
Abstract
Background:
The lymphovenous junction (LVJ) of the thoracic duct (TD) is the principle outlet of the lymphatic system. Interest in this junction is growing as its role in lymphatic outflow obstruction is being realized, and as minimally invasive procedures for accessing the terminal TD become more common. Despite the growing clinical significance of the LVJ, its precise form and function remain unclear. The aim of this article was to systematically review the literature surrounding the structure and function of the LVJ and its associated lymphovenous valve (LVV).
Methods and Results:
A systematic review of the structure and function of the LVJ and LVV was undertaken using the MEDLINE, Scopus, and Google Scholar databases. Human and animal studies up to November 2019, with no language or past date restriction, were included. Forty-six relevant articles were reviewed. The LVJ shows marked anatomical variation. A valve is frequently absent at the LVJ, but when present it displays numerous distinct morphologies. These include bicuspid semilunar, ostial, and flap-like structure. Other factors, such as functional platelet plugs, or the tangential/intramural course of the terminal TD across the vein wall, may work to prevent blood from entering the lymphatic system.
Conclusions:
The form and function of the LVJ remain unclear. Dedicated studies of this area in vivo are required to elucidate how this part of the body functions in both health and disease.
Introduction
The thoracic duct (TD) is the principal vessel of the lymphatic system. It drains the vast majority of the body's lymph back to the blood vasculature through a lymphovenous junction (LVJ), typically located at the confluence of the left internal jugular and subclavian veins (the left venous angle) 1 (for simplicity “LVJ” is used to refer to the lymphovenous junction associated with the TD). Approximately 3 L of lymph (including chyle: a lipid-rich emulsion absorbed by the intestines) crosses the LVJ per day. 2 This fluid contains protein, dietary lipid, metabolites, electrolytes, vitamins, immunoglobulins, and lymphocytes.3,4
Bicuspid semilunar valves are distributed along the TD lumen, preventing retrograde lymph flow.5,6 A lymphovenous valve (LVV) is thought to guard the mammalian LVJ by regulating the flow of lymph into the venous circulation and preventing blood from entering the lymphatic system.2,7 Previous descriptions of this valve are often conflicting, and its precise form and function remain unclear in humans.
It is known that ∼80% of TD lymph is derived from the abdominal viscera. 8 Supported by an increasing body of evidence, the “gut–lymph” concept 9 states that lymph draining the gut in acute and critical illness undergoes a profound compositional change. The altered and toxic gut–lymph is discharged through the LVJ into the blood circulation where it promotes systemic inflammation and multiple organ dysfunction/failure,10–12 which is a leading cause of mortality in intensive care units worldwide. 13
Furthermore, impairment of lymph flow at the LVJ (lymphatic outflow dysfunction) can cause edema of upstream tissues, including vital organs. This may occur in conditions of raised central venous pressure, such as right heart failure or aggressive fluid resuscitation, where the pressure gradient becomes too high for lymph to flow into the central veins.14–16
The LVJ has, therefore, become a focus for potential clinical intervention in critical illness 17 and conditions associated with chronic venous hypertension (e.g., right heart failure and liver cirrhosis). 8 Detailed knowledge of the LVJ and LVV is required to help advance these areas. The aim of this study was to systematically review the structure and function of the LVJ and LVV.
Materials and Methods
A systematic review of the scholarly literature surrounding the structure and function of the LVJ and LVV was performed. The MEDLINE (Ovid), Scopus, and Google Scholar databases were searched to identify the relevant literature. Publications up until November 2019 were included; no past date limit was imposed. The search terms used were the combined results of “thoracic duct” OR “ductus thoracicus” OR “lymphatic duct” OR “lymphovenous” AND the combined results of “anatomy” OR “morphology” OR “structure” OR “variation” OR “course” OR “imaging” OR “physiology” OR “function” OR “histology” OR “termination” OR “distal” OR “cervical” OR “neck” OR “ostium” OR “opening” OR “outlet” OR “drainage” OR “valve.” A manual search of the primary records' citations was performed to identify any relevant literature not found in the initial database searches. Relevant pre-18th century anatomical descriptions were found through manual archival searches.
Inclusion and exclusion criteria
Cadaveric, surgical, and medical imaging studies that described the structure and function of the LVJ and/or LVV were reviewed. Non-English studies were included where available. Both human and mammalian animal studies were included. Studies were excluded if they solely examined TD pathology, TD interventions, or TD structure or function without describing the LVJ/LVV.
Data extraction
Qualitative data of the LVJ/LVV and quantitative data surrounding the occurrence of a valve at the LVJ were extracted. Data surrounding the function/mechanism of the LVV based on empirical observation were also recorded.
Results
A total of 8716 records were identified through the database searches. After removing duplicates, 5405 records were screened based on their title and abstract. Of these, 111 articles were identified as appropriate for full-text review. Ultimately, 40 articles met the inclusion criteria. A further six articles were identified through manual searches. In total there were 46 articles included in this review (Fig. 1).

Preferred reporting items for systematic reviews and meta-analyses flow chart of literature search strategy.
Structural Anatomy
The following refers to human studies except where stated otherwise.
Terminal TD variation
The branching pattern of the terminal TD and location of its LVJ are both highly variable. The TD commonly joins the internal jugular vein, the subclavian vein, or enters directly at the venous angle.5,18 The TD may exist as a single vessel, or have multiple branches, each with their own communication with the veins.5,18 A terminal dilatation (ampulla, ampullary dilatation) of the TD is frequently described,5,19–25 and may be present in as many as 40% of individuals. 18 Some studies have reported the TD terminating in a lymph sac.26,27
TD valves
Semilunar valves are variably distributed along the course of the TD, with a preponderance in its cervical portion.5,6 These valves are typically bicuspid, though tricuspid and monocuspid valves may also occasionally form.5,6 Valve position has been associated with sites of increased pressure due to compression from adjacent mediastinal structures, such as where the TD is crossed by the aorta and esophagus.21,28
Characteristics of the LVJ
The TD may enter the venous lumen obliquely or perpendicularly19,20,25 and typically narrows at the LVJ.25,29 The LVJ diameter has limited extensibility compared with the proximal TD lumen.22,25 The TD may travel within the wall of the vein before communicating with the venous lumen (Fig. 2).19,20,24,30 The course of the terminal TD within the vein wall has thus been likened to the junction between the ureter and bladder,19,31 and the ampulla of Vater within the duodenal wall. 20 Zavgorodnii et al. 24 found that the smooth muscle cells of this intramural segment formed a discrete sphincter muscle.
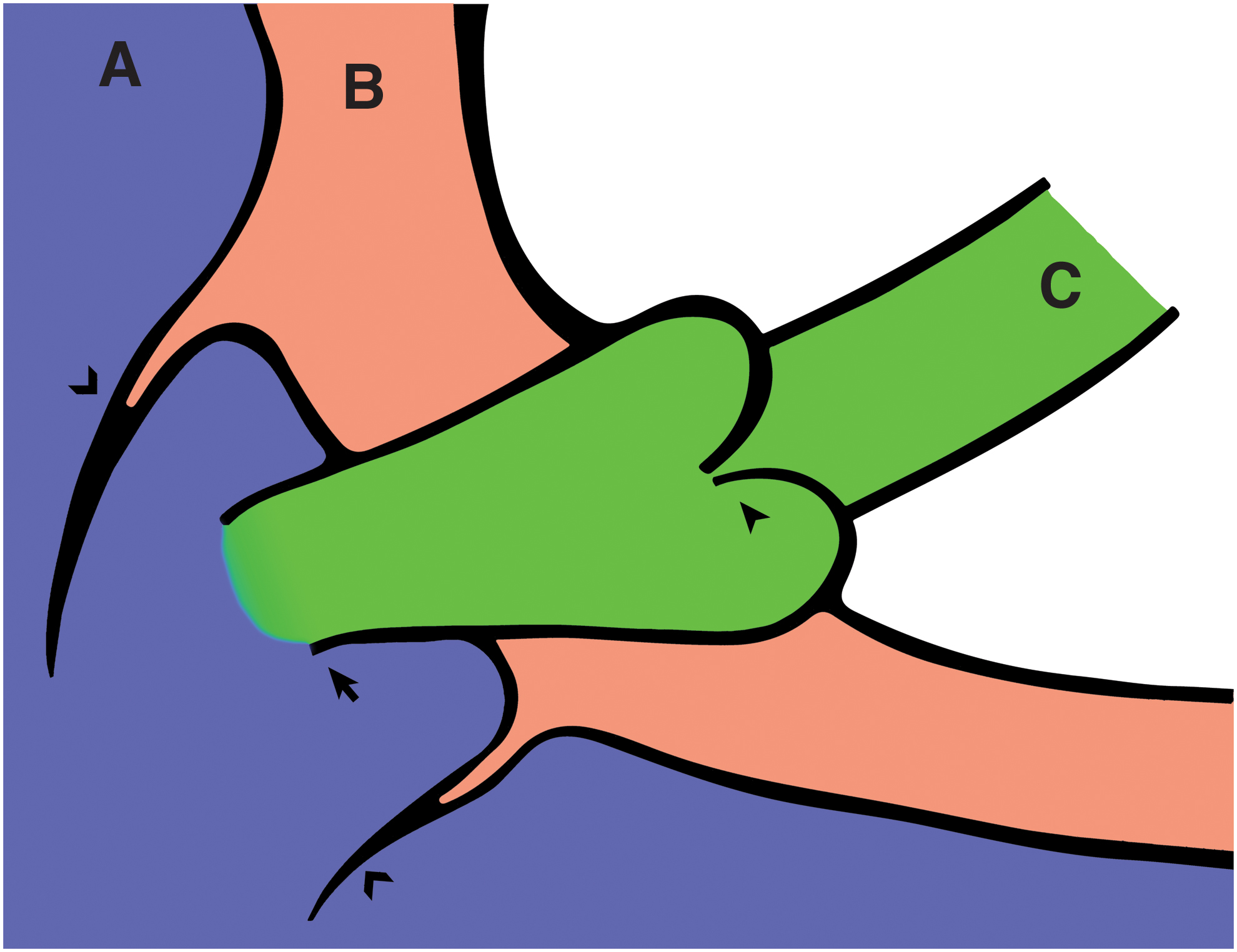
The intramural relationship of the terminal TD with the vein wall. The figure is “magnified,” showing the terminal most segment of the TD traversing the vein wall.
Other authors have described the TD joining at convex outpouchings of the veins.5,23 Jacobsson 5 found this was most common when there was no valve at the LVJ.
Relationship between the LVJ and adjacent venous valves
Numerous studies have noted the proximity of the LVJ to the large venous valves present in the internal jugular and subclavian veins19,20,24,31–34 (Fig. 2). Parsons and Sargent 20 found the LVJ was located under a cusp of the internal jugular vein valve in 65% of cases (26/40).
Existence of a valve at the LVJ
An LVV is thought to guard the mammalian LVJ, 7 however, close examination of the literature suggests such a valve is not consistently present in humans. Histological studies of human embryos have reported the LVJ forming a valve around the sixth week of development,35,36 however, Kampmeier 31 found that a typical or truly competent valve was frequently absent at the finished LVJ in 4-month old fetuses.
In adults, Parsons and Sargent 20 found the vast majority of LVJs to lack any definite morphological valves (n = 40). Jacobsson 5 found a valve positioned at the LVJ in 59%, valves within 1–6 mm of the LVJ in 40%, and no valves in the terminal TD in 1% of cases studied (n = 139). Miyakoshi et al. 6 found valves at 52% of LVJs (n = 36) and the proximal (upstream) lymphatic valve may be located up to 30 mm from the LVJ. Across 1081 postmortem examinations, Van Pernis 37 reported the terminal valve of the TD to exist anywhere in the last 10 mm of the duct. Cui 26 found 35% of terminal TDs to be valveless (n = 40). Other cadaveric studies place LVV occurrence between 74% and 100%.24,33,34,38,39 Using in vivo ultrasonography, Seeger et al. 25 could visualize the terminal TD in 96% of cases, but valves located directly at the LVJ were only seen in 40% of individuals in the study (n = 585).
Various morphologies of the LVV
Detailed studies of the morphology of the LVV in humans are scarce and the extant literature offers a discordant picture.
Jacobsson 5 examined the valves of the TD in 100 human cadavers and classified them into 5 different morphologies (Fig. 3). The LVV was typically an asymmetrical bicuspid semilunar valve, with one leaflet wider and deeper than its pair (Fig. 3D). This was especially manifest when the duct had an oblique insertion into the veins. The smaller valve cusp was sometimes so small that the valve could be mistaken for a monocuspid valve.
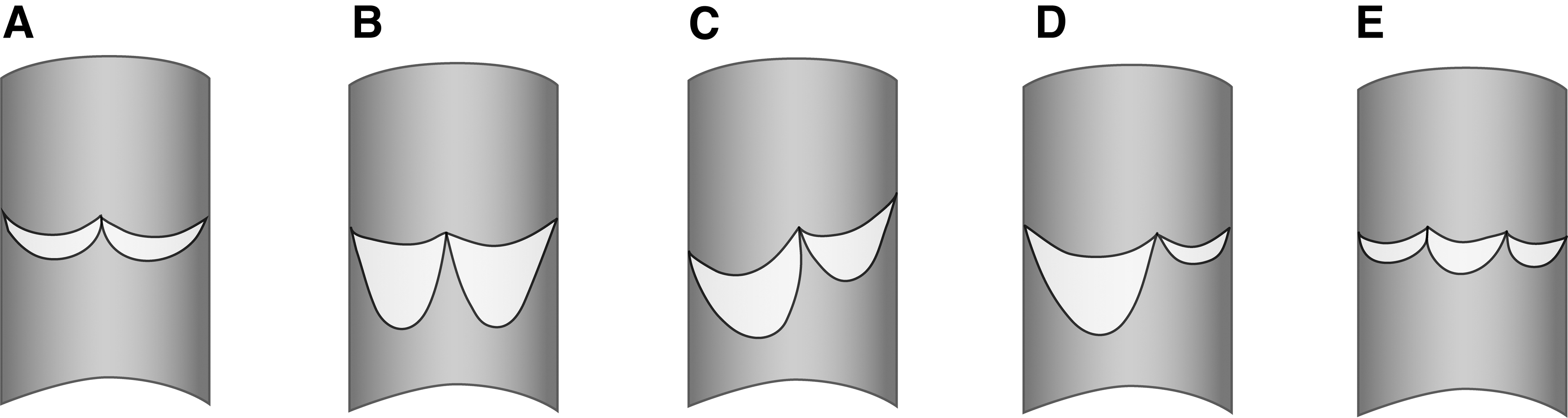
The different types of valves found in the TD.
Miyakoshi et al. 6 found 79% of LVVs to be bicuspid semilunar valves, 16% falciform (sickle shaped), and 5% were an inverted bicuspid semilunar valve (n = 19). In a study of five cadavers, Chiba et al. 27 described a complex multileafed funnel-shaped LVV, comprising large primary and small secondary leaflets. Bartholin 40 described the LVV as miter like.
Some studies have described the LVV as having a unique “ostial” morphology, distinct from the more proximal (upstream) semilunar valves (Fig. 4).33,34,38 Ostial cusps were said to lie flush with the vein wall at the LVJ. 33 El Zawahry et al. 34 found each ostial leaflet to be 10 mm in length.
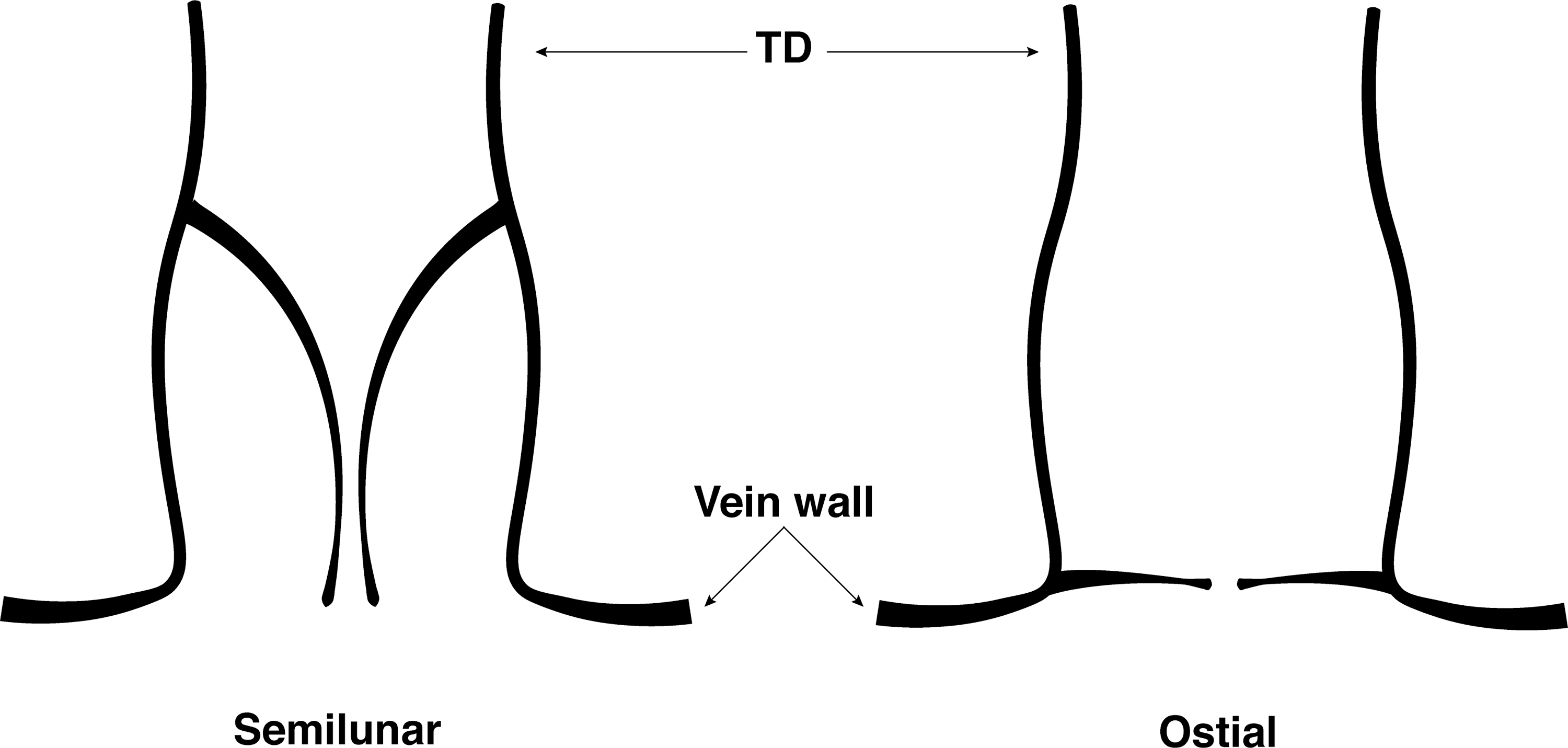
Buy and Argaud 19 examined the TD of four cadavers and found a monocuspid ostial valve at three of the LVJs, and a bicuspid ostial valve at one, which was likened to the ileocecal valve (Fig. 5).

Various modes of TD termination and lymphovenous valve morphology.
Other authors have described the TD narrowing at its LVJ with a single flap-like structure forming across the lumen (Fig. 6).29,31,41 In a study of 100 cadavers, Shimada and Sato 29 found the flap-like structure was relatively thick compared with the lymphatic valves present further upstream in the TD.
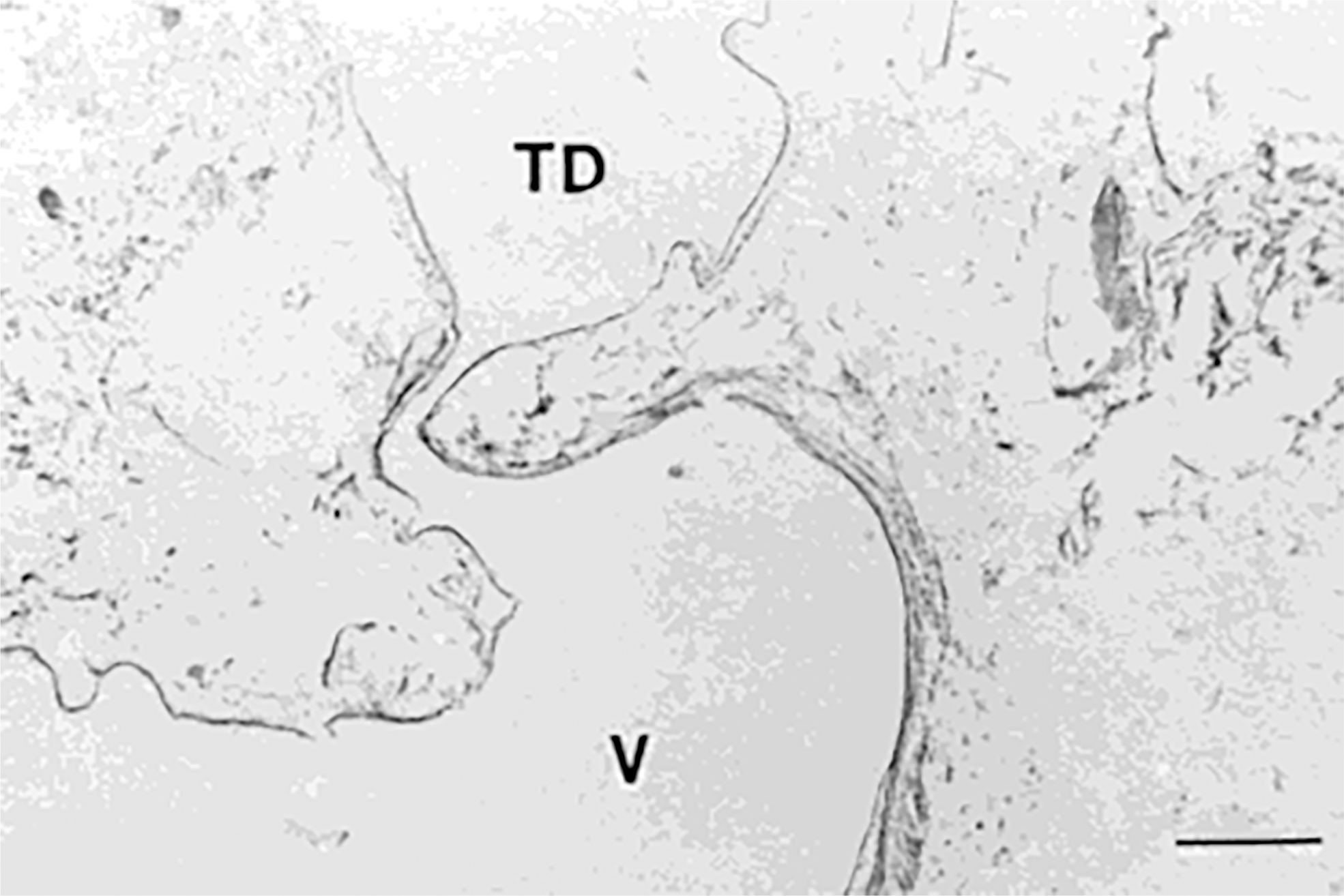
Photomicrograph of the lymphovenous junction, Elastic-van Gieson staining. Scale bar = 300 μm. The TD typically narrows at the lymphovenous junction, where a flap-like structure may be found. TD, thoracic duct; V, venous lumen. Reproduced from Shimada and Sato, 29 used with permission courtesy of Wiley and Sons.
Microanatomy of the LVV
Each cusp of the LVV is formed by a continuous projection of endothelium that envelops a core of connective tissue matrix, similar to regular lymphatic valves.24,33,34 The flap-like variant differed from the lymphatic valves by having a relatively thick subendothelial connective tissue zone (Fig. 6).29,41 Smooth muscle has been described extending into the base of the valve cusps,24,27,34 and Zavgorodnii et al. 24 found the smooth muscle myocytes formed a discrete valve tensor around the LVV.
The LVV in other mammals
LVV morphology appears to vary both within and between mammalian species. In dogs, monocuspid valves,42–44 bicuspid valves,33,45 and a monocuspid valve overlying a bicuspid valve 46 have all been reported. Bannykh et al. 45 found the canine and feline LVV to be an insufficient bicuspid valve, either crescent shaped or navicular. These valves possessed prominent buttresses and thickened free edges. In some older animals, the leaflets were seen to fuse into a funnel-shaped morphology. 45 Bicuspid valves have been described in mice 47 and monkeys. 48 Slit-shaped and papilliform junctions have been directly visualized in pigs. 49 Murine embryological studies have shown that the inner layer of the LVV leaflet is derived from lymphatic endothelial cells, whereas the outer layer forms from a subpopulation of venous endothelial cells and is continuous with the vein wall.7,50
Functional Anatomy
The function of the LVV is to allow antegrade lymph flow and prevent retrograde blood flow into the TD. The mechanism by which it does this is not clear, and its role is made more opaque by the fact that the valve is frequently absent, and when it is present it has a highly variable morphology. This section outlines the findings of previous studies that have investigated the mechanism of the LVV.
Relationship between valve morphology and mechanism
A bicuspid semilunar LVV forms a sinus downstream and to the leeward side of each valve cusp. No studies found in this review specifically examined the open/close mechanism of such a morphology, but its similarity to bicuspid semilunar valves seen elsewhere in the lymphatic (and venous) system would suggest its fundamental mechanism is based on retrograde flow filling the valves sinuses, causing the cusps to balloon out and obstruct the lumen. Ultrasonography has allowed for visualization of the human LVJ and LVV in vivo (Fig. 7),25,51–56 but no studies give a description of the valve's precise mechanism. In some of the ultrasound videos presented by Seeger et al., 25 reverse flow can be seen filling the semilunar valve sinuses, causing the cusps to balloon out and close off the LVJ.
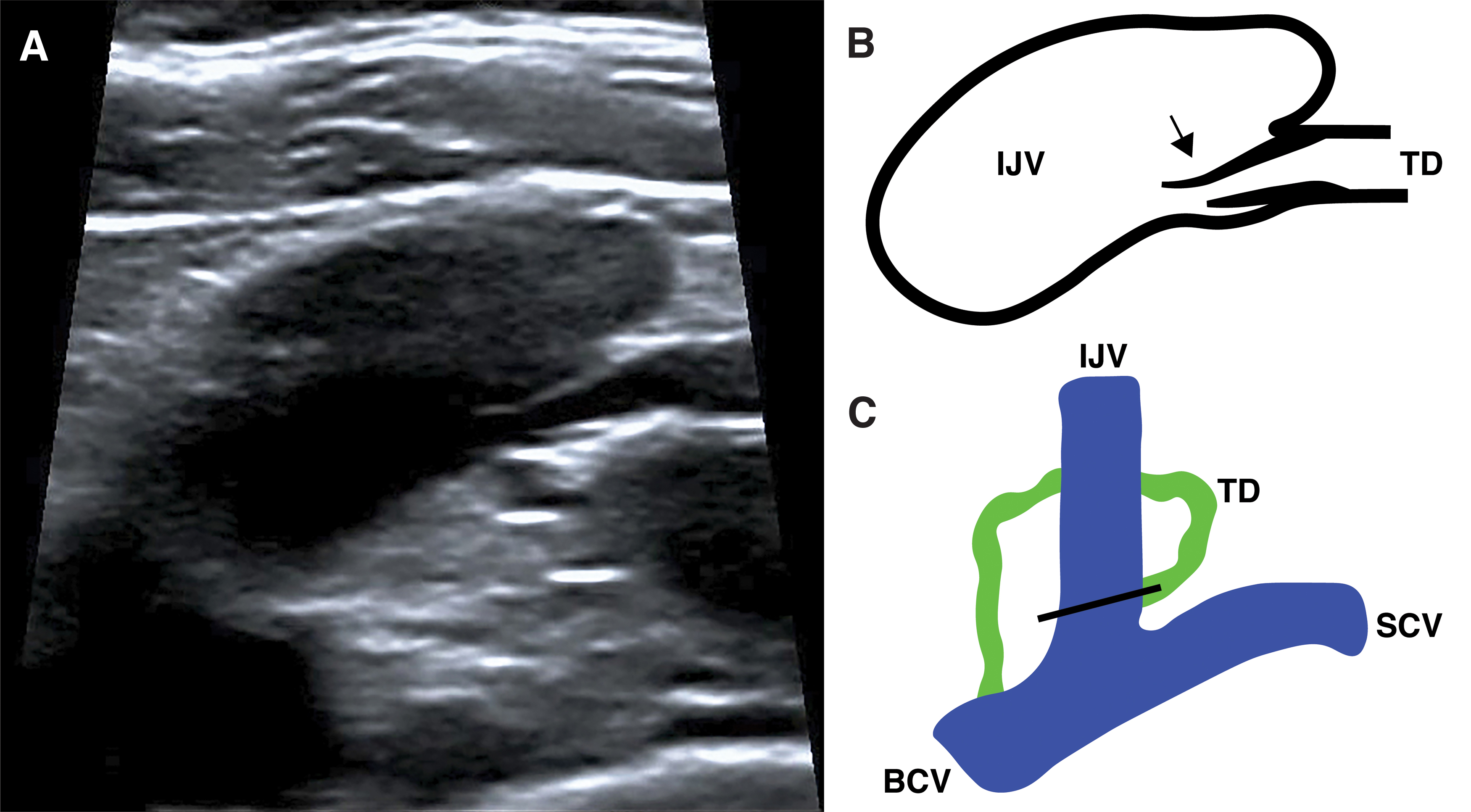
Ostial morphology leaflets differ from semilunar leaflets as they are thought to open and close based on the tension of the surrounding vein wall, rather than reverse flow filling sinuses. This is based on the study of Pflug and Calnan 33 who dissected and examined 20 canine and 7 human cadavers. The veins were opened and the LVJ was examined directly. When the vein wall was stretched, the ostial leaflets became taut across the orifice, closing off the LVJ. When the tissue was lax, the leaflets discretely collapsed in line with the vein wall like folding doors (Fig. 8). The authors, therefore, concluded that the state of the valve was dependent on venous wall tension, rather than reverse flow filling valve sinuses. 33

Proposed mechanism of the ostial valve morphology. The valve closes when the vein is under tension (left). The valve opens when the vein is lax (right).
Valve mechanism in relation to the respiratory and cardiac cycles
The effect of the respiratory and cardiac cycles on lymphatic outflow remains unclear. Direct endoscopic visualization of the canine LVJ in vivo,44,57 and cinelymphography in humans 58 have demonstrated lymph flowing across the LVJ during inspiration. Other in vivo studies were ambiguous, reporting an association between lymph efflux and respiration, but not clearly stating when during the respiratory cycle flow actually occurred.22,23,49,59,60 Based on canine TD pressure measurements, Pflug and Calnan 33 concluded that flow across the LVJ was probably continuous but greatest during expiration, and that the LVV functions to prevent blood reflux during extreme increases in central venous pressure (e.g., coughing, sneezing, and Valsalva maneuver). Bannykh et al. 45 reference a personal correspondence with Tzib, who, using cinelymphography in humans, observed that lymph efflux occurs independently of the respiratory cycle, five to seven times per minute. More recently, in vivo ultrasonography in humans has observed lymphatic outflow occurring during expiration, 56 however, subjects were children with congenital heart disorders.
No studies have specifically investigated the effect of the cardiac cycle on the LVV in humans, but ultrasonography videos 25 of the valve show it opening and closing in sync with the nearby venous valves, suggesting the valve may be synchronized with the transmitted pressure waves of the cardiac cycle.
Other potential mechanisms for preventing blood reflux
Buy and Argaud 19 reported insufficient monocuspid ostial LVVs, but it was reasoned that the intramural course of the terminal TD across the vein wall worked to overcome this apparent inadequacy. By obliquely traversing the vein wall (c.f. ureterovesical junction), it was hypothesized that the intramural TD would be subject to the distension of the vein at times of increased venous pressure, which would obliterate the TD lumen and render blood reflux impossible. Other authors have made similar inferences,20,30,31 but such a mechanism is yet to be confirmed in vivo (Buy and Argaud 19 also described longitudinal twisting of the terminal TD in one cadaver [Fig. 5C], which was similarly hypothesized to retard retrograde blood flow).
A physiological clotting mechanism, termed “lymphovenous hemostasis,” has recently been proposed to operate at the mammalian LVJ. Hess et al. 47 demonstrated in mice that platelet plugs intermittently form at the LVJ, in the absence of any vascular injury, and that these functional thrombi, as well as LVVs, are required to prevent blood from entering the lymphatic system. It was demonstrated that lymphovenous hemostasis can compensate for loss of a functional LVV, as when LVV development was impaired, more extensive clot formation occurred, which limited venous backflow across the LVJ. 47 Understanding of lymphovenous hemostasis remains in its infancy. Key questions such as how the platelet plugs allow for antegrade lymph flow, but at the same time prevent blood reflux, remain unknown. 61
Conclusions
This review examined the structure and function of the LVJ and LVV of the TD. The conventional understanding to date has been that the TD terminates in a bicuspid valve situated at its LVJ. However, it became apparent from this review that the LVV shows marked anatomical heterogeneity with widely variable descriptions present in the literature, including bicuspid semilunar, ostial, and flap-like morphologies. Surprisingly, a discrete valve may be frequently absent at the human LVJ. There was diverse opinion on how the TD functionally allows lymphatic outflow and prevents blood reflux. The respiratory cycle has long been considered a key determinant of flow across the LVJ, but empirical observation through various methodologies in humans and animals paints a conflicting picture about how inspiration and expiration are related to LVV mechanism and lymph flow. The mechanism of this physiologically and clinically important structure, therefore, remains surprisingly unclear in humans.
Recent literature62,63 has focused on the ostial LVV morphology in humans. The larger anatomical studies5,6 found in this review indicated that the human LVV, when present, was typically a bicuspid semilunar valve. This would suggest that retrograde flow filling the valve sinuses is fundamentally responsible for the valve's mechanism, similar to venous valves.64,65 Of course the mechanism of the LVV is likely to vary with its morphology, but recent ultrasonography of the human LVV in vivo appears to show the valve functioning in a manner consistent with a semilunar morphology. 25 This is in contrast to the conventional ostial valve concept, which is based on the tension of the vein wall resulting in valve closure, rather than retrograde flow filling valve sinuses. 33 It is noteworthy that evidence for the ostial valve mechanism is largely based on canine studies 33 and no study reporting ostial LVVs has been able to clearly demonstrate the morphology in humans.33,34,38
The LVV is believed to safeguard the TD from blood reflux.7,47 The frequent absence of the LVV in humans is, therefore, interesting. Other factors may compensate for an absent valve, such as a ureterovesical junction-like mechanism,19,31 but functional studies are needed to confirm this. Alternatively, perhaps LVV absence implies blood reflux is not necessarily deleterious and may be a more physiological than a pathophysiological event.
This review highlighted the large venous valves of the internal jugular and subclavian veins, which frequently overlie the LVJ. Further study is required to better understand the functional significance of these proximate venous valves. Knowledge of these valves may be relevant to the success of minimally invasive retrograde TD interventions.
It is noteworthy that this review yielded no clear and direct photographic depictions of the LVJ and LVV in humans. Histology, ultrasonography, comparative morphology, artists drawings, and author descriptions all offer glimpses, but because direct visual evidence of the structure is lacking, it is difficult to discern the precise structure and function of this part of the human body. Considering the growing clinical significance of the LVJ, further investigation of its fundamental form and function is merited. Only through dedicated in vivo studies, using sophisticated modern imaging techniques, such as high-definition ultrasonography, will it be possible to corroborate or refute the wide range of descriptions and concepts present in the literature.
Footnotes
Author Disclosure Statement
No competing financial interests exist.
Funding Information
No funding was received for this study.