
Abstract
Background:
Lymphatic vessels drain fluids and solutes from interstitial spaces and serosal cavities. Among the solutes, low-density lipoproteins (LDL) are drained and can be detected in peripheral lymph, where they have been reported to exert a modulatory action on lymphatic vessels intrinsic contraction rate. In the present work, we investigated lymphatic vessel mechanical properties (contraction frequency and amplitude) that may be modulated by LDL application and the consequence on lymph flow.
Methods and Results:
Human-derived LDL were resuspended in phosphate-buffered saline (PBS) and microinjected in the interstitial space surrounding spontaneously contracting lymphatic vessels of the rat diaphragm, in vivo. Vessels' contraction rate and diameter were measured in control conditions (PBS) and after LDL injection. Lymph flow (Jlymph) was computed from contraction rate and diameter change. In some animals, after the recording procedure, diaphragmatic tissue samples were excised and immunostained with antilymphatic muscle (LM) actin to investigate the correlation between LM signal level and contraction amplitude. Data indicate a positive, saturating correlation between the abundance of LM actin and contraction amplitude, and LDL microinjection caused an acute increase in contraction frequency (+126%), a reduction of contraction amplitude to 75% of that obtained after PBS injection, and a +63% increase in Jlymph.
Conclusions:
From our in vivo analysis of the mechanical parameters affected by LDL, Jlymph was increased by a predominant effect on the contraction rate rather than amplitude, suggesting that the still elusive messaging system might be linked to the pacemaker sites.
Introduction
Lymphatic vessels drain extravasated fluid and macromolecules from interstitial spaces and/or serosal cavities, 1 eventually emptying into the venous blood, against a hydraulic pressure gradient. 2 This efficient lymph recirculating system is made possible by the presence of an extrinsic and an intrinsic propulsive mechanism. The first is given by the contractions of skeletal muscle fibers and by the mechanical forces that surrounding tissues might exert upon lymphatic vessels,3–5 while the latter relies on the presence of lymphatic muscle (LM) cells in the wall of collecting lymphatic vessels, which are endowed with spontaneous, rhythmic contractions,6–9 and can be influenced by intraluminal pressure,10,11 chemical mediators, 12 and the surrounding environment, such as temperature and fluid osmolarity.13,14
Low-density lipoproteins (LDL) are one of the five groups of lipoproteins in aqueous media such as plasma, extracellular fluid, and lymph. 15 Accumulation of LDL in the artery wall is supposed to be responsible for atherogenesis. In this respect, several evidences in literature are emerging that link the lymphatic system to the removal of cholesterol and LDL from the artery wall, thus preventing or limiting the atherogenic process.16–19 More generally, lymphatic vessels can drain extravasated lipoproteins back into the blood stream.20–24
Since LDL receptors are expressed on the surface of lymphatic endothelial cells and muscle of lymphatic collectors, 25 and lymph flow is increased after lipid absorption;26,27 in this work, we wanted to assess in vivo the effect of the acute application of plasma-derived LDL on the mechanics of lymph propulsion in diaphragmatic collecting lymphatic vessels.
Materials and Methods
In vivo experiments
Experiments involving animals were reviewed and approved by the Ethics Committee of the University of Insubria (OpBA) and the Italian Ministry of Health in accordance of the Italian D.Lgs 26/2014. Experiments were performed on 11 adult Wistar rats (body weight: 319 ± 44 g), anesthetized by an intraperitoneal injection of ketamine (75 mg/kg, Imalgene 1000; Merial Italia, Italy) and medetomidine (0.5 mg/kg, Domitor; Pfizer, Italy) cocktail in saline solution; and half boluses were administered every hour until the end of experiments, while continuously checking the appropriate level of deep anesthesia.
After a subdiaphragmatic intraperitoneal injection of 0.8 mL of 2% fluorescein isothiocyanate (FITC)-dextrans (FD250S; Sigma-Aldrich, Italy) in saline solution, animals were let to breathe spontaneously for 60 minutes on a heated (37°C) pad to let lymphatic vessels drain the injected solution and become filled with FITC-dextran solution. Animals were then tracheotomized, paralyzed with a 0.3 mL bolus of 2 mg/mL pancuronium bromide (P1918; Sigma Aldrich) in saline, and mechanically ventilated (Inspira model; Harvard Apparatus) at tidal volume and respiratory rate automatically set by the instrument based on body weight. Then, after midsternal thoracotomy, diaphragmatic lymphatics were visualized under a LED epi-illuminated SV11 (Ex/Em: 490/520; Zeiss, Italy) stereomicroscope, being the diaphragmatic surface frequently rinsed with warmed saline to avoid tissue dehydration.
Spontaneous contractions of lymphatic vessels, often located at the muscle periphery and organized in loops, were video recorded at 1–5 Hz by a CCD camera (Orca ER; Hamamatsu, Italy) connected to a personal computer running SimplePCI software (Hamamatsu). The best candidate contracting sites, to be tested for LDL effect, were evaluated by the video data STD (standard deviation) analysis, based on a previous data set (n = 5 rats) specifically performed and analyzed (see Results, first section).
Injecting micropipettes were pulled from borosilicate glass capillaries (1B100-4, outer diameter 1.0 mm, and inner diameter 0.78 mm; WPI Europe, Germany) to tip diameters of ∼25 μm using a P-97 Flaming/Brown type Micropipette Puller (Sutter Instrument), and beveled at 30° to reduce indentation and tissue deformation during the injection maneuver into the diaphragmatic interstitium. The interstitial injection of 9.2 nL boluses of phosphate-buffered saline (PBS) alone or a solution of 0.25 μg/μL of LDL in PBS was performed using a microinjector (Nanoliter 2000; World Precision Instruments, Germany) set at an injecting rate of 10 nL/s and driven by a mechanical micromanipulator.
For each animal, at least 2–3 spontaneously contracting sites located in different and separated lymphatic vessels belonging to both hemidiaphragms could be detected. Therefore, for each animal it had been possible to perform 4–5 microinjections of either PBS (control injection) or LDL solution (test injection). To avoid possible effects due to subsequent microinjections in the interstitial space close to one single site, only one microinjection was performed at each site. PBS microinjection was also performed to assess that no artifacts could be attributed to the microinjection itself and to account for the effect of temperature on the LDL-dependent effect, since temperature was found to be a key factor affecting lymphatic contractility. 14 Indeed, at the time of injection, the measured temperature of the injected solutions (both LDL and PBS) was about 25°C, lower than the measured average 32°C of the exposed diaphragmatic surface in our in vivo experiments.
Each contracting site was thus recorded for 1 minute before the microinjection of either PBS or LDL, and then subsequently recorded to document the eventual effect of PBS or LDL.
At the end of the procedure, animals were euthanized with an anesthesia overdose.
LDL purification from plasma
Human native LDL purified from plasma of normolipidemic, healthy volunteers of both sexes were kindly provided by the “Ospedale di Circolo e Fondazione Macchi.” After isolation by means of ultracentrifugation as in Viola et al., 28 LDL were desalted and freed of ethylenediaminetetraacetic acid (EDTA) by filtration in PD-10 desalting columns (17085101; GE Healthcare, Italy) following manufacturer's instructions, and concentrated with Centriprep concentrators (Centriprep YM-10,4304; Merck, Italy). Moreover, absence of significant amounts of EDTA in the desalted LDL preparation was confirmed by testing parallel aliquots of the same LDL batch used for the experiments with oxidation using CuSO4 and evaluating the time-course of the process at 234 nm. After sterilization, performed by filtration through 0.2 μm filters, protein content was assessed by Bradford assay, whereas total cholesterol was measured by a colorimetric enzymatic assay (Olympus Cholesterol OSR6516). Then, LDL were diluted in PBS with calcium and magnesium (14040-091; Gibco, Italy) at a final concentration of 0.25 μg/μL, a concentration comparable to the one of LDL in rat serum. 29
LDL conservation and microinjection in the in vivo experiments
Since purified and desalted LDL are more prone to oxidation, with a literature-reported lag-time of about 60 minutes at worst, 30 LDL solution to be injected was freshly prepared from donor plasma the day before each experiment and stored at 4°C until used to preserve as much as possible its not-oxidized state.
On the day of the experiments, each aliquot of LDL was kept on ice until loaded into the micropipette tip just before the microinjection. From bolus microinjection into the interstitial space to the onset of LDL effect, an average 84.6 seconds elapsed. This time is much less than the worst lag time for oxidation of 60 minutes reported in literature, so that it was reasonable to assume that LDL were still in their not-oxidized state when the effect was evident at the lymphatic vessel level.
Whole mount assay
Immediately after animal suppression, specimens of diaphragmatic tissues containing the lymphatic vessels in vivo tested and video recorded were excised and fixed with 4% paraformaldehyde (158127; Sigma Aldrich) in PBS at room temperature for 45 minutes, washed three times in PBS, and then permeabilized with ice cold methanol and 0.5% Triton X-100 (Sigma Aldrich). Specimens were then incubated with blocking solution (1% bovine serum albumine, 5% goat serum in PBS) for 1 hour at room temperature followed by an overnight incubation with primary mouse anti-alpha smooth muscle actin antibody (NB600-536; Novus Biologicals) at 4°C. After three washes in PBS, samples were incubated with Alexa Fluor® 647 secondary goat anti-mouse antibody (1:100 goat anti-mouse IgG2a, Molecular Probes) for 2 hours at room temperature. Finally, after three rinses in PBS, specimens were mounted onto glass coverslips with FluoroShield with 4′,6-diamidino-2-phenylindole (F6057; Sigma-Aldrich), to be analyzed in their whole thickness with a confocal microscope (TCS SP5; Leica) with a 20 × dry objective and 0.5-μm z-axis steps. Maximum projections of adjacent fields were color-merged with ImageJ Software (https://imagej.nih.gov/ij/) and manually aligned in a continuous panorama using Adobe Photoshop Software.
Correlation between best contracting sites and LM density
In five rats, video data of spontaneously contracting lymphatic vessels were stabilized by the “Align slices in stack” ImageJ plugin and analyzed by the “Standard Deviation” Z-Project function, 31 resulting in black and white images (STD images) where the thickness of the white vessel edges represents the standard deviation of the location of the vessel wall during spontaneous contractions. Thus, the higher the white signal, the larger the standard deviation and hence vessel constriction for each contraction.
After performing whole mount (WM) assay, WM panoramas were color split by using ImageJ Software, to obtain 8 bit b/w images (WM images) of the LM mesh surrounding the lymphatic vessels wall, then regions of interest (ROIs) of about 100 μm of length were manually tracked for the full length of the lymphatic network, and for each ROI, the mean gray value (mean pixel intensity of the ROI) was measured, subtracting the mean gray value of the surrounding background. The same ROIs were manually placed in the corresponding STD images and again mean gray values were measured to be matched to corresponding ones from WM images. For each couple of images, data were plotted as gray WM values (X) versus gray STD values (Y) and grouped according to the X values to compute X data points at regular intervals of about 10 values of pixel intensity. For each sample, data were fitted by a three parameters single exponential rise to maximum equation of the form below:
where y0 is the STD lowest value (Y value when X = 0), a is the upper value and b is the increase rate of the WM/STD values ratio.
For each experiment, STD and WM mean values were normalized to their respective maximal value (from 0 to 100) and plotted in a normalized STD gray value versus normalized WM gray value graph. All data points (n = 42) were then fitted by the two parameters single exponential rise to maximum equation
where a is the upper value and b is the increase rate of the WM/STD ratio.
Data analysis
Diameter over time profiles of contracting lymphatic vessels were obtained from in vivo recorded data using “Diameter” plugin 32 of ImageJ software. Mean diastolic (dD, μm) and systolic (dS, μm) diameters were obtained from at least four contraction cycles of plotted profiles, and for each stroke, contraction amplitude (Δd, μm) was computed as Δd = dD − dS.
Diameter traces were analyzed by the “Autocorrelation” function of Clampfit 10 Software (Molecular Devices) to measure the contraction frequency (fc, cycles/min) and also the contraction time (tc, seconds, the time elapsed from the beginning of the contraction to the attainment of the smallest diameter as measured in diameter-time plots) was computed.
For each stroke, contraction velocity (vc, μm/s) was computed as the ratio between contraction amplitude and the corresponding contraction time.
Since submesothelial diaphragmatic lymphatics are ellipse-shaped
33
and the systolic to diastolic radius ratio is 0.35,
34
the stroke volume (SV, pL) was computed as
in a lymphatic vessel tract of 105.50 μm in length,
14
being rD and rS diastolic and systolic radii, respectively, and 106 the μm3 to pL conversion factor. From SV data, lymph flow (Jlymph, nL/min) was computed as
Statistical analysis
Data are presented as mean ± standard error. Statistical analysis (paired and/or unpaired Student's t-test and analysis of variance) was performed with SigmaPlot 10 Software (Systat Software) after data normality check and statistical significance was set at p < 0.05.
Results
Correlation between extent of vessel contraction and lymphatic smooth muscle abundance
To maximize the eventual effect of LDL application, we sought to find a method to quickly identify the sites were LM fibers could be more abundant in the lymphatic loop under investigation. To this aim, in five animals, we performed a careful analysis of the correlation between the abundance of LM fibers, witnessed by the signal level of the α-smooth muscle actin staining in WMs and the STD signal measured from the video recordings of the lymphatic network under investigation (see Materials and Methods section for details). Starting from 60 seconds video recordings of the spontaneous activity of a lymphatic loop, as in the example frame in Figure 1A, in few minutes, it was possible to obtain the STD analysis of the whole video (Fig. 1B, where the boxes d′ and e′ correspond to vessel segments marked d and e in Fig. 1A) and identify zones of higher gray values even by eye (box e′ in Fig. 1B). After the WM processing and imaging, from images such as the one in Figure 1C, it was then possible to measure the mean signal of α-smooth muscle actin staining (red signal) in the wall of the FITC-filled (green signal) lymphatic vessels. Figure 1D shows the plot of the relationship between WM signal and STD value of this specific example. Applied on all available samples, the analysis showed that the overall relationship (Fig. 2) between STD and WM mean values, expressed as percentage of their maximal WM and STD values, respectively, could be fitted with a single exponential curve (dashed line, see Materials and Methods section) with a rate constant of 0.043 ± 0.004 WM/STD (n = 42, r2 = 0.95).
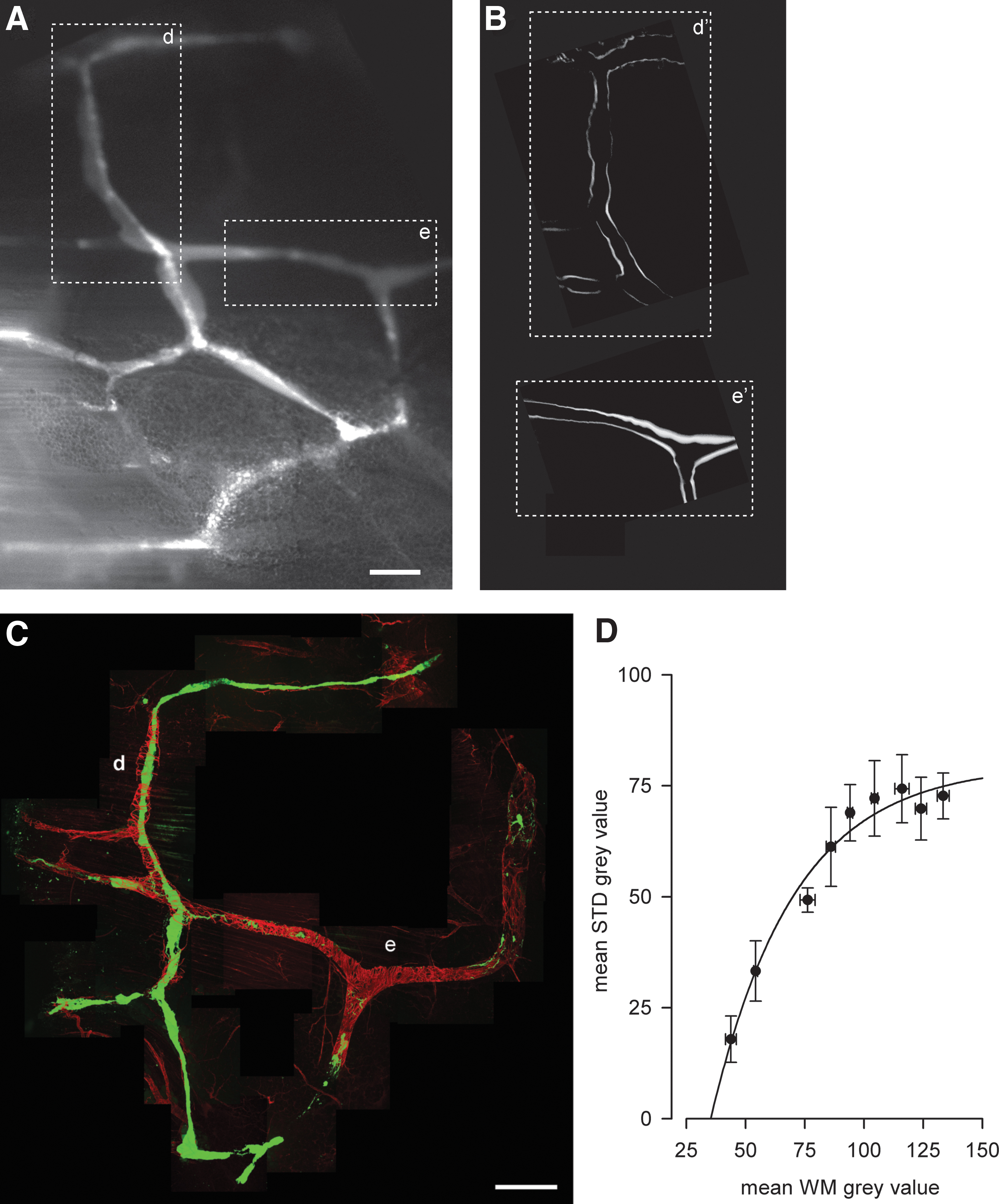
Overall analysis of all normalized STD versus WM data point highlighting that the higher STD gray value (due to thicker vessels edges from STD images) correlated to a much more density in LM (higher WM α-SMC actin signal) surrounding spontaneous intrinsically contracting vessels. r2 = 0.95.
Based on these analyses, in the following experiments, we injected the LDL solution at sites of the interstitial space surrounding the spontaneously contracting loop where the normalized STD signal was above 90%, confident that those segments were also endowed with an extensive mesh of LM around the vessel wall (normalized WM value >60%).
Effect of LDL application on contracting and not contracting vessel tracts
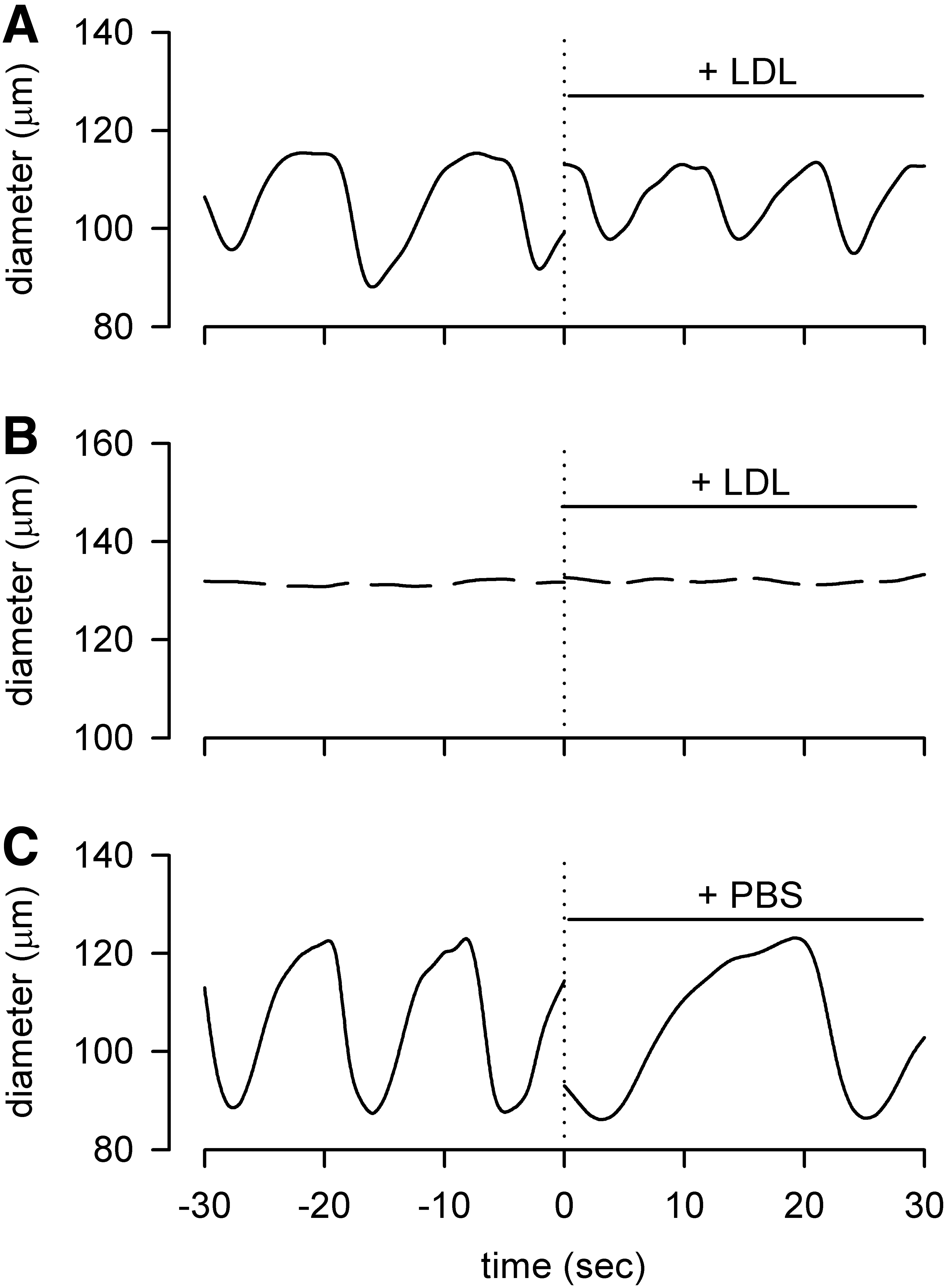
Microinjection of LDL solution in the interstitial space surrounding actively contracting lymphatic vessel tracts gave rise to a variation in both fc and contraction amplitude (Fig. 3A), whereas when LDL solution was microinjected in the interstitial space next to the vessel wall of a not contracting site, no change in vessel diameter was ever observed (Fig. 3B). The actual effect of LDL on vessel contractility was likely even more accentuated than that shown in Figure 3. Indeed, since the LDL solution was kept on ice and front-filled in the injecting micropipette 1–2 minutes before injection, it is reasonable to assume its temperature to be colder than the one of the exposed diaphragmatic surface (see Materials and Methods section). In fact, as expected from our previous study, 14 when PBS alone (kept on ice and prepared for the injection following the same method of LDL solution) was microinjected as a control, fc immediately decreased (Fig. 3C) as a result of the temperature drop in the interstitial space surrounding a spontaneously contracting vessel.
The analysis of mean fc values (Fig. 4A) showed that, starting from a control value of 4.7 ± 0.4 cycles/min, LDL application increased lymphatic intrinsic fc to 6.1 ± 0.7 cycles/min (*p < 0.05, n = 8 vessels, paired t-test), while PBS injection lowered fc from 4.3 ± 0.5 cycles/min to 2.7 ± 0.6 cycles/min (*p < 0.05, n = 6 vessels, paired t-test). However, if we account for the temperature-driven effect and compare the effect of LDL injection to the one of PBS injection, the difference is even more accentuated (6.1 ± 0.7 cycles/min in LDL versus 2.7 ± 0.6 cycles/min in PBS, ##p < 0.01, n = 14 total vessels, unpaired t-test). Further analysis of contraction time (tc, Fig. 4B) showed that LDL application not only exerted its effect at the pacemaker level, but it also lowered the tc. Indeed, from a mean value of 4.1 ± 0.3 seconds in control, LDL application shortened tc to 3.2 ± 0.2 seconds (**p < 0.01, n = 8 vessels, paired t-test), while PBS application had no significant effect on this parameter (4.3 ± 0.4 seconds in control vs. 4.1 ± 0.3 seconds in PBS, n = 6 vessels). Hence, also direct comparison of LDL application versus PBS showed a significant difference with respect to tc (#p < 0.05, n = 14 total vessels, unpaired t-test).
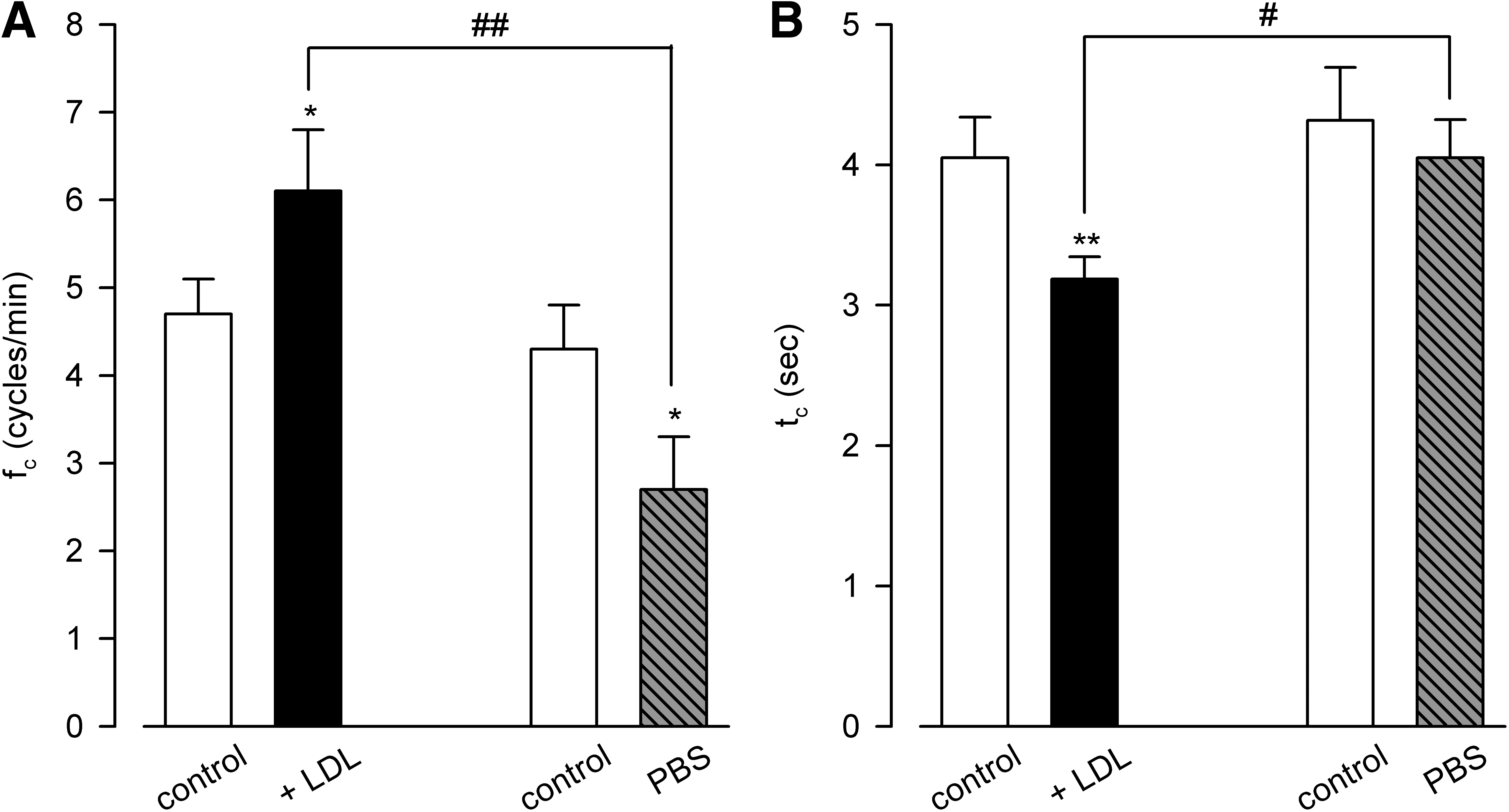
Figure 5A shows that LDL application significantly lowered contraction amplitude (40.11 ± 4.21 μm in control vs. 30.56 ± 4.25 μm in LDL, *p < 0.05, n = 8 vessels, paired t-test), whereas PBS application did not exert any significant effect on this parameter (38.53 ± 8.75 μm in control vs. 40.96 ± 7.84 μm in PBS, p = 0.35, n = 6 vessels, paired t-test). No significant effect of LDL or PBS was present on the resting, diastolic tone of the LM, since no statistically significant variation was induced by the application of either LDL or PBS (Fig. 5B) in the interstitial spaces around contracting and not contracting (n = 4 vessels) lymphatic vessels.

Upon LDL application, computed Jlymph (1.0 3 ± 0.18 nL/min, Fig. 6) did not vary significantly (p = 0.81 n = 8 vessels, paired t-test) with respect to control (1.05 ± 0.13 nL/min). However, since PBS application induced a statistically significant decrease in Jlymph (from 1.0 ± 0.2 nL/min in control to 0.63 ± 0.16 nL/min in PBS, *p < 0.05, n = 6 vessels, paired t-test) likely due to temperature drop, the difference between mean Jlymph values in LDL versus PBS became statistically significant (1.03 ± 0.18 nL/min for LDL application vs. 0.63 ± 0.16 nL/min for PBS application, #p < 0.05, n = 14 total vessels, unpaired t-test), with an average Jlymph increase of +63.7% ± 18.0% with respect to the one obtained following PBS application alone (p < 0.01, one sample t-test n = 8 vessels).
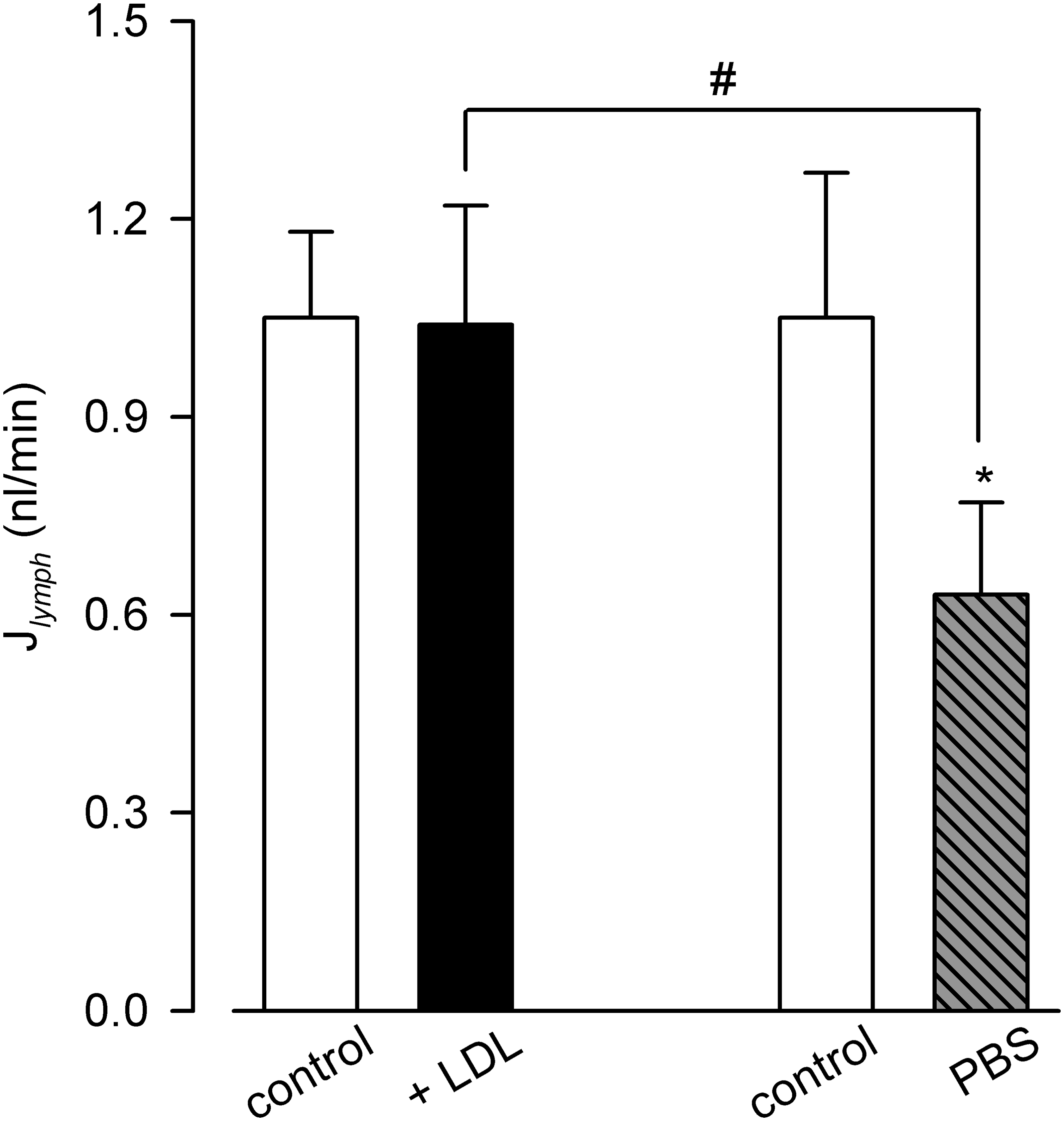
Lymph flow (J ymph ) computed in spontaneously contracting lymphatics in preinjection control conditions (white bars, not significantly different for the two data set) and after interstitial LDL (black bar) or PBS-only (gray striped bar) injections. PBS induced a Jlymph reduction (*p < 0.05), attributable to the decrease in temperature due to the saline injection. Comparing LDL to PBS effect on lymph flow resulted in a significant increase in Jlymph (#p < 0.05) of about +64%. *paired t-test. #unpaired t-test.
Conclusions
Relationship between STD and WM signal levels
The exponential relationship between STD and WM normalized values, found to develop a rapid method to assess the abundance of LM around a vessel tract so to direct the LDL injection in its proximity, per se shows two distinctive features of the spontaneous contractions: first, the saturation of STD value for increasing WM signal level implies that there is a finite limit of contraction amplitude, which does not correspond to complete occlusion of the lymphatic vessel during systole, and that the abundance of LM in the vessel wall, after a certain level, is not determinant for the contraction amplitude.
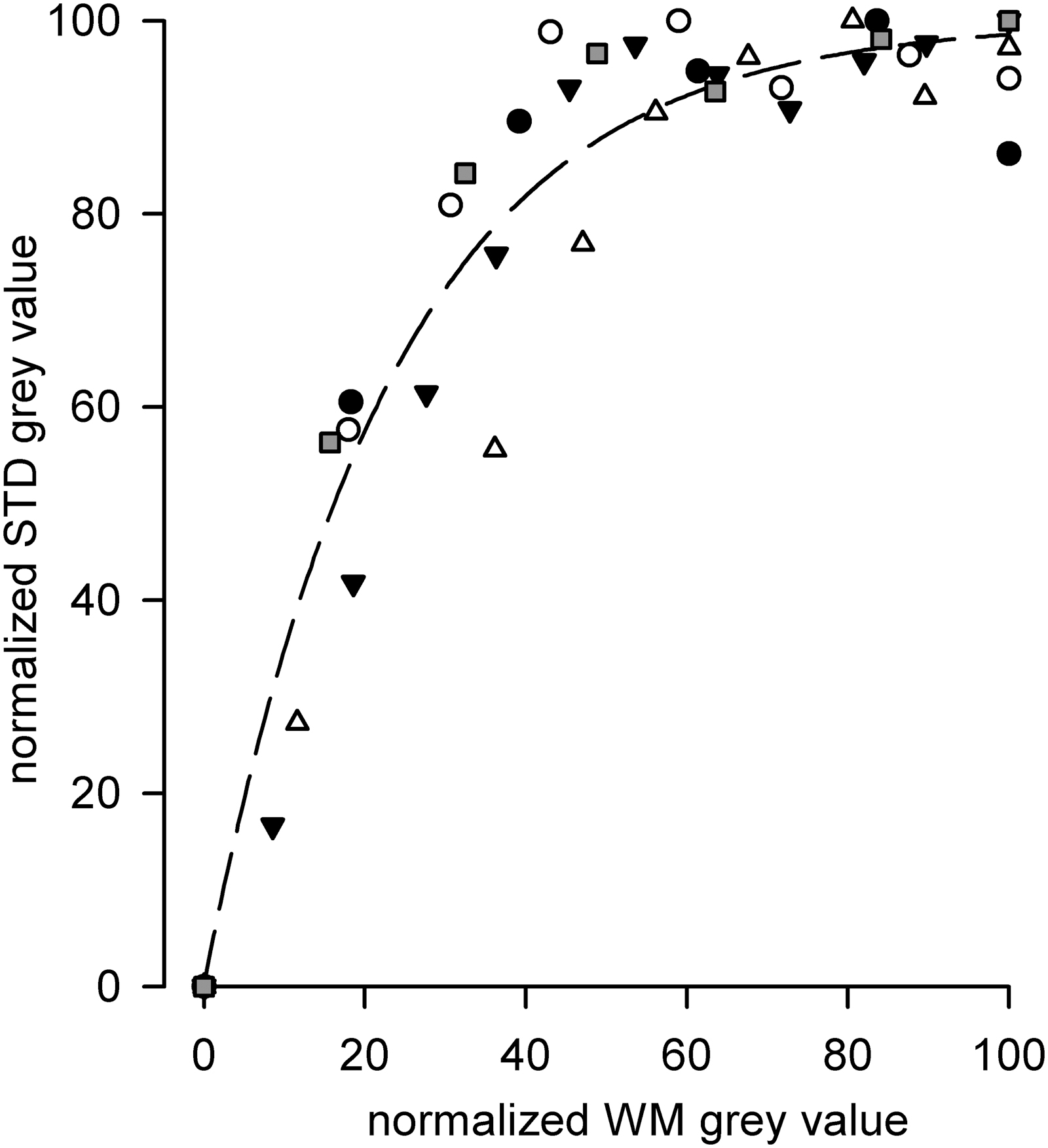
Second, from the plot of Figure 7, where the five individual STD/WM relationships analyzed were individually plotted, it is to note that a small STD value can be present also for WM signal levels in the middle range, thus confirming our previous findings 33 that LM is present also in the walls of lymphatic vessels which do not seem to contract spontaneously. Therefore, an actual contracting behavior might be the result of a proper, functional arrangement of the muscle around the lymphatic vessels and/or might be due to the presence of a pacemaker site functionally joined to the LM layer.12,35,36

Plot of STD versus WM data points from five experiments to correlate the STD gray value (representing the intrinsic active contraction of lymphatic vessels) to the LM extent (WM gray value) in the wall of spontaneous contracting vessels. Circles, squares and triangles identify data coming from each single vessel.
Net effect of LDL on spontaneous contractions and Jlymph
Due to the technical challenges of the in vivo LDL injection in the diaphragmatic tissues and the simultaneous need to maintain the LDL solution to be injected ice-cold until the injection time to preserve the native state of LDL, the effect of LDL solution injection on lymphatic function had to be adjusted for the effect of temperature, since we recently showed that lymphatic spontaneous activity is variable upon this physical parameter. Therefore, we chose to use ice-cold PBS (the solvent of LDL solution) as an appropriate control, keeping the same timing between LDL and PBS injections.
Hence, the fc recorded in control conditions (white bars of Fig. 4A) is lower than the one published in our previous article, 14 which had been recorded in an ex vivo model kept at a controlled temperature of 37°C, but it is coherent with the sigmoidal response to temperature described therein. More interestingly, however, the response of lymphatic vessels to the injection of cold PBS alone was, as expected, a lower, temperature-dependent fc (gray-striped bar, Fig. 4A). Therefore, to assess the net effect of LDL injection devoid of any temperature-driven component, the proper comparison to be made is between LDL and (cold) PBS application.
Overall, from data presented in Figure 4, LDL application increases fc with respect to PBS and shortens contraction time (tc). Moreover, the concomitant reduction in contraction amplitude (Fig. 5A) due to LDL causes the contraction rate to remain substantially the same between PBS and LDL application (10.24 ± 1.15 μm/min in PBS vs. 9.90 ± 1.18 μm/min in LDL, paired t-test p = 0.8675 n = 14 total vessels). These data, together with the fact that LDL did not modify the resting, diastolic tone of lymphatic musculature (Fig. 5B), seem to point in the direction of an LDL effect on the phasic, but not tonic, control of LM spontaneous contractions.
This phasic control is exerted at the pacemaker site (increase in fc) and might also be due to a modulatory action at the contraction site (decrease of contraction amplitude due to the decrease in contraction time). However, at difference to what can be expected for the cardiac sinus pacemaker, the increase in fc seems to reflect a shortening of the systolic, rather than the diastolic, phase.
This, in turn, raises the question whether Jlymph could be affected by LDL application, since the opposite variations of fc and contraction amplitude (and hence SV) might compensate for each other. Indeed, in the presence of LDL computed Jlymph significantly increased compared to what observed following PBS injection, likely reflecting the prevalence of fc elevation following LDL injection (Fig. 6).
The in vivo response of collecting lymphatic vessels to the interstitial injection of LDL became evident in about 90 seconds, although the experimental complexity during the whole procedure might have prevented a more accurate measurement of the exact time lag between microinjection and onset of the effect. Moreover, to properly dissect the intracellular pathway linking LDL exposure to the modification of lymphatic mechanics, a more controlled experimental environment is needed, such as ex vivo experiments on genetically modified animal models lacking LDL receptors or other modulators of LDL receptor-mediated response (Pcsk9−/− and/or ldlr−/− mice), which are currently under technical evaluation but are, however, beyond the scope of the present work.
Pathophysiological considerations on the LDL effect on lymphatic vessels intrinsic contractions rate
An actual determination of LDL concentration in the lymph of diaphragmatic lymphatic vessels has never been obtained by our group despite several attempts in the past years due to the technically challenging environment of the in vivo procedures that enabled us, in the best trials conducted so far, to collect no more than 200 nL of lymph; nor are we aware of any determination made by other authors in literature in this peculiar lymphatic district.
Diaphragmatic lymphatic vessels like the ones of the present work are superficial and located in the pleural surface of the diaphragm, 37 where they are exposed not only to the LDL content of the interstitial fluid surrounding them, coming from plasma-extravasated LDL, but also to the LDL content of pleural fluid, 38 drained into them by means of the pleural lymphatic stomata.
Extravasated LDL can use lymphatic vessels to return to blood flow, 20 although LDL concentration in the interstitial fluid and peripheral lymph is supposed to be lower than plasma due to LDL clearance during the interstitial space transit.39,40
In this respect, the LDL concentration used (0.25 μg/μL) was about 50% of that of rat plasma and thus higher than the 14% of plasma LDL concentration estimated to be present in the peripheral lymph of rabbits. 41
LDL are present in pleural fluid, and their concentrations varies, although not in a strict manner, linearly with serum LDL concentration. The LDL concentration used in this work is almost identical to the one of pleural fluid (0.27 ± 0.02 μg/μL) of a human having a normal (1.25 ± 0.04 μg/μL) serum LDL concentration. 42
Taken together, these data suggest that, in our in vivo preparation, diaphragmatic lymphatic vessels could have been challenged with a normal-to-high LDL concentration. However, since pleural and interstitial fluid LDL concentrations vary with the serum concentration, it is unlikely that in pathophysiological conditions, an abrupt increase of interstitial and/or pleural fluid LDL concentration could happen with the timing used in our experiments (less than one second for the complete delivery of the LDL solution bolus in proximity of the lymphatic vessel wall, see Materials and Methods section). Pairwise, since the need for restoration of proper ventilation to the animal and the need to avoid repeated microinjections at the same site prevented an observation of the LDL effect for periods longer than few minutes, it was not possible to ascertain whether a prolonged exposition to LDL might have led to eventual adaptation phenomena resulting in changes of the measured, acute effect that we have documented.
Indeed, the primary goal of the experimental design was to obtain a clear effect, if present, of LDL application on the lymphatic vessel, and thus LDL concentration and microinjection timing and procedure were optimized to deliver a steep increase of LDL concentration at the vessel wall, confident that, given the limitations that the in vivo system poses on spotting subtle differences, it would have been impossible to successfully record and analyze slow-developing changes that might well be present in pathophysiological conditions but develop in time frames of hours or more.
Overall, the present in vivo results might help to clarify the adaptation of lymphatic intrinsic pumping that is activated following an acute exposure to LDL, which seems to be due to a prevalent effect on fc together with a decrease in contraction amplitude that does not involve a change in the diastolic, basal tone of LM. These effects, in turn, gave rise to a net increase in Jlymph.
This latter phenomenon has already been assessed in experimental models where LDL and/or LDL receptor homeostasis and function have been altered 25 ; and in this context our in vivo data represent, to our knowledge, the first mechanical in vivo description of how this increase in lymph flow could be attained by an acute exposure of lymphatic vessels to LDLs.
Footnotes
Author Disclosure Statement
No competing financial interests exist.
Funding Information
E.S. holds a postdoc position granted to A.M. and funded by University of Insubria. B.B. holds a postdoc position funded by University of Insubria.