNAs in ML-EVs, and the presence of bacterial RNA in CLI ML. These intestinal-derived small RNAs may have a direct role in disease states, adding a new perspective to ML as a potential gut signaling axis and biomarker source, in the pathophysiology of critical illness.
Footnotes
Acknowledgments
The authors would like to thank Prof. Cristin Print for his advice on the bioinformatics analysis, and Liam Williams and Tim Lawrence in the Genomic Centre, University of Auckland (in conjunction with New Zealand Genomics Ltd.) for their sequencing services. Funding: This work was supported by the Faculty Research Development Fund and the Performance-Based Research Fund from the University of Auckland, the Auckland Medical Research Foundation Project Grant, the Maurice and Phyllis Paykel Trust Project Grant, and the Health Research Council of New Zealand Project Grant. J.H. was funded by the Hugo Charitable Trust.
Data Availability
The raw data files are available from the NCBI's Sequence Read Archive (SRP114999).
Authors' Contributions
J.H. and C.B. participated in study design, data collection and analysis, interpretation of data, and article preparation. P.T. participated in data collection and analysis, interpretation of data, and article preparation. R.P., S.N., S.M.T., and A.P. participated in rat surgery, sample collection, and article preparation. A.R.P., A.J.H., and J.A.W. participated in study design, article preparation, and editorial oversight.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.