
Abstract
The lymphatic system was first described in the 17th century independently by Olaus Rudbeck and Thomas Bartholin. Since then, there has been deep-seated fascination with its development, function, and dysfunction.
Olaus Rudbeck and Thomas Bartholin each (independently) initially described the lymphatic system in the 17th century. 1 Since then, there has been deep-seated fascination with its development, function, and dysfunction.
The functional role of the lymphatic circulation has been defined as follows:
Removal of excess fluids from body tissues, a crucial process due to the continuous leak of water, proteins, and other substances out of blood capillaries into surrounding body tissues. An inability to drain excess fluid from the interstitium results in buildup of lymphatic fluid within the body's tissues. Absorption of fatty acids and subsequent transport of fat, chyle, from the intestinal tract to the circulatory system. An important role in the maturation and specialization of immune cells (such as lymphocytes, monocytes, and antibody-producing cells called plasma cells).
Familiarity with the embryology of the lymphatic system is helpful in understanding the pathogenesis and subsequently, the necessary radiological approaches to lymphatic abnormalities. Molecular and structural insights recently identified have clarified some of the embryological origins of the lymphatic vasculature. By considering anatomic location and radiological appearance, one can reasonably predict the type of lymphatic anomaly present, the primordial lymph sac from which the malformation arose, and when it formed in embryonic life.
However, along with the ever-increasing understanding of the lymphatic system, there comes an acknowledgment that the lymphatic system is extremely complex with a wide range of pathology and pathophysiology. 2 Therefore, although significant strides have been made, we are nowhere near translating these understandings to clinical therapies, and certainly not curative approaches that some of these patients desperately seek.
A few points are recently substantiated that help in our understanding of lymphangiogenesis.
Lymphatic vessels develop later than the blood vascular structures, at least 4 weeks after the appearance of the first blood vessels. 3
Lymphatic vessels arise from endothelial cell aggregates through combined forces of vasculogenesis and angiogenesis. Jugular lymph sacs are the earliest identifiable embryonic lymphatic precursors. These paired embryonic structures are to be found in apposition to the jugular aspect of the cardinal vein (CV).
Normal lymphatic vascular development likely entails elements of both of the proposed models, namely, the effects of both centrifugal and centripetal developmental forces.
Lymphangiogenesis is divided into four distinctly identifiable stages: competence, commitment, specification, and coalescence and maturation.
The centrifugal theory proposes that lymph sacs are the earliest anlage of the lymphatic system. These lymph sacs arise from central embryonic veins and give rise to the entire lymphatic vasculature of the body by a process of centrifugal sprouting. The alternative centripetal theory proposes that lymphatic vascular precursors arise independent of venous structures and subsequently fuse to form lymphatic vessels.
Molecular insights have supported both models to some extent. One, noted by Wigle et al., supports the centrifugal theory of development, as identified by the expression pattern of the transcription factor Prox1 during murine embryonic development. 4 Another by Francois et al. noted that the Sox18 transcription factor drives Prox1 expression through a subset of venous endothelial cells. These respond to vascular endothelial growth factor (VEGF)-C and bud from the CV to form lymph sacs. 5
Improved insights into these processes have emanated from the elucidation of the inherent molecular mechanisms of lymphatic vascular development.
The earliest step in de novo mammalian lymphatic development is characterized by the detection of Prox1 expression in a subpopulation of blood endothelial cells within the CV. This occurs at a murine embryonal age approximating 9.5 weeks. These Prox1-positive lymphatic endothelial cells (LECs) leave the CV and contribute to the formation of lymph sacs. This cellular event entails active budding, in which the progenitor LECs are characterized by intercellular junctions that express VE-cadherin. These progenitors are present both in the CV and in the intersomitic vessels. As the LEC progenitors penetrate the neighboring mesenchyme, they begin to express podoplanin (PDPN). Through the investigation of Prox1-null embryos, it has been established that Prox1 expression is an obligatory molecular constituent of this migratory process. 6
As lymphatic development continues, the lymph sacs are the first true lymphatic structures noted. As previously stated, the first lymph sacs that can be identified are located at the junction of the internal jugular vein with the subclavian vein.
The unpaired retroperitoneal lymph sac at the root of the mesentery of the intestine subsequently appears. This structure develops from the primitive IVC and mesonephric veins.
The paired posterior lymph sacs develop from the iliac veins and produce capillary plexi and the lymphatic vessels of the abdominal wall, pelvic region, and lower limbs.
Excepting the anterior part of the lymph sac from which the cisterna develops, all lymph sacs eventually are invaded by mesenchymal cells. These develop into groups of lymph nodes.
Mesenchymal cells between layers of the dorsal mesentery of the stomach form the spleen.
In the current prevailing models, vasculogenesis of the lymphatic circulation requires four distinct stages, namely, lymphatic competence, commitment, specification, and vascular coalescence and maturation.
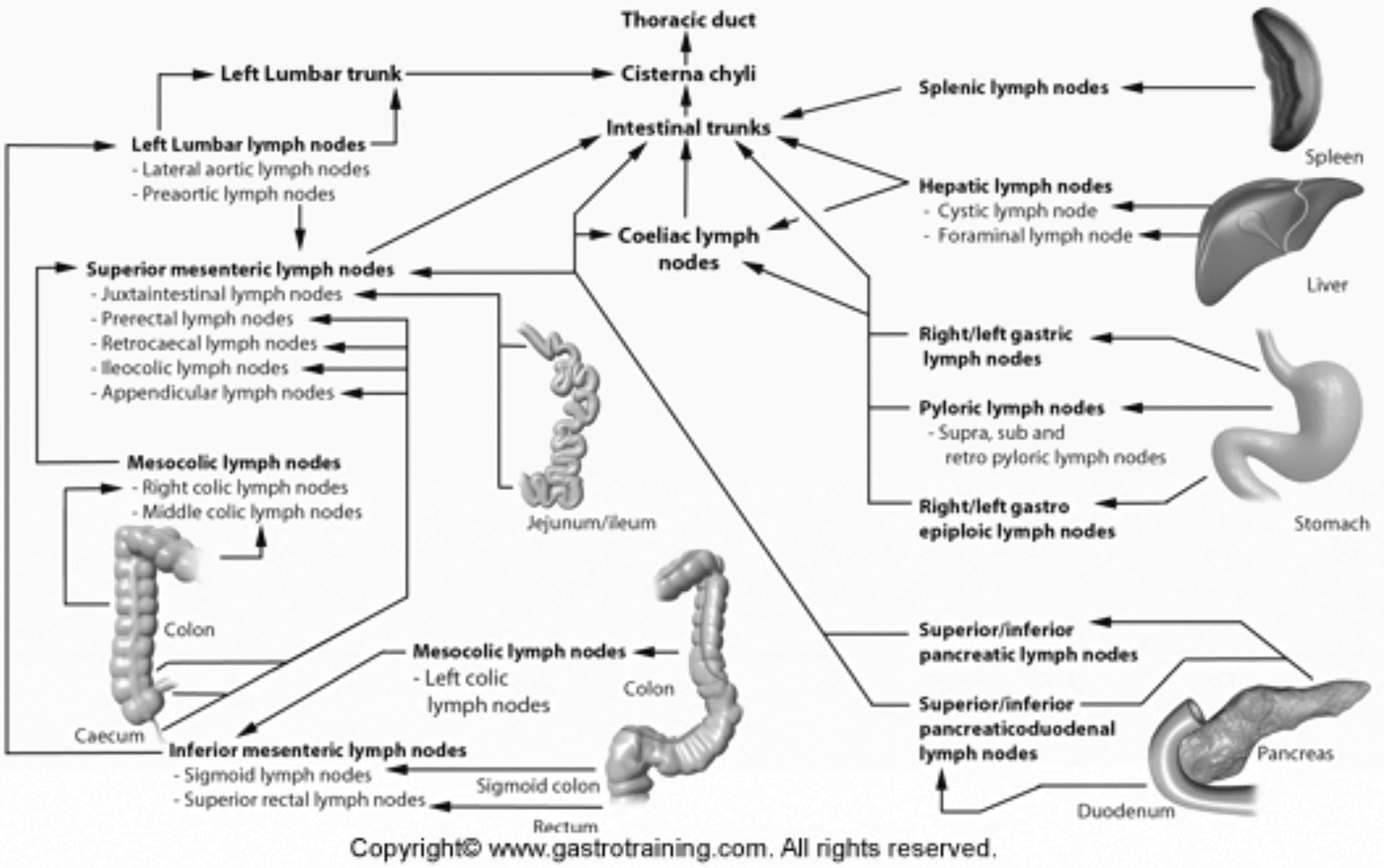
Overview of the lymphatic drainage of abdominal and pelvic organs. Reprinted from Ref. 45 with permission.
Lymphatic Competence
Lymphatic competence describes the ability of the cells to respond to the initial signals that induce lymphatic vascular differentiation. 7 The priming of LECs to initiate lymphatic development appears to be dependent on molecular signaling pathways that are distinct from those related to blood vascular development.8,9
The competence of LECs is recognized through cellular expression of the lymphatic vessel endothelial hyaluronan receptor-1 (LYVE1)3,10 and vascular endothelial growth factor receptor-3 (VEGFR3). The flt4 gene encodes for VEGFR3. 11 The interaction of VEGF-C ligand with its obligate VEGFR3 receptor promotes LEC proliferation, migration, and survival.12,13 In prior studies, mouse embryos lacking VEGFR-3 expression are lethal in the absence of lymphatic development. Mouse embryos lacking VEGF-C have been found to not develop lymph sacs. 14 It is safe to say that the VEGF-C/VEGFR3 pathway exerts multiple critical influences on lymphatic vascular development. Although VEGF-C signals can be transduced by a variety of biologically active receptors, binding to VEGFR3 is central and critical. 15
Lymphatic Commitment
Lymphatic commitment entails the necessary expression of Prox1, found only in cells that possess a committed lymphatic lineage. 3 With the appearance Prox1 expression, a venous endothelial cell sheds its blood vascular commitment in favor of that of lymphatic lineage. 4 This embryonic endothelial cell Prox1 expression is not only necessary for the process of lymphatic commitment, it is, in fact, independently sufficient. As these Prox1-positive LECs bud and migrate away from the cardinal and intersomitic veins, the streams of interconnected cells 16 initially aggregate into a lymphatic plexus that serves as the source for the subsequently formed lymph sacs.6,17,18
LEC Specification
LEC specification is characterized by the expression of unique molecular markers that, in aggregate, contribute to the final function and morphology of LECs. As these cells migrate from the parent veins, their identity is identifiable through their expression of lymphatic-specific markers, including PCPN, neuropilin-2, and VEGFR3.6,17,18 This lymphatic specification helps to establish autonomy from the parent venous structures. Primary lymph sacs form throughout the embryo, and ultimate phase of lymphatic development is characterized by secondary budding and migration. In aggregate, the lymphatic vasculature comes into being through this centrifugal formation of capillary structures. 11
Vascular separation of blood and lymph is critical to normal lymphatic vascular development. This important event is mediated through the expression of Syk, a tyrosine kinase, and Slp-76, an adapter protein. If either of these proteins is not properly expressed, there are abnormal blood-lymph connections, characterized by blood-filled lymphatics and chylous hemorrhage. 19 Normal embryonic development entails platelet aggregation at the loci of lymphaticovenous connection. Platelet aggregation commences through the binding of LEC-expressed PDPN to platelet C-type lectin receptor 2. This interaction activates Syk and Slp-76.20,21 Murine models have documented the presence of blood-filled lymphatics in response to inadequate megakaryocytes, platelets, C-type lectin receptor 2, PDPN, Syk, or Slp-76.16,19–26
Vascular Coalescence and Maturation
The next stage of embryonic lymphatic vascular development entails continued maturation and remodeling of the nascent structures. A hallmark of this stage of development is the appearance of the lymphatic valve apparatus. FOXC2, A forkhead transcription factor that is highly expressed in the mature lymphatic valve apparently specifies the collecting lymphatic vessel phenotype.27,28 Expression of GATA2 is also necessary for normal valvular development. 29 In mice, the absence of GATA2 expression produces defective valve structure and morphologically abnormal, distended collecting vessels. Expression of integrin-alpha9 with extracellular matrix deposition of its ligand, fibronectin-EIIIA, is also a requisite participant in valvular development. 30
Speaking to their collective importance, GATA2, FOXC2, and CCBE1 have all been described to be associated with lymphedema (ISSVA classification, 2018). GATA2 mutations cause lymphedema and myelodysplasia, FOXC2 lymphedema-distichiasis and CCBE1 mutations have both been identified in primary generalized lymphatic anomaly. Therefore, an increased understanding of lymphatic development may offer clear clinical implications for diagnosis, diagnostic imaging, and management. Valvular development is supported by additional signaling pathways, including bone morphogenetic protein (BMP), 31 Notch, 32 and semaphorin3a-neuropilin-1. 33
Lymphatic maturation also relies upon the expression of ephrins and the angiopoietins. In mice, insufficient eEphrinB2 expression promotes deficient valve formation, collecting lymphatic hyperplasia, and failure of lymphatic capillary remodeling. 34 EphB4 forward signaling also has a role in the development of the lymphatic valves 35 ; Angiopoietin-1 and -2 (Ang1 and Ang2) are required for the maturation of the embryonic lymphatics.36–38 In these vessels, in contradistinction to the blood vasculature, Ang2 is an agonist for theTie2 receptor. 37 Murine deletion of Ang2 produces lymphatic vessels that recruit smooth muscle cells prematurely into the vascular wall, and they fail to develop valves.36,37 Ang2 was recently shown to have high levels in the blood of patients with kaposiform lymphangiomatosis and has been shown to be useful as a biomarker of disease activity. 39 The Tie1 receptor participates in early lymphatic development.40,41 Reelin signaling also promotes lymphatic vascular maturation, regulating smooth muscle incorporation into the wall of the lymphatic collectors. 42
Clearly, lymphatic developmental biology is complex, and the current list of molecular participants is very likely to be incomplete. With continued delineation of the participating molecular events, it should be anticipated that identification of these signaling mediators may provide targets for future molecular therapeutics in lymphedema and other diseases of the lymphatic circulation.43,44
As the lymphatic network develops, it takes on a discontiguous developmental pattern that results in the differential imaging approach that is currently undertaken in the workup of lymphatic disorders.
The lower extremity lymphatics flow centrally, through the inguinal lymph nodes, into the posterior pelvic lymphatic channels. These then form the larger cisterna chyli, which is usually in the upper abdomen. The cisterna chyli continues in the thorax as the thoracic duct, once it has accepted flow from the mesenteric lymphatics. As such, imaging with nuclear lymphoscintigraphy, MR lymphangiography, or lipiodol-based lymphangiography are supportive techniques that determine integrity of the lower extremity and central lymphatic channels.
The mesenteric and liver lymphatics are not visualized with the current lower extremity techniques. This lymphatic system separately flows from the liver and intestines, into the mesentery, and then drain into the cisterna chyli. Of note, the uptake of these lipid and fat-rich materials results in a change in the appearance of the simple/serous lymphatic fluid to a more milky whiter chylous fluid. This appearance of the fluid traversing the lymphatic channels can also help localize the level of injury to the lymphatic system. The following diagram shows the complexity of abdominopelvic lymphatic drainage and easily explains why any maldevelopment can have serious deleterious effects.
In the upper extremities, the lymphatic drainage is also not seen through scintigraphy from the lower extremity, and thus requires separate radiotracer injection in the upper extremity to visualize the outflow. In recent years, the use of upper extremity MR lymphangiography has gained acceptance although its use remains limited due to the resource-intensive nature of the examinations. 46
Only with aggregation of different imaging techniques can the entire lymphatic system be radiographically visualized. However, these differing imaging techniques can offer the location of injury and point to potential therapeutic options.
Key Terms
Anlagen: the rudimentary basis of a particular organ or other part, especially in an embryo.
Mesoderm: one of the three embryonic tissue layers. During embryonic development, it is the source of the adult internal organs, including the muscles, the spine, and the circulatory system.
Cisterna chyli: the lymph from the intestinal trunk and the two lumbar lymphatic trunks empties into this dilated sac at the inferior aspect of the thoracic duct.
Lymph node: small ovoid components of the lymphatic system that are distributed along the length of the lymphatic vessels. They cluster in coalescent regions (e.g., armpits and groin). Their internal honeycomb of connective tissue serves as a filter containing lymphocytes and macrophages that process bacteria, viruses, and antigenic proteins in lymph.
Lymph sac: embryonic precursors of the lymph vessels.
Footnotes
Author Disclosure Statement
No competing financial interests exist.