
Abstract
Abstract
Aim:
To investigate the expression of embryonic stem cell (ESC) markers in microcystic lymphatic malformation (mLM).
Methods and Results:
Cervicofacial mLM tissue samples from nine patients underwent 3,3′-diaminobenzidine (DAB) immunohistochemical (IHC) staining for ESC markers octamer-binding protein 4 (OCT4), homeobox protein NANOG, sex determining region Y-box 2 (SOX2), Krupple-like factor (KLF4), and proto-oncogene c-MYC. Transcriptional activation of these ESC markers was investigated using real-time polymerase chain reaction (RT-qPCR) and colorimetric in situ hybridization (CISH) on four and five of these mLM tissue samples, respectively. Immunofluorescence (IF) IHC staining was performed on three of these mLM tissue samples to investigate localization of these ESC markers. DAB and IF IHC staining demonstrated the expression of OCT4, SOX2, NANOG, KLF4, and c-MYC on the endothelium of lesional vessels with abundant expression of c-MYC and SOX2, which was also present on the cells within the stroma, in all nine mLM tissue samples. RT-qPCR and CISH confirmed transcriptional activation of all these ESC markers investigated.
Conclusions:
These findings suggest the presence of a primitive population on the endothelium of lesional vessels and the surrounding stroma in mLM. The abundant expression of the progenitor-associated markers SOX2 and c-MYC suggests that the majority are of progenitor phenotype with a small number of ESC-like cells.
Introduction
Lymphatic malformation (LM) affects 1:5000 births 1 with 50% diagnosed prenatally or at birth and the remainder become apparent as a slowly increasing swelling as the child grows. 2 Seventy-five percent of LM affects the head and neck area with the remainder involving the axilla, chest, and perineum in decreasing frequencies. 3 LM is categorized according to its morphology as macrocystic (lumen >1 cm) or microcystic (lumen <1 cm) or mixed. 4 It may be localized or diffuse. 5 Histologically, LM appears as irregular thin-walled ectatic channels lined by flat endothelial cells with few pericytes surrounded by a fibrous stroma. 1
LM may be complicated by infection that can lead rapidly to septicemia, intralesional bleeding, 6 ulceration, and visual or airway obstruction.3,7
Treatment for LM remains unsatisfactory, particularly for extensive microcystic lesions. The majority of LMs are partially or wholly microcystic with the lesion infiltrating normal tissue. Sclerotherapy is the preferred treatment for large macrocystic lesions, using OK-432, 8 doxycycline, 9 sodium tetradecyl sulfate, 10 ethanol, 11 or bleomycin. 12 Surgical debulking is used for lesions unsuitable for sclerotherapy 3 and recurrence is common.3,7
The pathogenesis of LMs is poorly understood. Some cases are associated with gene mutations, 13 but the etiology remains unknown for the majority of the cases. Wu et al. 7 have described a population of cells on the endothelium as well as cells within the surrounding stroma that express progenitor cell marker CD133, endothelial precursor proteins CD90, CD146, c-Kit, and VEGFR-2, and transcription activation of octamer-binding protein 4 (OCT4) and homeobox protein NANOG, and they suggest the presence of a progenitor cell population within LM. We have recently demonstrated an embryonic stem cell (ESC)-like population on the endothelium of the lesional vessels as well as cells within the stroma, in subcutaneous and intramuscular venous malformation (VM). 14
ESCs are characterized by pluripotency and self-renewal. The search for markers of ESCs started with the isolation of cells from the inner cell mass of human blastocysts. 15 Since then many transcription factors, cell surface markers, peptides, and lectins have been associated with ESCs. 16 Induced pluripotent stem cells (iPSCs), with the hallmark features of ESCs, have been generated from mouse fibroblasts using OCT4, sex determining region Y-box 2 (SOX2), Krupple-like factor (KLF4), and proto-oncogene c-MYC. 17 This finding was reproduced in human somatic cells with the addition of NANOG. 18 OCT4 belongs to the OCT family of POU transcription factors. High expression of OCT4 occurs during embryogenesis and decreases with cellular differentiation. 19 It regulates the expression of a number of genes, including SOX2 and NANOG, which are active in early development.20,21 Sry-related high-mobility group box-containing (SOX) family of transcription factors are key mediators of pluripotency. 22 SOX2 expression is associated with multipotent cells as opposed to OCT4, which is exclusive to pluripotent cells. 16 NANOG is a transcription factor considered the third key maintainer of the ESC identity, being closely aligned with OCT4 and SOX2. 20 It is expressed by pluripotent cells and downregulates differentiation. 16 The KLF4 family are transcription factors important in a number of processes, notably cell proliferation, differentiation, and apoptosis.16,21 Along with SOX2 and OCT4, it is indispensable for cell reprogramming.17,18 c-MYC belongs to the family of MYC genes and plays a key role in cell proliferation. 20 It is thought that c-MYC is associated with progenitor cells rather than pluripotent stem cells, given that iPSCs have been generated without c-MYC. 23
This study aimed to identify and characterize a primitive population within microcystic LM (mLM) using the ESC markers OCT4, SOX2, NANOG, KLF4, and c-MYC at both the transcriptional and translational level using 3,3′-diaminobenzidine (DAB) and immunofluorescence (IF) immunohistochemical (IHC) staining, reverse transcription quantitative polymerase chain reaction (RT-qPCR), and colorimetric in situ hybridization (CISH).
Materials and Methods
Tissue samples
Cervicofacial mLM tissue samples from nine patients were sourced from the Gillies McIndoe Research Institute Tissue Bank and used for a study that was approved by the Central Health and Disability Ethics Committee (Ref. 13/CEN/130). Written informed consent was obtained from all participants.
Histology and IHC staining
The presence and correct diagnosis of mLM were confirmed by an anatomical pathologist (H.D.B.) by hematoxylin and eosin (H&E) staining and DAB IHC staining for D2-40 (1:100, cat# M3619; Dako, Glostrup, Denmark) on 4 μm-thick formalin-fixed paraffin-embedded sections of nine mLM tissue samples. DAB IHC staining with primary antibodies NANOG (1:100, cat# ab80891; Abcam, Cambridge, UK), OCT4 (1:30, cat# MRQ-10; Cell Marque, Santa Cruz, CA), SOX2 (1:200, cat# PA1-094; Thermo Fisher Scientific, Waltham, MA), c-MYC (1:1000, ca# 9E10; Abcam), and KLF4 (1:200, cat# NBP2-24749SS; Novus Biologicals LLC, Littleton, CO) was performed on the mLM tissue sections using the Leica Bond Rx auto-stainer (Leica), as previously described. 24 To confirm coexpression of two proteins, IF IHC staining was performed on two representative mLM tissue samples from the original cohorts of nine patients included for DAB IHC staining using the same primary antibodies at the same concentrations. Appropriate secondary antibodies such as Vectafluor Excel antirabbit 594 (ready-to-use, cat# VEDK-1594; Vector Laboratories) combinations were used for IF IHC detection. All antibodies were diluted with Bond TM primary antibody diluents (cat# AR9352; Leica). All IHC experiments were performed as single runs.
Positive human control tissues were seminoma for NANOG and OCT4, normal skin for SOX2, breast carcinoma for KLF4, and prostatic tissue for c-MYC.
Colorimetric in situ hybridization
Four micrometers-thick formalin-fixed paraffin-embedded sections of five mLM tissue samples from the original cohort of nine patients included in the DAB IHC staining were subjected to CISH mRNA analysis. Staining was performed using the Leica Bond Rx autostainer with probes for c-MYC (NM_002467.4), KLF4 (NM_001314052), NANOG (NM_024865.2), OCT4 (NM002701.4), and SOX2 (NR_075091.1). All probes used for CISH were obtained from Advanced Cell Diagnostics (Newark, CA). Probes were detected using the RNAscope 2.5 LS Reagent Brown Kit (cat# 322100; Advanced Cell Diagnostics).
Positive controls for OCT4, NANOG, and SOX2 were demonstrated on sections of human seminoma, c-MYC on prostatic tissue, and KLF4 on breast carcinoma. Negative controls were demonstrated on sections of mLM using a probe for DapB (ED191515) (cat# 3120358; Advanced Cell Diagnostics).
Microscopy and image analysis
All DAB IHC-stained slides were viewed and the images were captured using an Olympus BX53 light microscope fitted with an Olympus SC100 digital camera (Olympus, Tokyo, Japan). IF IHC-stained slides were viewed and the images were captured using an Olympus FV1200 biological confocal laser-scanning microscope (Tokyo, Japan). All IF images were subjected to 2D deconvolution with cell Sens Dimension 1.11 software (Olympus).
Reverse transcription quantitative polymerase chain reaction
Total RNA was isolated from 20 mg each of the four snap-frozen mLM tissue samples from the original cohort of nine patients included for DAB IHC staining. The tissue sections were suspended in 350 μL of RLT lysis buffer (cat# 79216; Qiagen, Hilden, Germany) and homogenized using the Omni Tissue Homogenizer (Omni International, Kennesaw, GA). Homogenized samples were then prepared using the RNeasy Mini Kit (Qiagen) then QIAcube (Qiagen). RNA was also isolated from PC3 cells as a positive control. Total RNA quantity and quality were determined using the NanoDrop 2000 Spectrophotometer (Thermo Fisher Scientific). All extractions were performed in triplicates. Extracted RNA was subjected to RT-qPCR using the Rotor-Gene Q (Qiagen). The expression of the ESC markers was detected using gene-specific TaqMan (cat# 4331182; Thermo Fisher Scientific) primers probes: SOX2 (Hs01053049_s1), KLF4 (Hs00358836_m1), NANOG (Hs04399610_g1), OCT4 (Hs00999632_g1), and c-MYC (Hs00153408_m1). All measurements were performed in triplicates and the values were compared against the endogenous housekeeping gene GADPH (Hs99999905_m1, cat# 4351370; Thermo Fisher Scientific). Graphs were generated using Microsoft Excel and the results are presented as relative expression.
Statistical analysis
To determine statistically significant differences, paired samples t-tests were carried out using SPSS V.24.
Results
Histology and DAB IHC staining
mLM was characterized by irregular ectatic vessels lined with flat endothelial cells, surrounded by pericytes on H&E staining (Fig. 1A) and by positive staining for the lymphatic marker D2-40 25 (Fig. 1B, brown) on the lesional vessels, all nine mLM samples. DAB IHC staining also demonstrated mainly cytoplasmic with some perinuclear expression of OCT4 (Fig. 1C, brown), predominantly on the endothelium of the lesional vessels and a few cells within the stroma of mLM. Abundant cytoplasmic and nuclear expression of SOX2 (Fig. 1D, brown) were seen on the endothelium of the lesional vessels and cells within the stroma, whereas weak staining of NANOG (Fig. 1E, brown) was observed on the endothelium of the lesional vessels in all samples. Predominant cytoplasmic expression of KLF4 (Fig. 1F, brown) was seen on the endothelium of the lesional vessels. Abundant nuclear expression of c-MYC (Fig. 1G, brown) was present on the endothelium of the lesional vessels and cells within the stroma in all nine samples.
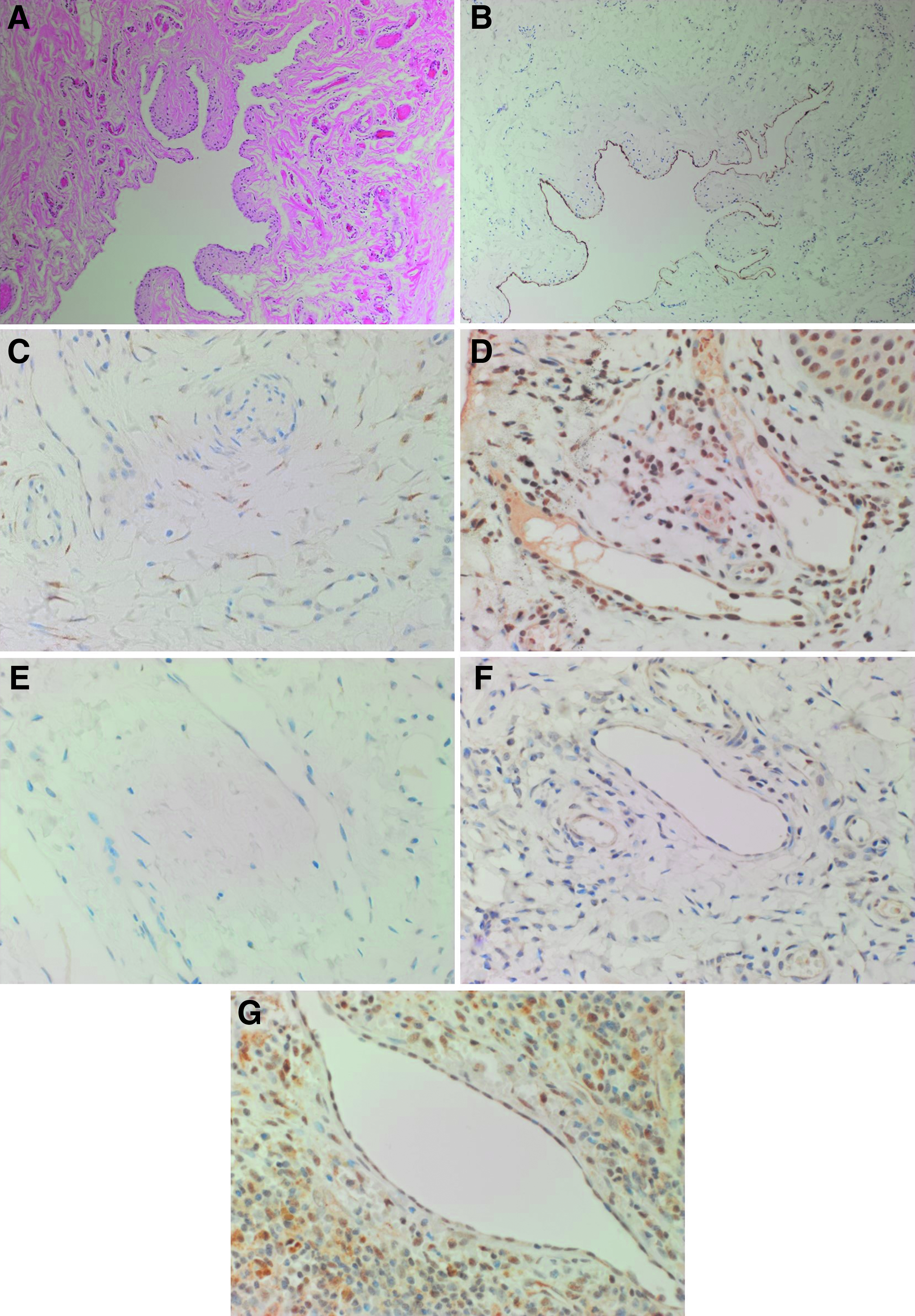
A representative hematoxylin and eosin-stained section of microcystic lymphatic malformation showing characteristic ectatic lymphatic channels lined with flat endothelial cells
Expected positive staining patterns for OCT4 (Supplementary Fig. S1A, brown), NANOG (Supplementary Fig. 1B, purple), SOX2 (Supplementary Fig. S1C, brown), c-MYC (Supplementary Fig. S1D, brown), and KLF4 (Supplementary Fig. S1E, purple) were demonstrated on human seminoma, normal skin, breast carcinoma, and prostatic tissue, respectively. The negative control was an mLM tissue sample that had undergone DAB IHC staining using an IgG isotype (Supplementary Fig. S1F).
IF IHC staining
IF IHC staining demonstrated strong nuclear staining of SOX2 (Fig. 2A, red) by the D2-40+ (Fig. 2A, green) endothelium of the lesional vessels and cells within the stoma. The SOX2+ (Fig. 2B, red) cells showed cytoplasmic expression of OCT4 (Fig. 2B, green) primarily on the endothelium of the lesional vessels. Cytoplasmic and nuclear expression of NANOG (Fig. 2C, red) and cytoplasmic expression of KLF4 (Fig. 2D, red) were present on the D2-40+ (Fig. 2C, D, green) endothelium of the lesional vessels. The SOX2+ (Fig. 2E, red) endothelium of the lesional vessels also expressed c-MYC (Fig. 2E, green).
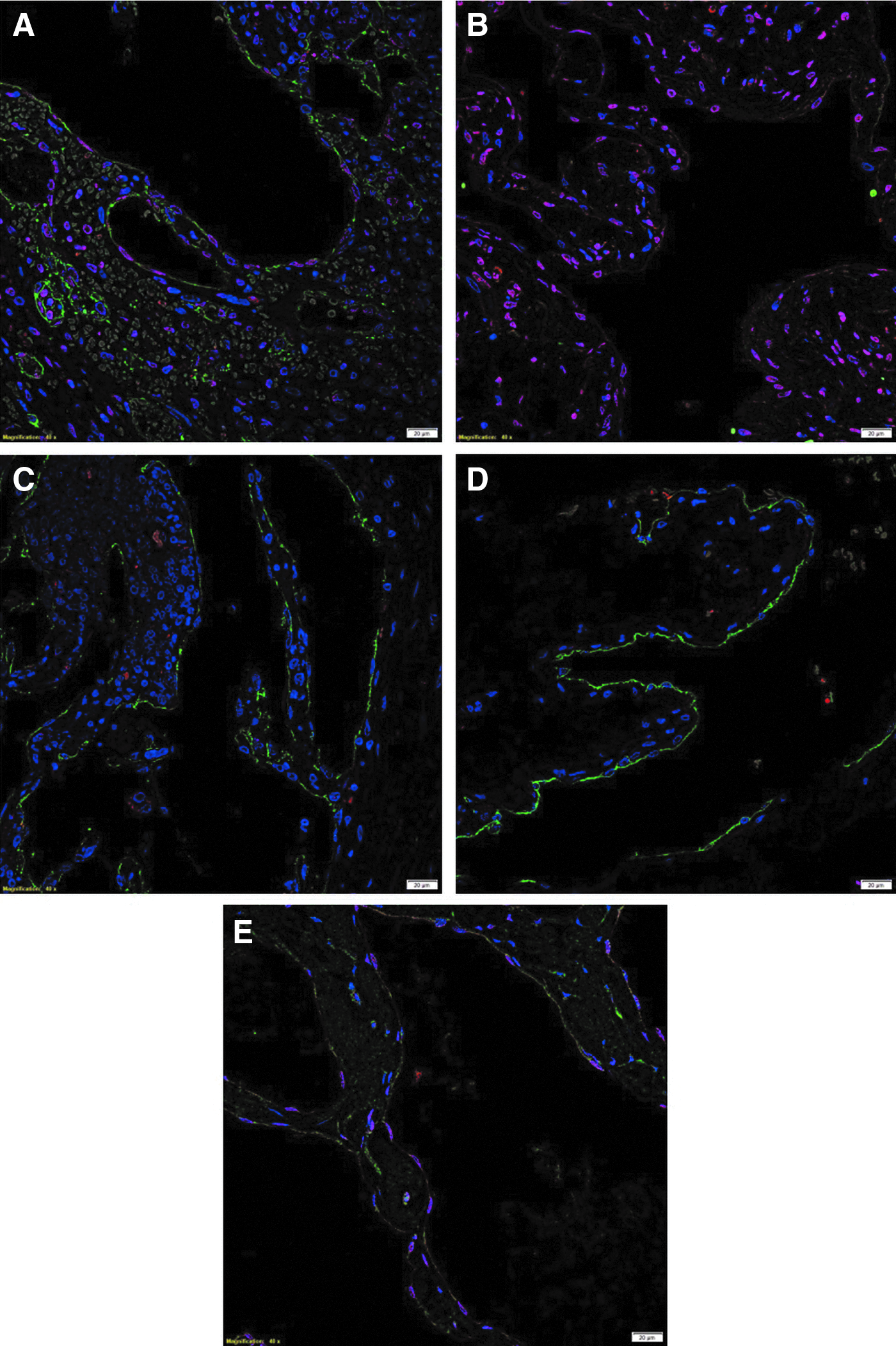
Representative immunofluorescence IHC-stained sections of microcystic lymphatic malformation demonstrating nuclear expression of SOX2 (
Split images of the stains presented in Figure 2 are shown in Supplementary Figure S2. Minimal staining was present on the negative control (Supplementary Fig. S2K), confirming the specificity of the primary antibodies used.
Colorimetric in situ hybridization
CISH demonstrated abundant expression of mRNA transcripts for OCT4 (Fig. 3A, brown, arrows), SOX2 (Fig. 3B, brown, arrows), NANOG (Fig. 3C, brown, arrows), KLF4 (Fig. 3D, brown, arrows), and c-MYC (Fig. 3E, brown, arrows) in all five mLM tissue samples.
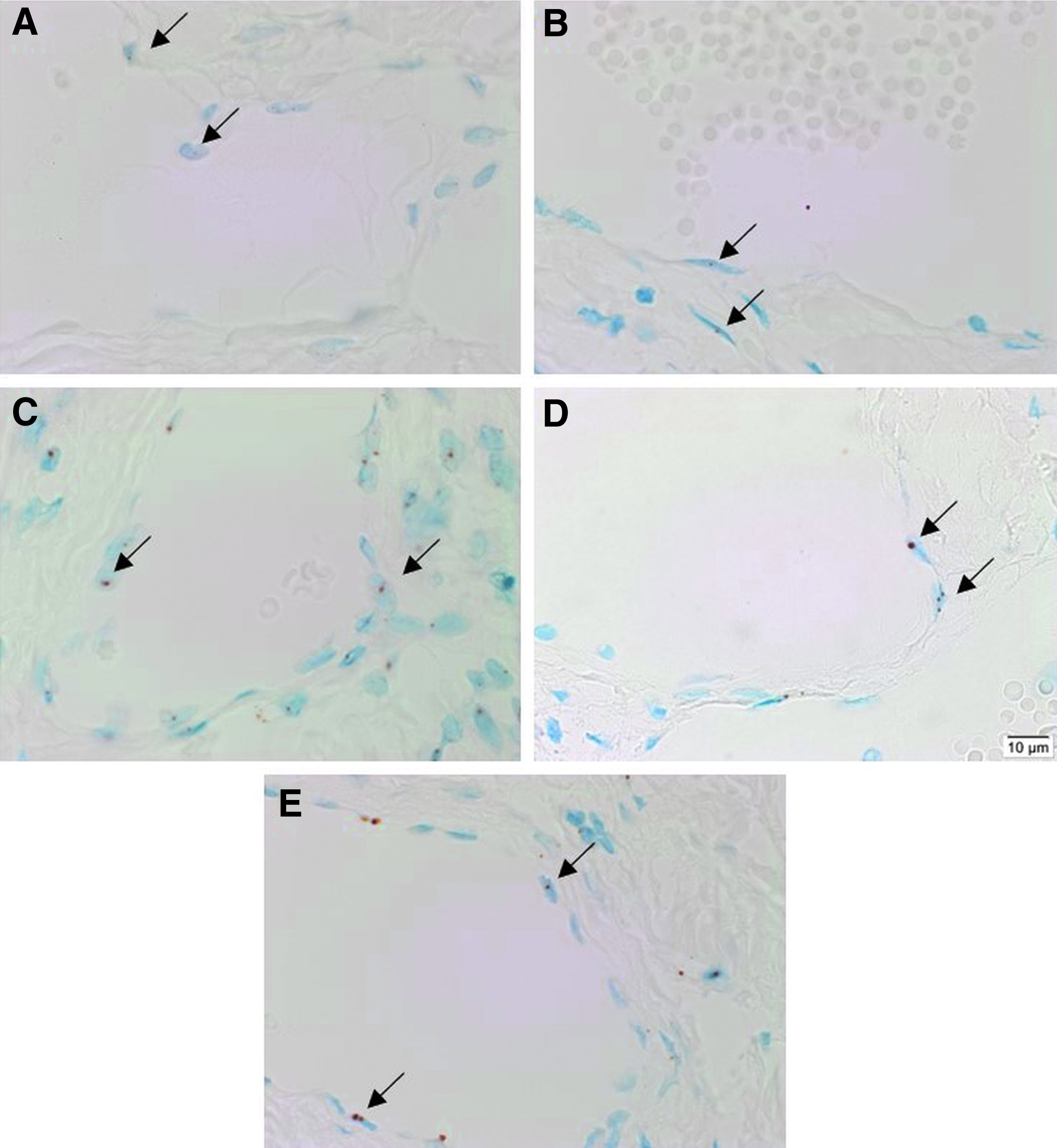
Representative sections of four microcystic lymphatic malformation tissue samples subjected to colorimetric in situ hybridization demonstrated the presence of mRNA transcripts for OCT4
Positive controls for OCT4 (Supplementary Fig. S3A, brown, arrows), SOX2 (Supplementary Fig. S3B, brown), NANOG (Supplementary Fig. S3C, pink, arrows), KLF4 (Supplementary Fig. S3D, brown), and c-MYC (Supplementary Fig. S3E, brown) on human seminoma, normal skin, seminoma, breast carcinoma, and normal prostatic tissue, respectively. The negative control was a sample of Bacillus (NM_L38424) (Supplementary Fig. S3F).
Reverse transcription quantitative polymerase chain reaction
RT-qPCR on the four snap-frozen mLM tissue samples demonstrated abundant expression of mRNA transcripts for OCT4, KLF4, SOX2, c-MYC, and NANOG, over the housekeeping gene GAPDH (Fig. 4). SOX2 was most abundantly expressed compared, in descending order, with KLF4 (p < 0.05) and c-MYC (p < 0.05), which had similar transcriptional levels, followed by NANOG (p < 0.01). OCT4 had the lowest expression. The expression of OCT4 was significantly different from that of SOX2 (p < 0.05), KLF4 (p < 0.05), and c-MYC (p < 0.05). The expression of NANOG was significantly different from that of KLF4 (p < 0.05).
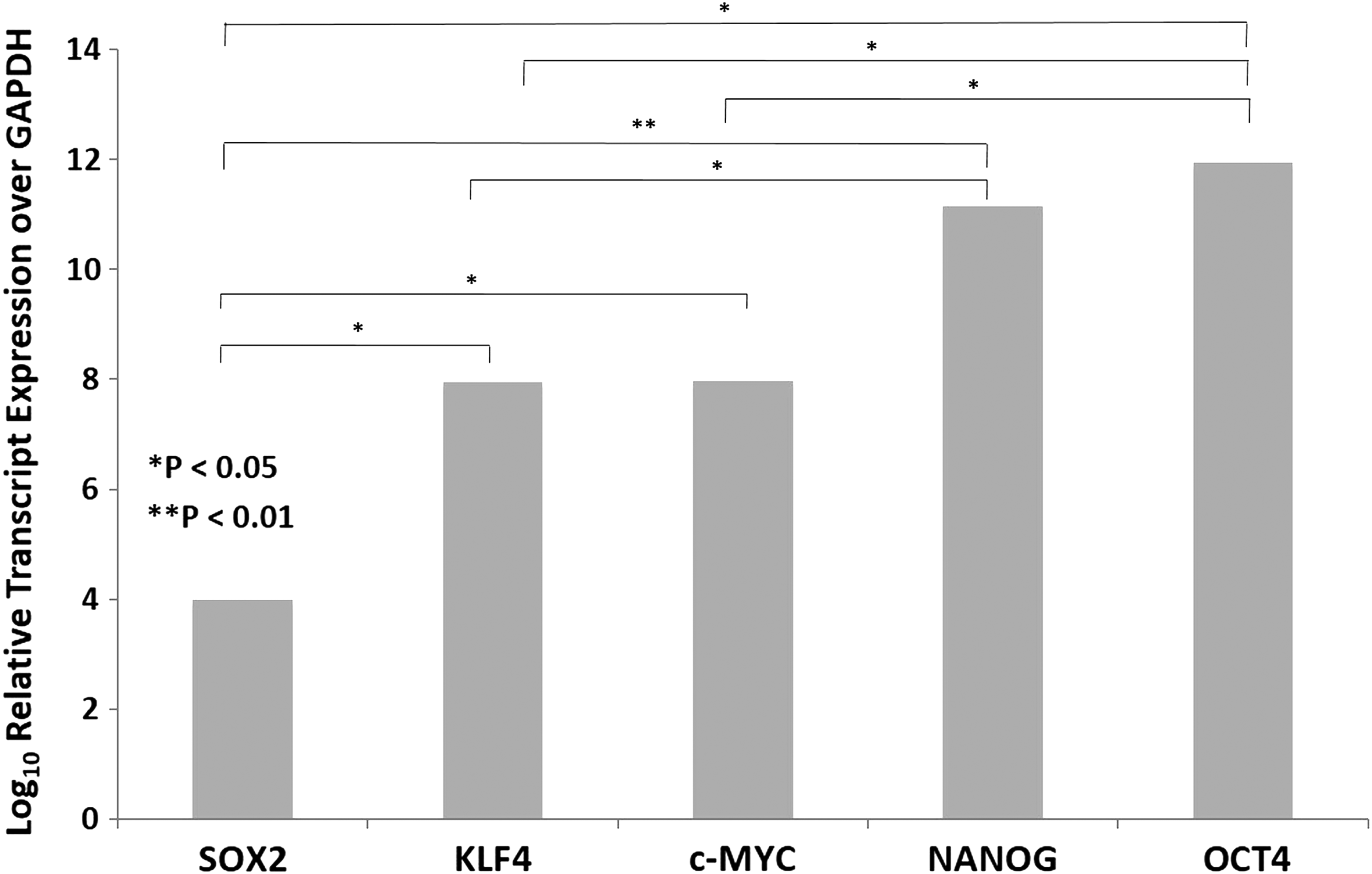
Graph showing average ΔCT values of reverse transcription quantitative polymerase chain reaction runs performed on four microcystic lymphatic malformation tissue samples. mRNA transcripts were amplified for SOX2, KLF4, c-MYC, NANOG, and OCT4. ΔCT was calculated by comparing cycle threshold values of these embryonic stem cell markers with housekeeping gene GADPH (*p < 0.05; **p < 0.01).
Conclusions
This report demonstrates the expression of ESC markers OCT4, SOX2, NANOG, KLF4, and c-MYC, markers involved in iPSCs,17,18 on the D2-40+ endothelium of the lesional vessels in mLM, with c-MYC and SOX2 also present within cells within the fibrous stroma. The results generated by IHC staining, RT-qPCR, and CISH showed similar expression patterns with SOX2 being most abundant, and OCT4 and NANOG having the lowest expression.
The ESC markers examined in this study were selected for their ability to confer and maintain stem cell properties.17,18 The expression of these markers has been well documented in a number of conditions, including cancer,20,21 Dupuytren's disease, 26 keloid disorder, 27 infantile hemangioma, 28 and VM. 14 These findings suggest the presence of a primitive population with capabilities of self-renewal and differentiation, driving these processes.
ESC markers have been extensively studied, particularly in cancer stem cells. The core nuclear transcription factors OCT4, SOX2, and NANOG work in concert to maintain the undifferentiated state. 29 KLF4 and c-MYC, with their respective roles in regulating cell cycle and cell proliferation, are also key mediators of stemness,16,17 with a recent report supporting the upregulation of c-MYC in mLM. 30 OCT4 has been labeled a rheostat protein, with genomic-wide effects critical for the reprogramming of somatic cells.19,31 NANOG is also associated with an early progenitor cell phenotype, whereas SOX2 is associated with a more differentiated state,16,20 therefore, it has been used as a progenitor cell marker in some studies. 20 Increased expression of these stem cell markers has been found to correlate with poor differentiation and recurrence after chemoradiotherapy in a number of cancer types.32–36 It has been proposed that cancer, analogous to normal tissue, is composed of a hierarchy of cells, with the ESC-like cancer stem cells giving rise to the downstream progenitor cells and differentiated cancer cells. 20
The current literature on LMs suggests that these lesions arise from aberrant lymphatic endothelial cells. 37 Wu et al. 7 have reported a population of LM cells expressing the progenitor cell marker CD133. These CD133+ cells demonstrate increased transcript expression of NANOG and OCT4 and reduced expression of lymphatic endothelial markers VE-cadherin, CD31, podoplanin, VEGFR-3, and Prox-1, compared with CD133− LM cells. These multipotent cells are located on the endothelium of the lesional vessels and within the stroma of LM. CD133 is a surface protein expressed by hematopoietic stem cells. Although its expression is associated with a number of malignancies, it is not essential for tumorigenesis, nor is it expressed on all cancer stem cells. Thus CD131 is considered a marker of progenitor cells rather than ESCs.16,20 In this report, we have shown the expression of the ESC markers OCT4, KLF4, and NANOG proteins predominantly located on the endothelium of the lesional vessels and transcriptional activation of these markers. There was greater expression of the progenitor-associated markers SOX2 and c-MYC on the endothelium of the lesional vessels and on the cells within the stroma. These findings suggest the presence of a primitive population on the endothelium of the lesional vessels and the surrounding stroma of mLM, the majority of which possess a progenitor phenotype with a small number of ESC-like cells.
Similar findings in VM 14 and those in mLM presented in this report suggest that this putative ESC-like population within both types of vascular malformations may be central to their development, although larger studies including in vitro and in vivo investigations are needed to confirm these results.
The novel finding of this study provides insights into the biology of this challenging clinical condition. Further elucidation of the mechanisms by which these cells maintain mLM is required to develop effective therapies.
Footnotes
Acknowledgment
We thank Ms. Liz Jones of the Gillies McIndoe Research Institute for her assistance in IHC and CISH staining.
Author Disclosure Statement
T.I. and S.T.T. are inventors of a provisional patent Treatment of Vascular Anomalies (PCT/NZ2017/050032), 2016. The authors otherwise declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest. This research did not receive any specific grant from funding agencies in the public, commercial, or not-for-profit sectors.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.