
Abstract
Background:
Expressed by endothelial cells, CDH5 is a cadherin involved in vascular morphogenesis and in the maintenance of vascular integrity and lymphatic function. The main purpose of our study was to identify distinct variants of the CDH5 gene that could be associated with lymphatic malformations and predisposition for lymphedema.
Methods and Results:
We performed Next Generation Sequencing of the CDH5 gene in 235 Italian patients diagnosed with lymphedema but who tested negative for variants in known lymphedema genes. We detected six different variants in CDH5 five missense and one nonsense. We also tested available family members of the probands. For family members who carried the same variant as the proband, we performed lymphoscintigraphy to detect any lymphatic system abnormalities. Variants were modeled in silico. The results showed that CDH5 variants may contribute to the onset of lymphedema, although further in vitro studies are needed to confirm this hypothesis.
Conclusions:
Based on our findings, we propose CDH5 as a new gene that could be screened in patients with lymphedema to gather additional evidence.
Introduction
The Cadherin 5 (CDH5) gene, also known as vascular endothelial cadherin, encodes a cadherin protein of the cadherin superfamily. 1 Cadherins are responsible for intercellular adhesion and interactions. They are important for the functional and structural organization of cells in many tissues, 2 playing a role in cell growth, differentiation, and migration. 3
All cadherin superfamily members show significant homology and similar domain structures. Although various cadherins are expressed predominantly in different types of tissues, one cadherin type can be expressed in many tissues and one cell type can express more than one cadherin. 4 The only exception so far is CDH5, which is only expressed in endothelial cells.
Endothelial cells form the inner lining of blood, and lymphatic vessels are indispensable for intracellular junction formation. 5 CDH5 is the major adhesion molecule for endothelial cells, being involved in vascular morphogenesis through intracellular signaling and contributing to the maintenance of vascular integrity.5,6 The role of CDH5 in the vascular system has been highlighted in many in vivo studies. Inhibition of Cdh5 by antibody injection loosens blood vascular junctions, resulting in increased permeability and neutrophil extravasation in mice.2,7 Deletion of Cdh5 results in developmental arrest around E9.5 due to massive vascular malformations. 8 An inducible Cdh5 deletion in a mouse model led to increased vascular permeability. 9
The effect of Cdh5 deletion in mouse lymphatic endothelial cells was studied by Hägerling et al., 10 who found that cadherin 5, a major endothelial adhesion molecule, is crucial for the development and maintenance of lymphatic vessels. Although Cdh5−/− embryos formed a primitive network of lymphatic vessels, these failed to mature, leading to massive edema and premature death of the embryos. 10 These results established the importance of Cdh5 in embryo development. Introducing deletion of the Cdh5 gene postnatally, the same authors showed that Cdh5 requirements vary at different stages of life and in different lymphatic vessel types. While dermal lymphatic vessels were susceptible to Cdh5 deletion early in life, in later stages loss of Cdh5 did not affect the phenotype of mouse dermal lymphatic vessels. In contrast, mesenteric lymphatic vessels were sensitive to loss of Cdh5 throughout life. 10 The study demonstrated the consequences of inappropriate Cdh5 expression for lymphatic system development during embryogenesis and later in life.
Cdh5 also has a role in the maturation of lymphatic valves, as showed by Yang et al. 11 Lymphatic valves are important for the maintenance of forward lymph flow. They are formed and maintained constantly throughout life. 11 Cdh5 regulates lymphatic valve development by binding to β-catenin. β-Catenin is known to be necessary for proper lymphatic valve development during embryogenesis, but it can undergo degradation in the cytosol. 12 CDH5 binds β-catenin, giving it stability and preventing it from degradation. 11 CDH5 also binds vascular endothelial growth factors 2 and 3 (VEGFR2 and VEGFR3) and mediates AKT cell signaling. Dysfunction of lymphatic system has been shown not only in lymphedema but also in several other conditions such as obesity, cardiovascular disease, atherosclerosis, Crohn's Disease, inflammation, hypertension, Alzheimer's, and glaucoma.13,14 Lymphedema can be described as the accumulation of interstitial fluid in tissues. This happens due to malformations in the lymphatic system, 15 such as lymph node aplasia, inappropriate lymphatic vessel development, and excessive lymphangiogenesis. 12 Lymphatic valve dysfunction characterized by variants in at least six genes (FOXC2,12,16 FAT4,17,18 GJC2, 16 GATA2, 19 ITGA9, 20 PIEZO1 21 ) has also been implicated in the development of lymphedema.12,15
Taken together, all this evidence indicates that CDH5 plays an important role in lymphatic vessels and lymphatic valve maturation and that its dysregulation may be associated with lymphatic disorders such as lymphedema.
In this study, our aim was to screen variants in CDH5 using Next Generation Sequencing (NGS) in a cohort of 235 lymphedema patients, who previously tested negative for variants in 29 known lymphedema genes. 16 We performed segregation analysis to evaluate genotype–phenotype correlations between CDH5 variants and the development of lymphedema. To evaluate subclinical phenotypes, lymphoscintigraphy was performed in available family members found to carry CDH5 variants. In silico modeling helped to shed light on the possible role of the identified variants in CDH5 protein structure.
Materials and Methods
Clinical evaluation and genetic analysis
We retrospectively enrolled 246 Caucasian patients diagnosed with lymphedema in hospitals across Italy. No consanguinity was reported among their families. Clinical diagnosis of lymphedema was made according to generally accepted criteria. Genetic testing was performed on germline DNA extracted from probands' saliva or peripheral blood. This study was conducted in accordance with the Declaration of Helsinki and was approved by the Ethics Committee of Azienda Sanitaria dell'Alto Adige, Italy (Approval No. 94-2016).
A custom-made oligonucleotide probe library was designed to capture all coding exons and flanking exon/intron boundaries (±15 bp) of 29 genes known to be associated with lymphedema. 22 DNA from probands who tested negative for variants in known genes associated with lymphedema was analyzed for genetic variants in the candidate gene CDH5 (OMIM 601120) that we added to our NGS targeted panel. Identified variants were confirmed by bidirectional Sanger sequencing on a CEQ8800 Sequencer (Beckman Coulter, CA).
We searched the international databases dbSNP and Human Gene Mutation Database professional (Qiagen, CA) for all nucleotide changes. In silico evaluation of the pathogenicity of sequence changes in CDH5 was performed using the Variant Effect Predictor tool 23 and MutationTaster. 24 Minor allele frequencies were checked in the Genome Aggregation Database (gnomAD). 25 All variants were evaluated according to American College of Medical Genetics and Genomics guidelines. 26 Detailed pretest genetic counseling was provided to all subjects, who were then invited to sign informed consent to use of their anonymized genetic results for research.
In silico analysis
The primary amino acid sequences of CDH5 in FASTA format (Table 2) were used as targets to search the Swiss-Model template library (SMTL)—version 2019-10-24—and Protein Data Bank (PDB)—release 2019-10-18 27 —for matching evolution-related structures by means of BLAST (Basic Local Alignment Search Tool) 28 and HHBlits. 29 Models were based on target-template alignment using ProMod3 of the Swiss-Model server. 30 Coordinates conserved between the target and the template were copied from the template to the model. Insertions and deletions were remodeled using a fragment library. Side chains were then rebuilt. Finally, the geometry of the resulting model was regularized with the CHARMM27 force field. 31 In the case of failure of loop modeling with ProMod3, an alternative model was built with ProMod-II. 32 Global and per-residue model quality was assessed using the QMEAN scoring function. 33 BioVia Discovery Studio Visualizer ver17.2 was used to visualize the modeled protein, to vary the targeted amino acids and to analyze interactions at molecular level. 34
Results
Clinical and genetic evaluation
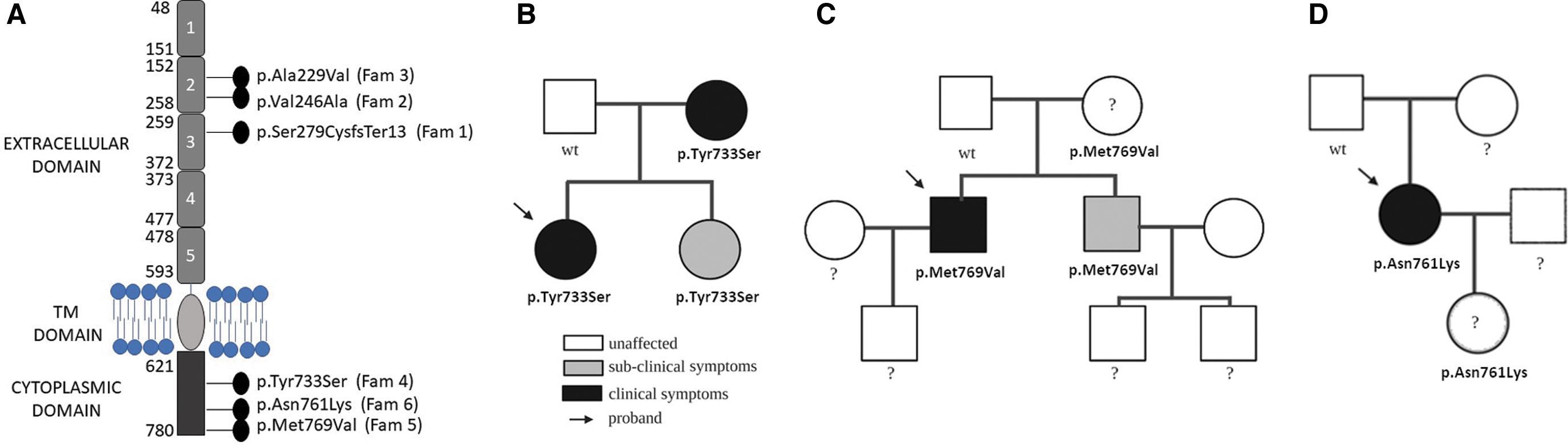
Our cohort for the study of the CDH5 gene consisted of 235 Italian patients with lymphedema. These patients previously tested negative for variants in 29 known lymphedema genes. We found CDH5 variants in six samples, all heterozygous (Fig. 1). The clinical features of the probands are summarized in Table 1.
Clinical Features of the Six Probands and Details of the Cadherin 5 Variants Identified, Showing Sex, Age at Onset (Years), and Genotype
F, female; Fam, family; gnomAD, Genome Aggregation Database; het, heterozygous variant; M, male; NA, not available; VUS, variant with unknown significance accordingly to ACGM classification.
The first proband (female, 60 years) was diagnosed with edema of the lower limbs at age 10. The case reported no other family members with lymphedema. No other family members were available for testing. The proband carries a frameshift variant due to a CTCT deletion that leads to a shorter protein (292 amino acids vs. 785 amino acids of the wild type, Fig. 1B). The variant, classified as VUS (variant with unknown significance accordingly to American College of Medical Genetics and Genomics [ACGM] classification), is not listed in dbSNP, and its allele frequency reported in a cohort of healthy control is 0.000014.
In the second family, the proband (female, 40 years) was diagnosed with lymphedema of the lower limbs at age 28. She carries a single nucleotide missense variant leading to a Val–Ala substitution at residue 246. No other family members were reported affected by lymphedema, and none was available for testing. This variant is not listed in dbSNP or reported in gnomAD.
The third proband (female, 42 years) has a form of lymphedema affecting the left lower limb, diagnosed at age 10. No other family members were tested. The proband carries a missense variant, c.686C>T, with a reported frequency of 0.00000797 (GnomAD). The variant is listed in dbSNP (rs756929820).
In the fourth family, the proband (female, 35 years) was diagnosed with bilateral lymphedema extending from knee to foot (more evident on the left side, Fig. 2A) at age 10, when the menarche appeared. She was found to carry the c.2198A>C heterozygous missense variant, which is classified as VUS by ACGM. Its dbSNP ID is rs900821583, and the GnomAD allele frequency is 0.00000405. This is a familial case, where at the time of genetic testing the proband and her mother both suffered from lymphedema. The mother manifested lymphedema since menopause. She presented bilateral lymphedema from ankles to thighs that can be treated with manual lymphatic drainage. Genetic analysis identified the variant in the mother, as expected, but also in the proband's younger sister (Fig. 1B). The sister (age 19) was considered to be healthy; however, lymphoscintigraphy showed evidence of lymphatic dysfunction (Fig. 3A) that progressed in a year to minor edema in both legs. She also showed lipedema, like her older sister, at the thighs. Her treatment consists in manual lymphatic drainage, too.
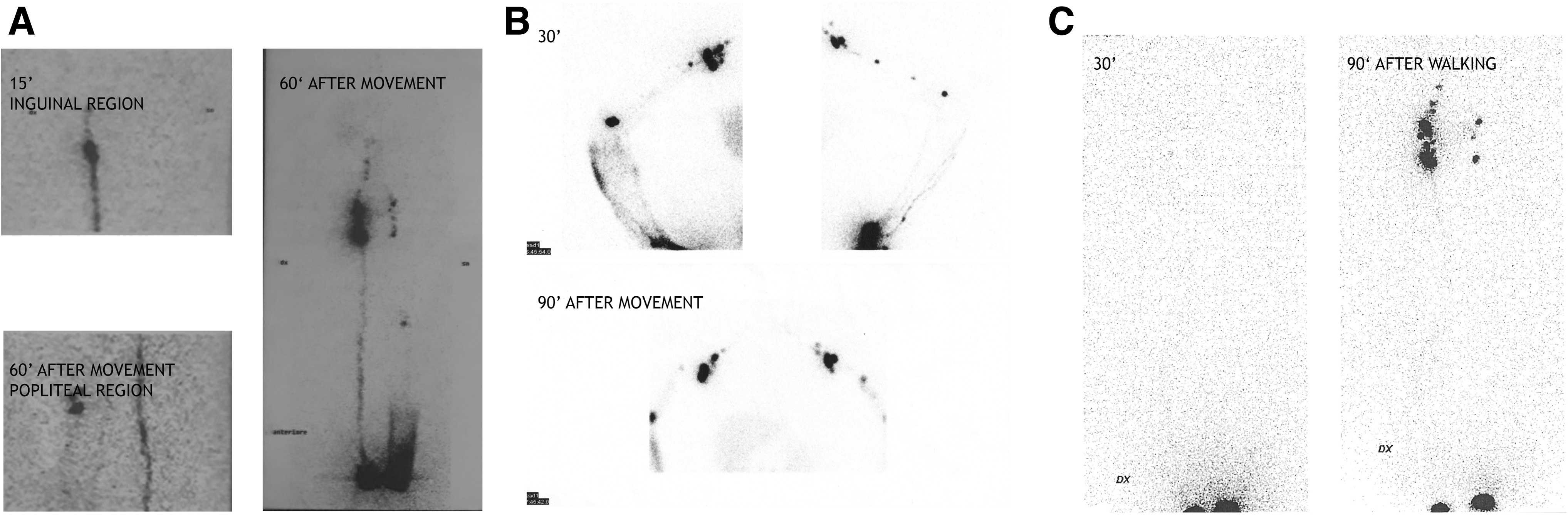
Lymphoscintigraphic scans of probands in family 4

Lymphoscintigraphic scans of
The fifth proband (male, 48 years) has a history of primary lymphedema of the upper right arm and inflammation of the lymphatic vessels since age 35. This patient showed multiple bilateral lymph node stops of the upper limbs with dermal back flow on the right (Fig. 2B). A missense variant in CDH5, leading to a Met769Val variation, was identified. The variant is listed as rs140804041 in dbSNP, and its reported frequency is 0.0000812. The same variant was found in the proband's mother and brother (Fig. 1C): neither of them manifests overt lymphedema, although the brother's lymphoscintigraphy showed impaired lymph transport in both upper limbs and few left axillary lymph nodes. In the lower limbs, reduced uptake by the right inguinal lymph nodes was found (Fig. 3B). Lymphoscintigraphy was not performed in the mother.
The last proband (female, 58 years) has had lymphedema of the left foot since age 56. Lymphoscintigraphy showed reduced left inguinal uptake (Fig. 2C). She carries a missense single nucleotide variant (c.2283C>A), not listed in dbSNP or GnomAD. It is classified in Varsome as VUS. 35 Samples from the mother and daughter, neither of whom have lymphedema, were also analyzed; the daughter was found to carry the variant. Unluckily, we were unable to perform lymphoscintigraphy in the latter subject. The family tree is shown in Figure 1D.
Although Hägerling et al. 10 showed that the intestinal lacteals were dilated on Cdh5 deletion in mouse model, none of our probands reported any symptoms attributable to lymphangiectasia or diarrhea.
In silico analysis, template selection, and model building
Our template search for the CDH5 amino acid sequence (Table 2 in FASTA format), submitted to the Swiss-Model server using BLAST and HHBlits, resulted in a total of 356 templates that matched with different sequence identity and quality percentages. Details of the top 10 templates are given in Table 3.
The Cadherin 5 Amino Acid Sequence in FASTA Format
Top 10 Matching Templates from the Swiss-Model Template Library
Homo, homo-dimer; QSQE, quaternary structure quality estimate.
Based on the percentage of sequence identity, similarity, and best quality square, the 3q2w.1.A chain was selected to align the template and query sequences for model building.
We used the Discovery Studio Visualizer to generate the Ala229Val, Val246Ala, and Met769Val versions of the structure. Due to a lack of structural details, we were unable to model Tyr733Ser and Asn761Lys. Molecular level interaction analysis was performed between native/mutant residues and interacting residues, snapshots of which are shown below (Figs. 4–6). Details of the residues involved in interactions along with the types of bond they formed and bond lengths in Angstrom units are shown in Table 4. The Ser279CysfsTer13 variant was not modeled, since this frameshift variant loses two thirds of the wild-type protein structure and it might undergo nonsense mediated decay, as highlighted also by MutationTaster results.

Molecular interactions of the
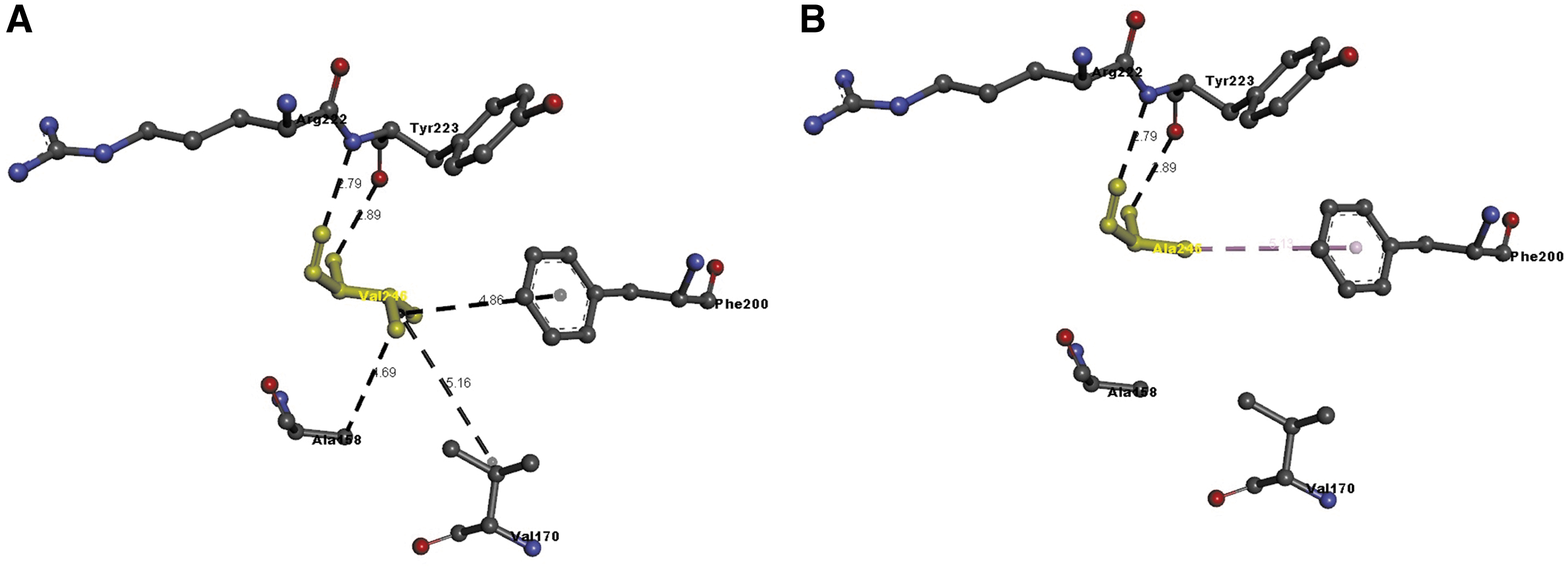
Molecular interactions of the
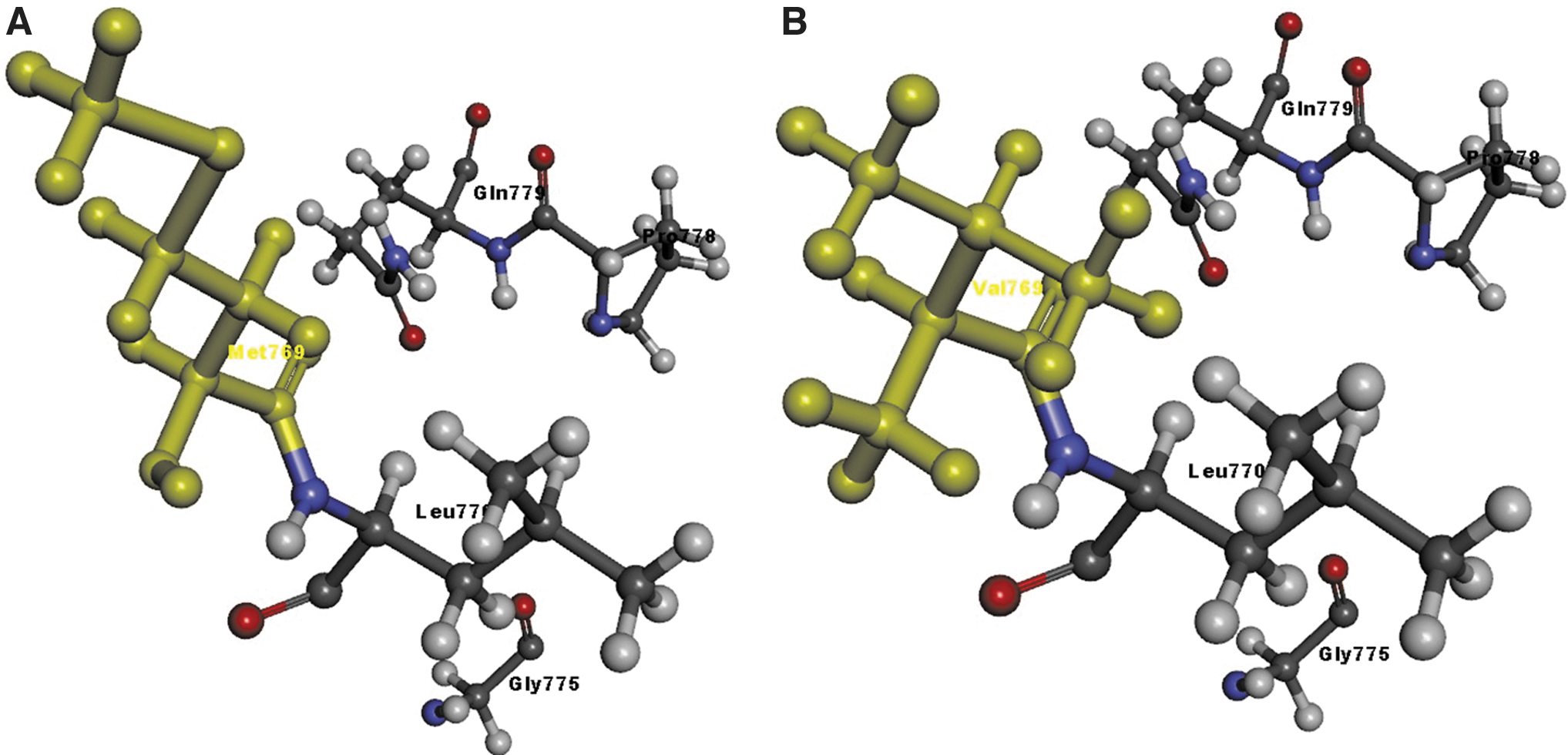
Molecular interactions of the
Details of Molecular Interactions of the Native (Above) and Substituted (Below) Amino Acid Found in Families 2, 3, and 5 with Adjacent Residues in the Modeled Cadherin 5 Protein
The type of interaction and the length in Angstrom units are reported.
In silico analysis showed the following differences. Ala229Val did not differ significantly in the number of bonds formed; however, the analysis showed changes in the length of the bonds between the wild-type amino acid and the one found in the proband (Fig. 4).
Val246Ala showed a significant loss of bonding because the variant sacrificed two bonds with Ala158 and Val170 (Fig. 5).
Met769Val did not form any bonds nor did the native protein. However, the residue in the modeled variant was bulkier (Fig. 6), therefore suggesting a possible conformational change of the protein.
The loss of hydrogen bonds in the case of Val246Ala and the loss of a great portion of the protein if translated in the case of Ser279Cys could be the reason for conformational changes in the overall structure of the protein, resulting in its malfunction. The Asn761Lys variant introduces a change from a negatively charged amino acid to a positively charged one, and the variant Tyr733Ser changes an aromatic residue to a nonaromatic one. Both these substitutions could modify the structure and interaction of the protein; unluckily, we were unable to model the structure of the protein in this region.
Discussion
The genetics of lymphedema are still not entirely clear: the genes that have been associated with lymphedema so far can only explain diagnosis in about one third of cases. To increase the diagnostic sensitivity of lymphedema testing and to achieve a better understanding of the biology of this disorder, new gene associations need to be discovered by increasing the number of genes screened.13,36
The human vascular system consists of specialized vessel types, including arteries, veins, and lymphatic vessels. The lymphatic system mediates fluid homeostasis, immune cell trafficking, and nutrient distribution. CDH5 is essential for a properly functioning lymphatic system, especially for lymphatic vessels and valves. Although attention to CDH5 has mostly been in the context of cardiovascular disorders and cancer,37–39 recently its role in the lymphatic system was also demonstrated. Studies of mouse models clearly demonstrate the importance of Cdh5 in the development of the lymphatic system.10,37 Complete deletion of Cdh5 leads to severe lymphatic vessel malformations, massive edema, and embryo death. The phenotype of postnatal conditional knockout mice shows that expression of Cdh5 depends on the specific type of lymphatic vessel and on mouse age. Conditional deletion also impairs the maturation of lymphatic valves, leading to fluid accumulation and edema.10,11
These findings highlight the possible role of CDH5 in lymphatic system development and maturation, and they sustain the utility of NGS screening for variants in CDH5 in lymphedema patients tested negative for variants in genes clearly associated to this disease.
In this article, we tested DNA samples of 235 lymphedema patients for variants of the CDH5 gene. These patients had previously tested negative for variants in 29 genes known to be involved in the development of lymphedema. Six patients carried a rare CDH5 variant (6/235; 2.5531%). Five probands were female (83.3%) and one male (16.7%). Five carried a heterozygous missense variant, and one variant was a deletion resulting in a frameshift. Family members of three of the six probands were also tested. We could not detect a clinical characteristic that could accumulate all of the patients identified to harbor variants in CDH5; they do not clearly differ from other groups of patients affected by lymphedema with involvement of other genes.
Segregation analysis in family 4 and 5 showed that, although not all members of our cohort who carry a variant in CDH5 have been diagnosed with lymphedema, lymphoscintigraphy in available subjects showed lymphatic system anomalies. This suggests that CDH5 could play a role—yet to be clarified and confirmed by in vitro and in vivo functional studies on the reported variants—with lymphatic malformations or lymphatic system dysfunctions that may or may not lead to lymphedema later in life. Indeed, incomplete penetrance and variable phenotype are often seen in genes associated with lymphedema. 36
To further explore the effects of CDH5 variants on the function of Cadherin 5, we performed bioinformatic analyses that highlighted differences between the interaction of native and mutated residues with the immediate molecular environment. Two reported variants were located in the extracellular domain of Cadherin 5 (i.e., p.Ala229Val in family 3, p.Val246Ala in family 2), while three other variants were reported in the cytoplasmic domain of the protein (i.e., p.Tyr733Ser in family 4, p.Asn761Lys in family 6, and p.Met769Val in family 5). Moreover, p.Ser279CysfsTer13 variant found in family 1, located in the extracellular domain, might lead to a translation of a truncated protein (missing the cytosolic and the transmembrane domain), if it does not undergo RNA-mediated decay. The Cadherin 5 cytosolic tail is involved in the interaction with β-catenin, plakoglobin, and p120; so variants in this region could interfere with binding with these proteins, thus altering communication with the cytoskeleton, essential for proper adhesive function. 8 The CDH5 transmembrane domain is known to mediate binding to VEGFRs 2 and 3, 40 while extracellular domain is mainly involved in adherent junction formation. Therefore, variants in this latter domain could lead to dysfunctional establishment and maintenance of cell to cell adhesion, actin cytoskeleton remodeling. 41 Further functional studies are needed to provide insights on the role of CDH5 variants in the lymphatic system.
Conclusions
Data from reviewed literature show that the normal development and maturation of lymphatic endothelial cells, which form the lymphatic system, are critical for its proper function. Although in vitro and in vivo study are needed to confirm the pathogenicity of the variants we identified in lymphedema patients, we propose CDH5 as a candidate gene to be screened in patients with negative genetic testing in lymphedema, aiming at gathering further evidence of its involvement in the predisposition or in the development of lymphedema. Indeed, future study gathering a wider number of patients harboring variant in CDH5 is needed to highlight if this subgroup of patients presents specific clinical or instrumental characteristics.
Footnotes
Acknowledgment
The authors thank Helen Ampt for English language editing.
Authors' Contributions
Conceptualization, M.B.; investigation, S.M., M.R., and R.S.; resources, S.M., M.R., and M.B.; data curation, M.B., M.E., M.D., J.K., S.H.B., S.P., J.P.B., and M.B.; writing—original draft preparation, S.K. and D.V.; writing—review and editing all coauthors; visualization, M.B., E.M., S.H.B., S.P.; supervision M.B.; project administration, M.B. All authors have read and agreed to the published version of the article.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by fundings from the Provincia Autonoma di Trento in the framework of the initiative LP 6/99 (dgp 1045/2017).