
Abstract
Background:
This study investigated the expression and localization of cathepsins B, D, and G in relationship to the embryonic stem cell (ESC)-like population we have previously identified in microcystic lymphatic malformation (mLM).
Methods and Results:
Immunohistochemical staining demonstrated expression of cathepsins B, D, and G in cervicofacial mLM tissue samples from 11 patients. Immunofluorescence staining of two representative mLM samples showed localization of cathepsins B and D to the OCT4+ and the c-MYC+ cells on the endothelium of lesional vessels and the stroma, while cathepsin G was localized to the OCT4+/tryptase+ cells within the stroma. Transcript expression of cathepsins B, D, and G was confirmed using reverse transcription quantitative polymerase chain reaction (RT-qPCR; n = 5). Western blotting (n = 3) performed on the mLM tissue samples revealed protein expression of cathepsins B and D, which were demonstrated to be enzymatically active using enzymatic activity assays.
Conclusion:
This study demonstrated expression of cathepsins B and D by the ESC-like cells on the endothelium of lesional vessels and the stroma, while cathepsin G was localized to the OCT4+ phenotypic mast cells within the stroma of mLM.
Introduction
Lymphatic malformation (LM) is a low-flow vascular malformation affecting ∼1/2000–1/4000 live births. 1 Fifty percent of LM are evident at birth, with up to 90% being diagnosed by the end of the second year, 2 or later in life by the presence of symptoms, often precipitated by infection, trauma, or hormonal changes. 3
Seventy-five percent of LM occur in the head and neck 4 and other common sites include the axilla, chest, and perineum. 5 LM may become infected, cause bleeding, psychosocial stress, 5 and mass effects on adjacent structures.6,7
LM may be classified as macrocystic (cysts >1 cm), microcystic (cysts <1 cm), or mixed. 1 Microcystic lymphatic malformation (mLM) poses a management challenge because of its tendency to affect deep structures and its association with recurrence rates of 50%–66%. In addition, mLM often cross anatomic regions and tissue planes and can be intimately associated with hollow visceral and solid organs, thus complicating therapeutic interventions. 3 Histologically, LM is composed of thin-walled, cystic, and dilated channels lined by lymphatic endothelial cells, filled with proteinaceous lymph fluid. 2
LM is thought to result from aberrant lymphatic morphogenesis related to failure of sequestered lymphatic tissue to communicate with the lymphatic and venous systems after budding.6–8
Intervention for LM is indicated for symptomatic lesions causing pain or deformity, or posing a threat to vital structures. 5 Treatment modalities for mLM include surgical resection, sclerotherapy, and laser therapy. 9 While surgical excision is recommended for operable lesions, 10 the recurrence rate is up to 22%. Several sclerosants have been used, including alcohol, bleomycin, doxycycline, polidocanol, sodium tetradecyl sulfate and OK432 (picibanil), which cause inflammation of the vessel wall, leading to fibrosis with subsequent obliteration of the lumen of the vessels. Sclerotherapy may be used in conjunction with other therapies such as surgical resection and radiofrequency ablation. 10
We have previously identified an embryonic stem cell (ESC)-like population within mLM that expresses the ESC markers OCT4, SOX2, NANOG, KLF4, and c-MYC on the endothelium of lesional vessels with abundant expression of c-MYC and SOX2, which were also present on cells within the stroma. 12
The renin–angiotensin system (RAS) is classically known for its role in regulation of blood pressure and body fluids homeostasis. 13 Expression of the RAS has been observed in venous malformation,14,15 infantile hemangioma (IH), 16 and pyogenic granuloma 17 There is increasing evidence of the role of the RAS in angiogenesis, prosurvival signaling, stem cell proliferation, and tumor invasion.13,18 Cathepsins B, D, and G are proteases that form bypass loops of the RAS,13,19 contributing to the generation of angiotensin peptides without involvement of the classical RAS cascade. 20 Expression of components of the RAS, 16 as well as cathepsins B, D, and G has been demonstrated in IH. 19
The presence of cathepsins has been observed in various vascular anomalies.21–23 Increased expression of cathepsins B, K, and S in rat and human cerebral aneurysms has been observed. 22 Cathepsins have also been implicated in the pathogenesis of abdominal aortic aneurysm expansion and rupture via degradation of extracellular matrix proteins. 21 Moreover, increased expression of cysteinyl cathepsins has been noted in human varicose veins. 23 We have shown expression and localization of cathepsins B, D, and G to the ESC-like population on the microvessel endothelium within keloid-associated lymphoid tissues of keloid lesions. 24 Cathepsins B and D have also been localized to the ESC-like population in the endothelium of microvessels in Dupuytren's disease. 25
We hypothesized that cathepsins B, D, and G may be expressed by the ESC-like population we have previously identified in mLM. 12 In this study, we investigated the expression and localization of these cathepsins in relationship to the primitive population within mLM using immunohistochemical and immunofluorescence staining, western blotting and reverse transcription quantitative polymerase chain reaction (RT-qPCR). Enzymatic activity assays were performed to investigate the enzymatic activity of these cathepsins.
Materials and Methods
Tissue samples
Cervicofacial mLM tissue samples from 11 patients included in our previous study 12 were obtained from the Gillies McIndoe Research Institute Tissue Bank for this study, which was approved by the Central Health and Disability Ethics Committee (Ref. 13/CEN/130). Written informed consent was obtained from all participants.
Histology and immunohistochemical staining
The diagnosis of LM for all the 11 samples was confirmed by hematoxylin and eosin (H&E) staining and positive staining for the lymphatic endothelial marker D2-40 (1:100; cat. no. M3619; Dako, Glostrup, Denmark) of 4 μm-thick formalin-fixed paraffin-embedded consecutive sections of mLM. 26 Immunohistochemical staining was then performed on the Leica BOND™ RX autostainer (Leica) with primary antibodies for cathepsin B (1:1000; cat. no. sc-6490-R; Santa Cruz), cathepsin D (1:200; cat. no. NCL-CDm; Lecia), cathepsin G (1:200; cat. no. sc-33206; Santa Cruz), OCT4 (1:30; cat. no. MRQ-10; Cell Marque, Santa Cruz, CA), c-MYC (1:1000; cat. no. 9E10; Abcam, Cambridge, UK), and tryptase (1:300; cat. no. NCL-MCTRYP-428; Leica) with 3,3′-diaminobenzidine as the chromogen. Antibodies diluted and antigens retrieved using a protocol previously described. 25
To further localize the expression of the cathepsins, two representative samples of mLM of the original 11 patients underwent immunofluorescence staining. Immunofluorescence dual-staining was performed using the same primary antibodies at the same concentrations used in immunohistochemical staining. Appropriate secondary antibodies were selected (donkey anti-mouse Alexa-488 or donkey anti-rabbit Alexa-594; Life Technologies). Immunofluorescence-stained slides were mounted in VECTASHIELD HardSet Antifade Mounting Medium and counter-stained with 4′6-diamino-2-phenylinodole (cat. no. H-1500; Vector Laboratories).
Positive controls for immunohistochemical staining for cathepsins B and D were performed on human placenta and breast carcinoma, respectively, and cathepsin G and tryptase on human tonsil. Negative controls for immunohistochemical staining was performed using a matched immunoglobulin G/immunoglobulin M isotype control for mouse (cat. no. IR750; Dako, Copenhagen, Denmark) and rabbit (cat. no. IR600; Dako) primary antibodies, to assess specificity of the amplification cascade. Negative controls for immunofluorescence staining was performed using a combination of primary isotype mouse (ready-to-use; cat. no. IR750; Dako) and rabbit (ready-to-use; cat. no. IR600; Dako) antibodies.
Western blotting
Total protein was extracted and precipitated from seven snap-frozen mLM samples of the original cohort of 11 patients, and subsequently separated by sodium dodecyl sulfate-polyacrylamide gel electrophoresis, then transferred to polyvinylidene difluoride membrane for detection as previously described. 27 iBind Flex (cat. no. SLF2000; Thermo Fisher Scientific) was used for antibody binding with the primary antibodies for cathepsins B, (1:250; cat. no. SC-6490-R; Santa Cruz), cathepsin D (1:250; cat. no. ab75852; Abcam), cathepsin G (1:500; cat. no. ab197354; Abcam), and α-tubulin (1:1000; cat. no. 62204; Thermo Fisher Scientific). Appropriate secondary antibodies were goat anti-rabbit Alexa Fluor 647 (1:2000; cat. no. A21244; Life Technologies) for cathepsins B and D, goat anti-rabbit horseradish peroxide (HRP; 1:2000; cat. no. ab6721; Abcam) for cathepsin G, and goat anti-mouse Alexa Fluor 488 (1:2000; cat. no. A21202; Life Technologies) for α-tubulin. Clarity Western enhanced chemiluminescence (cat. no. 1705061, Bio-Rad) was used as the substrate for visualizing HRP-detected protein bands and the ChemiDoc MP Imaging System (Bio-Rad) and Image Lab 6.0 software (Bio-Rad) was used for band detection and analysis. Snap-frozen human tonsil was used as control tissue for cathepsins B and D with a recombinant cathepsin G protein (cat. no. H00001511-Q01; Novus Biologicals, Littleton, CO) as the control for cathepsin G. Matched mouse (1:500; cat. no. ab18443; Abcam) and rabbit (1:500; cat. no. ab171870; Abcam) isotype controls were used as appropriate negative controls.
Enzymatic activity assays
Enzymatic activity of cathepsins B and D was determined in the same three mLM samples used for western blot analysis, using enzyme activity assay kits for cathepsin B (cat. no. ab65300; Abcam) and cathepsin D (cat. no. ab65302; Abcam). All steps of the procedure were performed as previously described. 25 Fluorescence was measured in a black, clear bottomed 96-well plate (cat. no. 3631; Corning, ME) using the Varioskan Flash plate reader (cat. no. MIB5250030; Thermo Fisher Scientific). Human tonsil and denatured tonsil tissue lysates were used as appropriate positive and negative controls, respectively.
Reverse transcription quantitative polymerase chain reaction
Total RNA was isolated from five snap-frozen mLM tissue samples of the original cohort of 11 patients. Approximately 20mg of the tissue was suspended in 350 μL lysis buffer RLT (Qiagen) and homogenized using the Omni Tissue Homogenizer (Omni TH; Omni International, Kennesaw, GA). Homogenized tissues were prepared using the RNeasy Mini Kit (Qiagen) on the QIAcube system (Qiagen). The quantity and quality of the RNA was determined using a NanoDrop 2000 Spectrophotometer (Thermo Fisher Scientific). The expression of cathepsin genes was analyzed using the Rotor-Gene Q (Qiagen) and the Rotor-Gene Multiplex RT-PCR Kit (Qiagen). The primer probes used for cathepsin B (HS00947439_M1), cathepsin B (HS00157205_M1), and cathepsin G (HS00175195_M1) (cat. no. 4331182; Thermo Fisher Scientific). Gene expression was determined against the housekeeping gene glyceraldehyde 3-phosphate dehydrogenase (GADPH) (Hs99999905_m1; cat. no. 4351370; Thermo Fisher Scientific). Endpoint amplification products were checked for the presence of single bands of the correct size by agarose gel electrophoresis (cat. no. G402001; Thermo Fisher Scientific).
Results
Histology and immunohistochemical staining
H&E staining showed mLM consisted of irregular, dilated vessels lined by flat endothelium (Supplementary Fig. S1A) with positive staining for the lymphatic endothelial marker D2-40 (Supplementary Fig. S1B, brown) in all 11 tissue samples. Immunohistochemical staining demonstrated predominantly cytoplasmic expression of cathepsin B (Fig. 1A, brown) and cathepsin D (Fig. 1B, brown) on both the endothelium of the lesional vessels and the cells within the stroma. Cytoplasmic expression of cathepsin G (Fig. 1C, brown) and tryptase (Fig. 1D, brown) was predominantly localized to cells within the stroma.
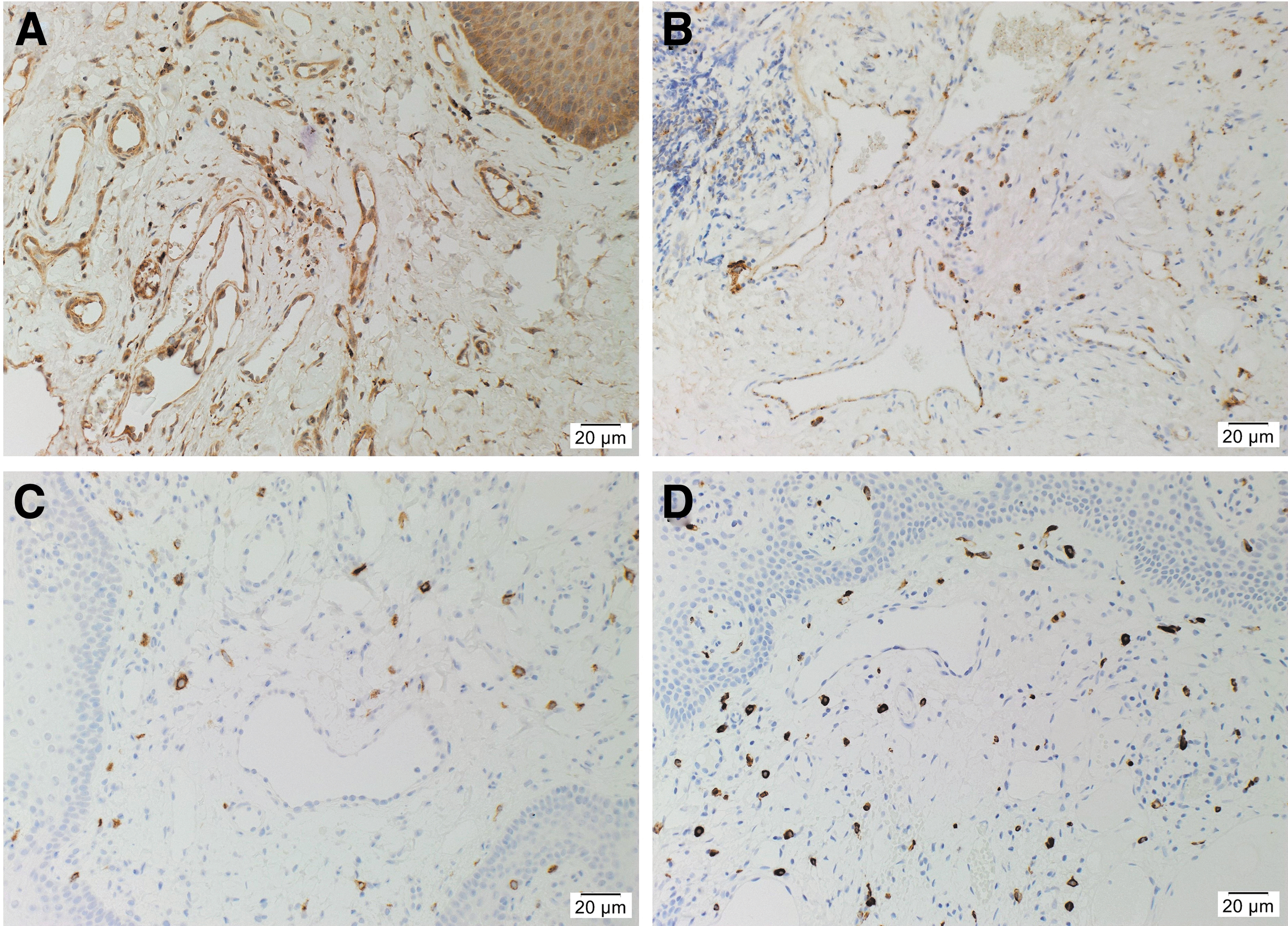
Representative immunohistochemical-stained sections of microcystic lymphatic malformation (mLM) showing expression of cathepsin B
Positive controls for cathepsin B (Supplementary Fig. S1C, brown) and cathepsin D (Supplementary Fig. S1D, brown) demonstrated the expected staining patterns in human placenta and breast carcinoma, respectively, and cathepsin G (Supplementary Fig. S1E, brown) and tryptase (Supplementary Fig. S1F, brown) in human tonsil. The negative control demonstrated no staining (Supplementary Fig. S1G) confirming the specificity of the primary antibodies used.
Immunofluorescence staining
Immunofluorescence staining performed on two representative mLM tissue samples of the original cohort of 11 patients demonstrated expression of cathepsin B (Fig. 2A, B, red) by the OCT4+ (Fig. 2A, green) and the c-MYC+ (Fig. 2B, green) cells within the endothelium of the lesional vessels (arrows) and the cytoplasm of the cells within the stroma (arrowheads). Cathepsin D (Fig. 2C, D, red) was also expressed by the OCT4+ (Fig. 2C, green) and the c-MYC+ (Fig. 2D, green) cells within the endothelium of the lesional vessels (arrows) and the cytoplasm of the cells within the stroma (arrowheads). Cathepsin G (Fig. 2E, F, red) was expressed by the tryptase+ cells (Fig. 2E, green, arrowheads) within the stroma, majority of which also expressed OCT4 (Fig. 2F, green, arrowheads) suggesting a primitive mast cell phenotype.
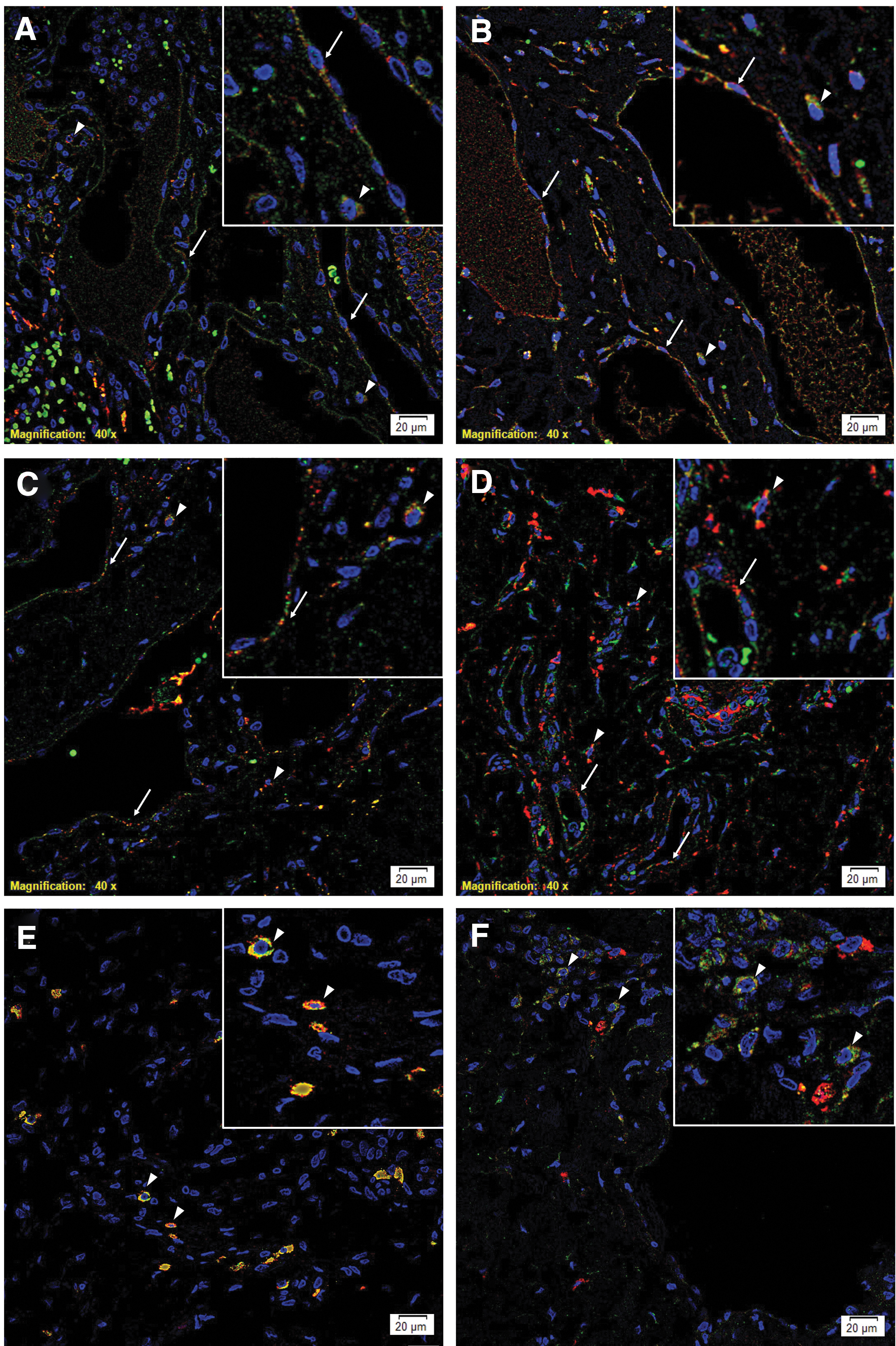
Representative immunofluorescence-stained sections of microcystic lymphatic malformation (mLM) demonstrating cytoplasmic expression of cathepsin B
Images of individual stains of Figure 2 are presented in Supplementary Figure S2. Minimal staining was present on the negative control (Supplementary Fig. S2M), confirming the specificity of primary antibodies used.
Western blotting
Western blotting of three snap-frozen mLM samples of the original cohort of 11 patients demonstrated the presence of cathepsin B (Fig. 3A, blue) and cathepsin D (Fig. 3B, blue) in all three mLM samples at the expected molecular weight of 25 kDa and 28 kDa, respectively. Tonsil tissue was used as positive control for cathepsins B and D. Equal loading of the samples was demonstrated by α-tubulin (Fig. 3A, B, green). Cathepsin G (Supplementary Fig. S3A, red) was not detected in all three mLM samples investigated.

Representative western blot images of total protein extracted from three microcystic lymphatic malformation tissue samples demonstrating the presence of cathepsin B at 25 kDa
Enzymatic activity assays
Enzymatic activity assays on the same three snap-frozen mLM tissue samples included in WB analysis demonstrated enzymatic activity for both cathepsins B and D, relative to that of the positive control tissues (Fig. 4).
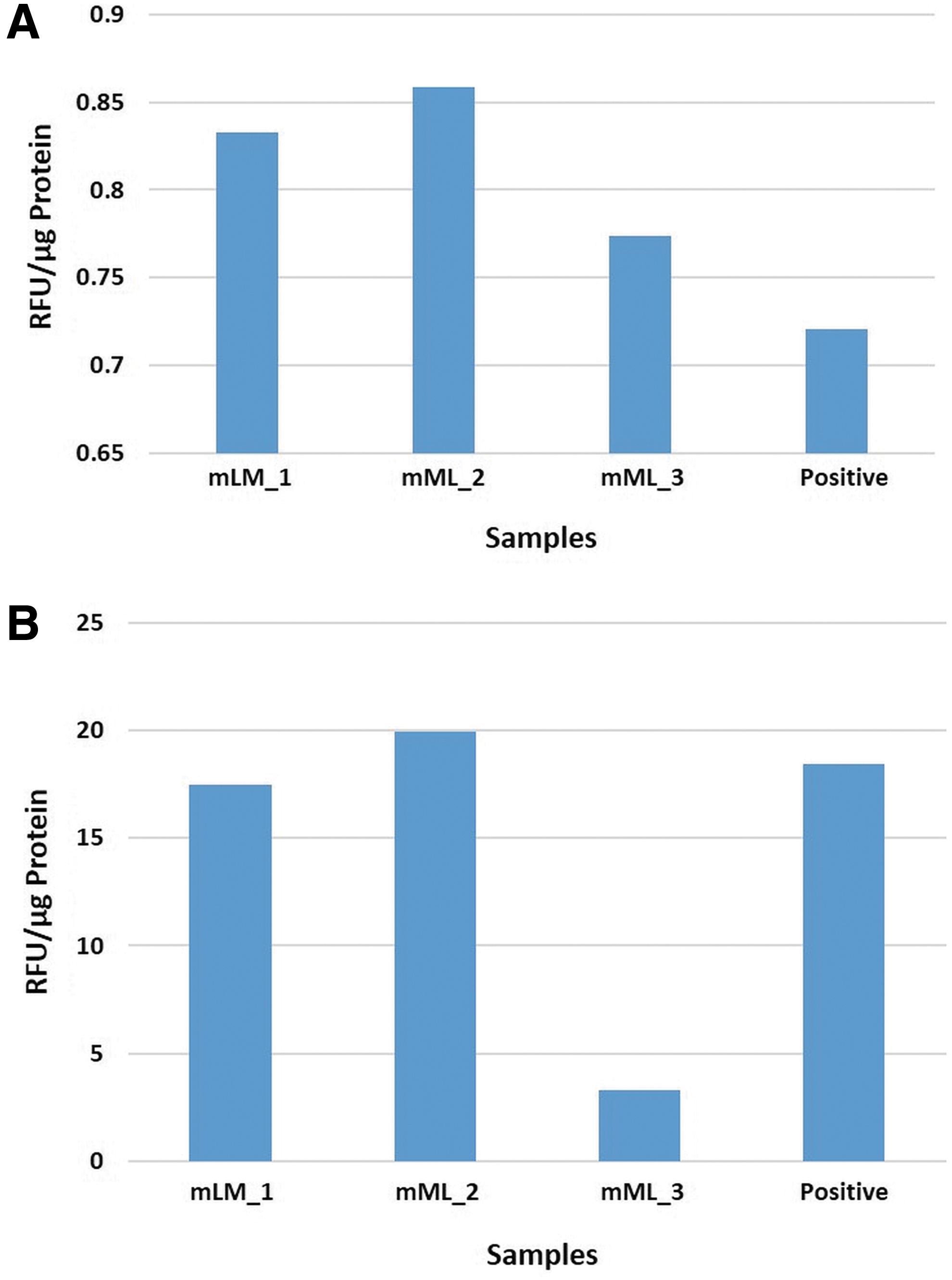
Enzymatic activity assays performed on three microcystic lymphatic malformation tissue samples showed enzymatic activity of cathepsin B
Reverse transcription quantitative polymerase chain reaction
RT-qPCR demonstrated the presence of messenger RNA transcripts for cathepsins B, D, and G (Fig. 5) in all five mLM tissue samples examined. The delta cycle threshold was determined against the housekeeping gene GAPDH. Specific amplification of the primers was demonstrated by electrophoresis of PCR products on 1% agarose gels, with the expected sized amplicons observed, and no products observed in any template control lanes (data not shown).
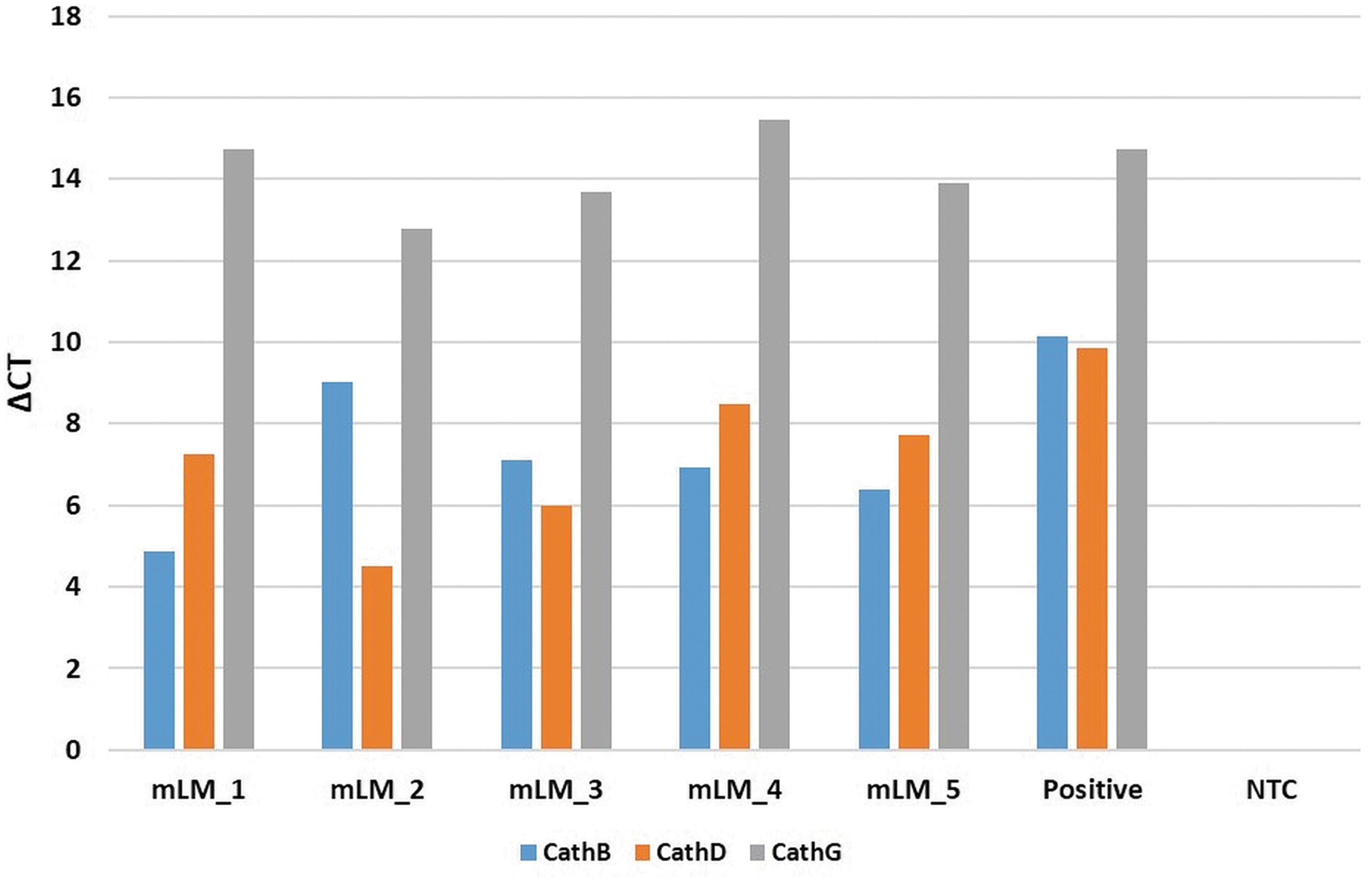
Reverse transcription quantitative polymerase chain reaction delta cycle threshold (ΔCT) values demonstrating transcript expression of cathepsin B (blue), cathepsin D (orange), and cathepsin G (gray) in microcystic lymphatic malformation (mLM) tissue samples. ΔCT values were calculated by comparing CT values of cathepsins B, D, and G against the housekeeping gene glyceraldehyde 3-phosphate dehydrogenase (GAPDH), with graphs displaying average ΔCT values of triplicate trials run on each sample. Positive controls were demonstrated on tonsillar tissue, and specificity of probes was confirmed using a no template control (NTC). Endpoint amplification products were checked for the presence of bands at the correct size by 1% agarose gel electrophoresis.
Conclusion
The presence of cathepsins B and D proteins in mLM tissue samples demonstrated by immunohistochemical staining was confirmed by western blotting, and enzymatic activity assays showed that these cathepsins were enzymatically active. Immunohistochemical staining also showed localization of cathepsin G to cells within the stroma, although it was below detectable levels by western blotting analysis, which may be attributed to low levels of the protein within the mLM tissue samples. The transcript expression of cathepsins B, D, and G was confirmed by RT-qPCR. Consistent with western blotting results, RT-qPCR demonstrated greater transcript expression of cathepsins B and D, compared with cathepsin G.
Cysteine cathepsins have been implicated in the pathogenesis of atherosclerosis, abdominal aortic aneurysm and associated complications, via their role in vascular cell apoptosis, foam cell formation, gene expression of matrix proteins, and activation of various substances, including proenzymes, latent cytokines, chemokines, and growth factors. 28 Cysteine cathepsins also participate in arterial remodeling in atherosclerosis, 29 and their potent elastolytic activity renders them key contributors to aneurysmal development. 30 Several studies have demonstrated that expression of cathepsins B, H, K, and L in tumor vasculature is associated with increased angiogenesis. 30
Recent reports demonstrate the expression of cathepsins B, D, and G by the ESC-like population in IH 19 and the ESC-like population on the microvessel endothelium in keloid lesion 24 and Dupuytren's disease. 25
We have recently demonstrated an ESC-like population within mLM that expresses the ESC markers OCT4, SOX2, c-MYC, KLF4, and NANOG. 12 Immunofluorescence staining demonstrated expression of cathepsins B and D by the OCT4+ and the c-MYC+ ESC-like population on the endothelium of lesional vessels and cells within the stroma in the same mLM tissue samples from the same 11 patients we have previously studied. 12 Cathepsin G is known to be expressed by mast cells 31 which also express tryptase. 19 In this study, we showed the expression of cathepsin G by the tryptase+ mast cells that also expressed OCT4, within the stroma of mLM, suggesting a primitive phenotype.
There are existing inhibitors of cathepsin B,32–34 cathepsin D,35,36 and cathepsin G37,38 with inhibitors of cathepsin B currently in clinical use. The expression of cathepsins B and D by the ESC-like population on the endothelium of lesional vessels and cells within the stroma and primitive mast cells within mLM suggests these primitive cells may be a novel therapeutic target by using inhibitors of these cathepsins.
Components of the RAS have been identified in the ESC-like populations in IH, 16 pyogenic granuloma, 17 venous malformation, 14 and fibrotic conditions.39,40 The RAS has been shown to play a role in stem cell proliferation and differentiation41,42 and tumor-associated stem cell growth and spread through remodeling of the tumor microenvironment.13,43
In the classical RAS, angiotensinogen is physiologically released from the liver and then cleaved in the circulation by the enzyme renin, released from the juxtaglomerular apparatus of the kidney, to produce angiotensin I (ATI). 18 ATI is cleaved by angiotensin converting enzyme to form angiotensin II (ATII), which acts upon AT1R and AT2R. 18 Cathepsins B, D, and G form bypass loops for the RAS. 20 Cathepsin B, a lysosomal cysteine protease, converts prorenin into active renin. Cathepsin D, an aspartic lysosomal protease functionally homologous to renin, converts angiotensinogen to ATI. Cathepsin G, a serine protease, produces ATII from ATI and also directly from angiotensinogen.13,19,44 As cathepsins B, D, and G constitute bypass loops for the RAS without activating the classical RAS cascade, 13 and their expression has been observed in IH, 19 investigation into the expression of the RAS in mLM, would be worthwhile.
Limitations of this study include a relatively small sample size. Future studies with a larger sample size and investigation of components of the RAS in mLM, and in vitro and in vivo functional work, are required to determine the precise role of these cathepsins within mLM.
Footnotes
Acknowledgments
We thank Ms. Liz Jones of the Gillies McIndoe Research Institute for her assistance in immunohistochemical and immunofluorescence staining.
Ethics Statement
This study was carried out with the approval of the Central Health and Disability Ethics Committee (Ref. 13/CEN/130) in accordance with the Declaration of Helsinki and written informed consent from all participants.
Authors' Contributions
T.I. and S.T.T. formulated the study hypothesis and designed the study. S.M., N.B., H.D.B., S.T.T., and T.I. interpreted the immunohistochemical data. S.M., N.B., T.I., and S.T.T. interpreted the immunofluorescence data. B.v.S. carried out the RT-qPCR analysis and B.v.S., T.I., and S.T.T. interpreted the results. N.B. conducted the western blot analysis and N.B., T.I., and S.T.T. interpreted the results. K.B. conducted the enzymatic activity assays and K.B., S.T.T., and T.I. interpreted the results. S.M. and S.T.T. drafted the article and all authors commented and approved the article.
Author Disclosure Statement
T.I., P.F.D., and S.T.T. are inventors of a provisional patent Treatment of Vascular Anomalies (PCT/NZ2017/050032), 2016. The authors otherwise declare that the research was conducted in the absence of any commercial or financial relationship that could be construed as a potential conflict of interest.
Funding Information
This research did not receive any specific grant from funding agencies in the public, commercial, or not-for-profit sectors.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.