
Abstract
It is known that nitric oxide (NO) is a gas and synthesized from
Introduction
Currently, new lymphology combined with cardiovascular medicine, immunology, oncology, and its basic sciences is proposed by the members in Japanese Society of Lymphology. 1 In this review, as one example of the new lymphology, it is clarified that the lymph flow plays a key role in lymphatic active pump activity in vivo through the release of nitric oxide (NO). In addition, the lymph flow not only stimulates the NO release but also increases the molecular expression of endothelial constitutive NO synthase (NOS) (ecNOS) mRNA and the protein in lymphatic endothelial cells (LEC).
The lymphatic system supports an important homeostatic function with returning fluid and plasma proteins from the interstitial tissues through thoracic duct to subclavian vein. The lymphatic system has also been recognized as both a metastatic route of carcinoma cells and an immune surveillance system. The lymphatic function depends on lymph production in lymphatic capillaries, the lymph transport, and the physiological and immunological properties of lymph nodes.2–6 The lymph transport depends on passive such as respiratory movement, arterial pulsation, and skeletal exercise, and active driving forces. The intrinsic contractility of lymphatic smooth muscles produces heart-like spontaneous contractions in mesenteric lymph vessels, which play a key role in the active driving force, resulting in centripetal propulsion of lymph.7–9
The rhythm and amplitude of the spontaneous contractions are regulated by neural and vasoactive agents included with NO.10–18 Numerous studies of endothelium-dependent relaxation in blood and lymph vessels have been reported since original description by Furchgott and Zawadzki. 19 The endothelium-derived relaxing factor (EDRF) was released from endothelial cells by vasoactive agents such as acetylcholine (ACh) or shear stress stimulation. In 1987, Palmer et al. 20 demonstrated that EDRF was actually NO. Since that time, the research regarding NO has surged in almost every discipline of biomedical sciences. In the lymphatic system, NO plays a marked vasodilator that acts at the Ca2+-dependent contraction pathway.21–32 In addition, it is known that the volume of lymph node is regulated through the activation of NO release, which may contribute to help for the proliferation of lymphocyte-mediated increase in the volume. Thus, 5-hydroxytryptamine produces the NO-dependent relaxation of the smooth muscles of the capsule in monkey popliteal lymph node. 33
The Biological Properties of NO
NO is a gas and synthesized from
There are three different types of NOS, namely ecNOS, neuronal NOS (nNOS), and cytokine-inducible NOS (iNOS). They exhibit two expression patterns. Thus, the ecNOS and nNOS are expressed constitutively at low levels. They can be activated rapidly by an increase in cytoplasmic calcium ions (Ca2+). In contrast, the iNOS is induced when macrophages are activated by cytokine, but no intracellular increase in Ca2+ is required. The induction of iNOS results in production of larger amounts of NO, which may produce different pathophysiological effects. 35
In contrast, NO produced by the endothelium through the activation of ecNOS can quickly diffuse to vascular smooth muscle cells in the walls. The NO activates cytoplasmic GMP in the smooth muscle cells and then reduces intracellular Ca2+ via the activation of Ca2+ outflux, resulting in vasodilation of blood and lymph vessels. 34
Flow-Mediated Release of NO from LEC of Canine Thoracic Duct
It is known that lymph flow-mediated NO release from the endothelial cells of thoracic ducts produces a marked vasodilation of the lymphatic smooth muscle cells.28,36,37
With the isolated and perfused preparations of canine thoracic duct, it was clearly demonstrated that lymph flow mediated NO-dependent relaxation of smooth muscles in the bioassay canine arterial preparation. 36 Cylindrical segments isolated from canine thoracic ducts were perfused at a constant flow rate ranging from 0.5 to 2.0 mL/min. The NO activity in the perfusion effluent through the lymphatic segments was evaluated by a relaxation level of the endothelium-denuded canine coronary arterial bioassay rings. To keep a patent cross section of the lymphatic segment, a constant transmural pressure (∼5 cmH2O) of the segments was maintained throughout the experiments.
Figure 1 shows representative recordings for the effects of the perfusion effluent through the lymphatic segment, ACh (10−5 M), and sodium nitroprusside (10−5 M) on the mechanical activity in arterial bioassay preparation. The bioassay preparation is perfused with the effluent solution through the thoracic ducts with (Fig. 1A) and without (Fig. 1B) intact endothelium. The effluent solution circulated through the intact endothelium of thoracic duct (Fig. 1A) produced the flow rate-dependent relaxation of the arterial bioassay preparation. The effluent solution through the incubated thoracic duct with 10−5 M ACh also produced an additional relaxation of the bioassay preparation (Fig. 1A).
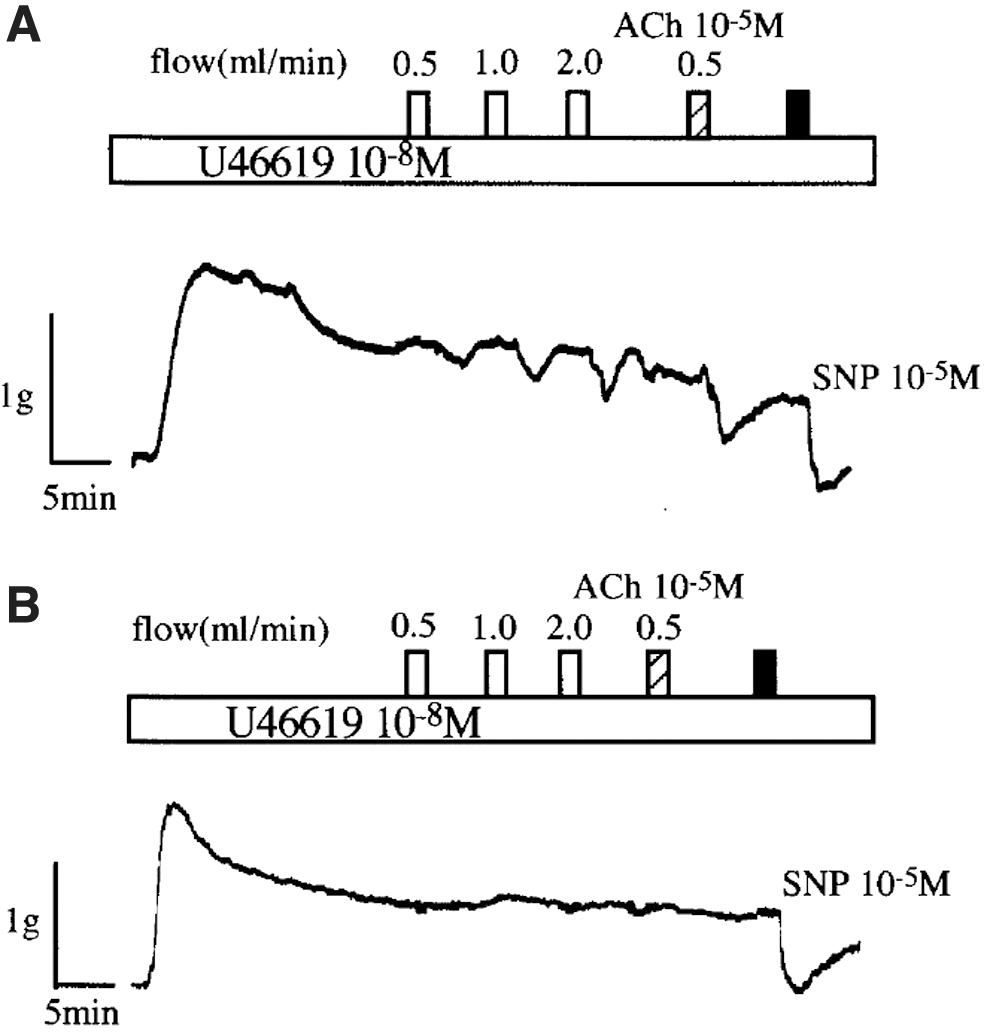
The representative recordings of the effects of the increasing flow ranging from 0.5 to 2.0 mL/min, 10−5 M ACh, and 10−5 M SNP on the mechanical activity of endothelium-denuded arterial bioassay preparations perfused with the effluent solution through canine thoracic ducts with
In contrast, the effluent solution circulated through the thoracic duct without intact endothelium produced no significant reduction of the muscular tone in the bioassay preparation (Fig. 1B). In addition, pretreatment with Nw-nitro-
The NO-Dependent Control of Heart-Like Spontaneous Contraction and Active Lymph Pump in Mesenteric Lymph Vessels
It is well known that the rhythmical heart-like activity of lymph vessels, in corporation with the presence of valves, creates active pump in mesenteric lymph vessels.5,6,9,10,38,39 Thus, the mesenteric lymphatic vessels perform a sump pump for the tissues to propel excess free fluid away from the interstitial space. In all vertebrates, including human beings, sheep, cattle, rat, and mouse, the mesenteric collecting lymph vessels have shown spontaneous rhythmic contractions. The frequency of contractions is determined by the amount of lymph volume-mediated stretch of the lymphatic smooth muscles 5 and regulated by the sympathetic nerve fibers activity. 10 It is also noteworthy that there exist lots of vasa vasorum in bovine mesenteric collecting lymph vessel walls, 40 which may be essential for maintaining the spontaneous contractions of lymphatic smooth muscles because of the lower oxygen tension (25–35 mmHg) in lymph. 6 In contrast, murine peripheral lymph vessels do not develop large amplitude of spontaneous contractions. 41
Lymph Flow-Released NO Inhibits the Spontaneous Contractions in Mesenteric Lymph Vessels
Gashev et al. 15 demonstrated that physiological saline solution perfused intraluminally at a range of transmural pressures from 0 to 7 cmH2O produced markedly negative inotropic and chronotropic effects on the spontaneous contractions in isolated rat mesenteric lymph vessels. The flow-mediated negative responses were significantly reduced by the pretreatment with L-NAME. In addition, ACh also induced negative chronotropic and inotropic effects on regular rhythmic activity in isolated bovine mesenteric lymph vessels. 14 The ACh-induced responses of spontaneous contractions were reduced by the pretreatment with L-NAME.
The additional treatment with
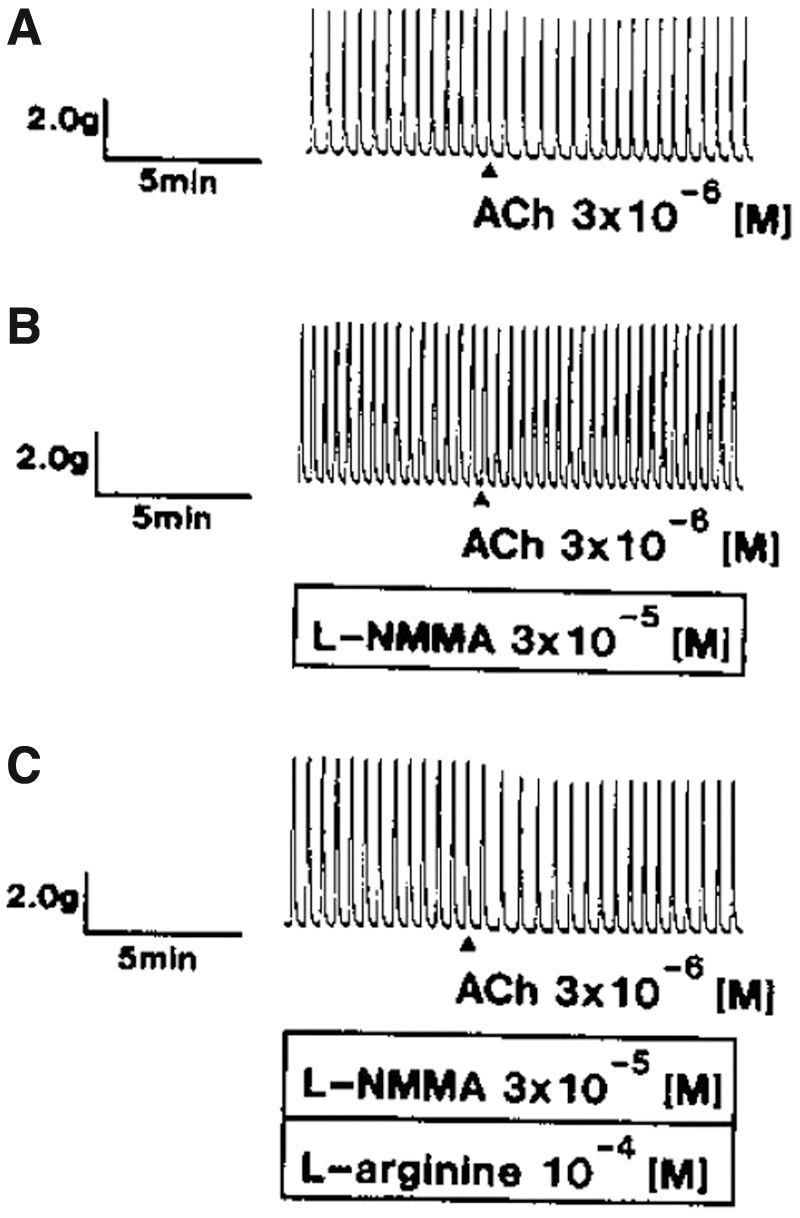
The representative recordings of the effects of L-NAME
Endogenous NO Has Physiologically Modulated the Lymph Active Pump Activity in Rat Mesentery In Vivo
The impressive study 23 has demonstrated the effects of lymph flow on the active pump in in vivo rat mesenteric lymph vessels. Using the intravital video microscopy, the active pump activity in rat jejunal-originated mesenteric lymph vessel was observed in vivo. Figure 3A shows a representative video image of rat mesentery. The collecting lymph vessel (85–95 μm in outer diameter) was observed as a white channel. Figure 3B demonstrates a representative rhythmical change in the diameter of the segment in one lymphangion, a segment of lymph vessel lies between valves. It shows a sequence of nine images recorded every 0.2 seconds at the same area of one lymphangion shown as a white square in Figure 3A. The lymphatic images labeled 5 and 9 are a maximal constriction and dilation in one lymphangion. In control condition, the pumping frequency, end-diastolic diameter, and endo-systolic diameter were 4.5 ± 0.7, 82 ± 6, and 45 ± 3 μm, respectively.
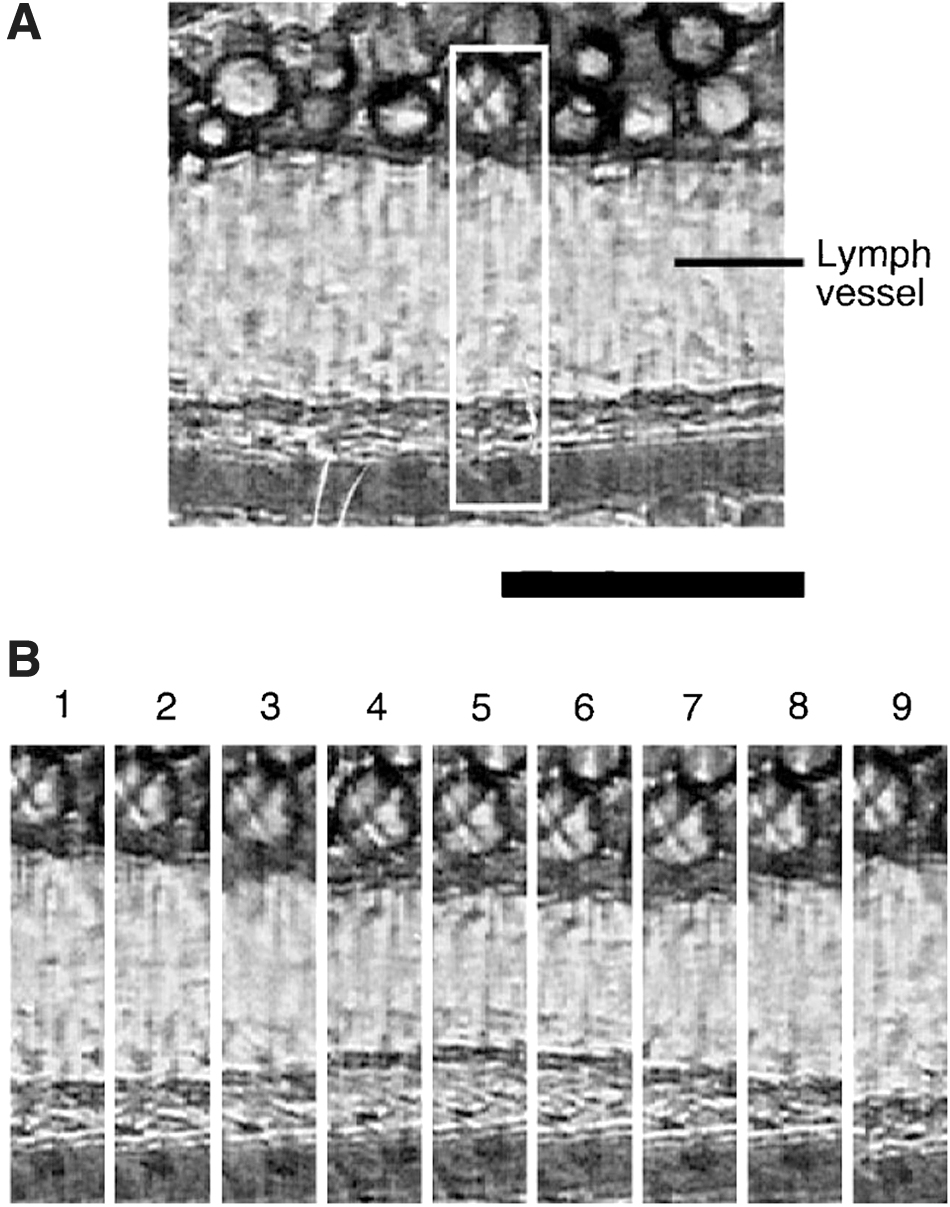
The representative video images of rat mesentery
Figure 4 demonstrates the effects of 30 μM L-NAME on pumping frequency (Fig. 4A) and end-diastolic (Fig. 4B) and endo-systolic diameters (Fig. 4C) of one lymphangion in the in vivo mesenteric lymphangion. In the control group (○, open circles), there were no significant time-dependent changes in the frequency, and end-diastolic and end-systolic diameters of the lymphangion. In contrast, the physiological saline solution containing 30 μM L-NAME perfused on the in vivo lymphangion rapidly increased the frequency of active pump and simultaneously decreased significantly the end-systolic and end-diastolic diameters of the lymphangion (●, solid circles). Simultaneous perfusion of 1 mM

Effects of 30 μM L-NAME (●, n = 4) on the pumping frequency
These findings suggest that endogenous NO has physiologically modulated the lymphatic pump activity in rat mesenteric lymphangion in vivo. Thus, lymph flow-mediated shear stress stimulation accelerates to release endogenous NO from the endothelial cells of the lymph vessels in vivo and then regulates the spontaneous contraction-mediated active lymph transport in vivo.
Lymph Flow Accelerates ecNOS mRNA and the Protein Expression in LEC
It is known that exercise training enhances endothelium-dependent vasodilation 42 and the expression of ecNOS mRNA in dogs. 43 Thus, shear stress stimulation increases ecNOS mRNA expression in aortic and arterial endothelial cells.44,45 In addition, Yamamoto et al. 46 demonstrated that shear stress stimulation released ATP in human pulmonary arterial endothelial cells through the activation of cell surface ATP synthase.
However, no study except for one 47 demonstrated that lymph flow accelerated the ecNOS mRNA and the protein in the LEC. In addition, ATP released from the LEC by the stimulation of lymph flow played key roles of the lymph flow-mediated ecNOS mRNA and the protein expression in the lymphatic cells. Figure 5 shows representative recordings of lymph flow-mediated shear stress stimulation-dependent ecNOS mRNA and the protein expression in cultured human LEC. Figure 6A shows representative photomicrographs of ecNOS in the LEC produced by 2-hour shear stress stimulation at three different strengths.
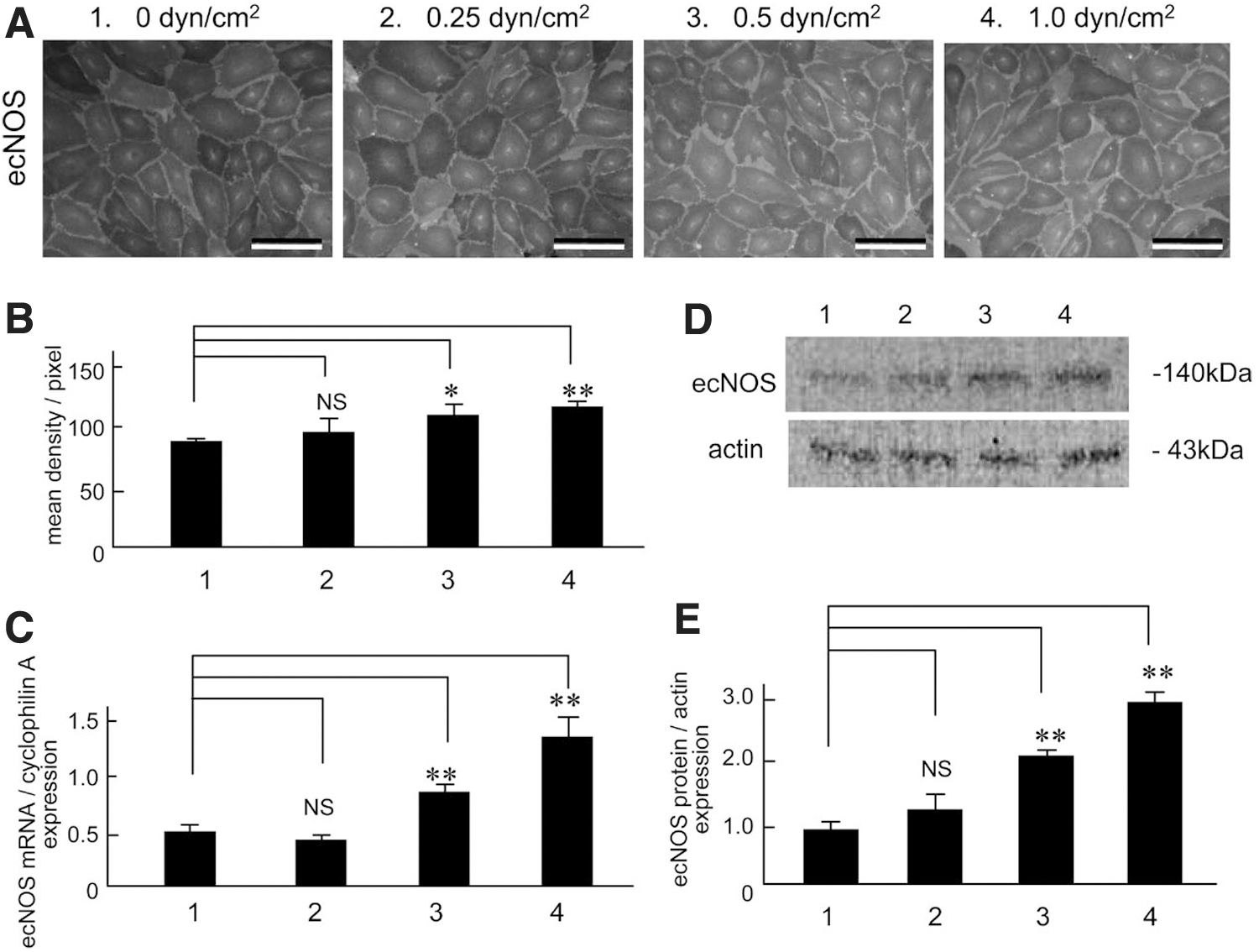
Effects of shear stress strength on ecNOS mRNA
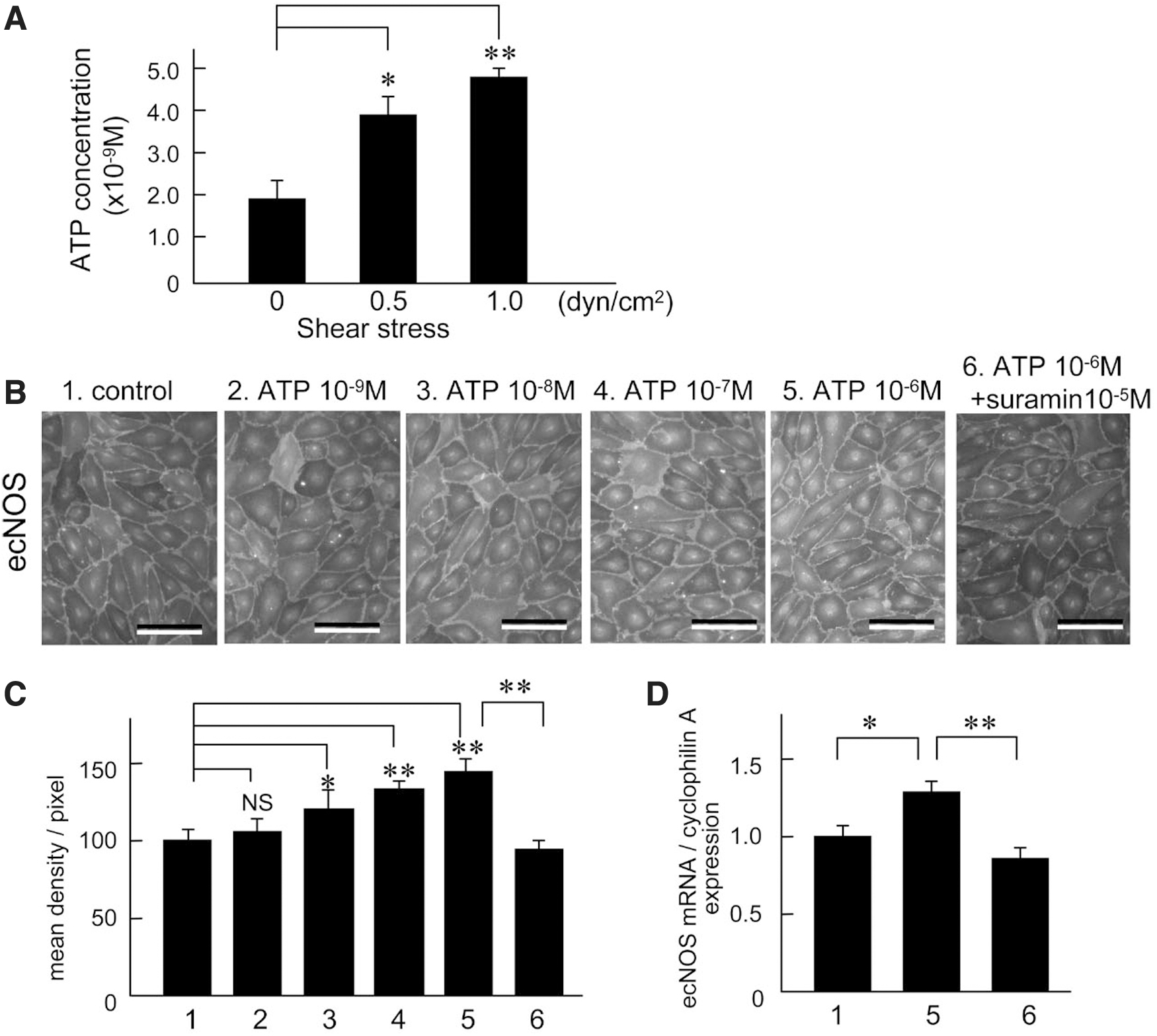
The intensity of shear stress was calculated using the formula, 6 μQ/a2b, where μ is the viscosity of the perfusate, Q is the flow volume, and a and b are the cross-sectional dimensions of the flow path in the parallel plate-type experimental apparatus. The ecNOS expression measured with density measurements was increased dependent with the shear stress strength (Fig. 5B, n = 5). Figure 5C is the summarized data for the effects of shear stress strength on ecNOS mRNA expression in human LEC, as evaluated by reverse transcription-polymerase chain reaction (n = 4). Figure 5D shows representative tracings of the three levels (1, 2, 3, and 4 are 0, 0.25, 0.5 and 1.0 dyn/cm2, respectively) of shear stress-induced ecNOS protein expression, as evaluated by Western blot analysis (n = 5). The ecNOS protein expression increased dependent with shear stress strength (Fig. 5E).
The same strength of shear stress stimulation produced a significant release ATP from the LEC through the activation of cell surface F1/Fo ATP synthase (Fig. 6A). 47 Exogenous ATP also produced a significant increase of ecNOS immunocytochemical (Fig. 6B, C) and ecNOS mRNA expression (Fig. 6D). Suramin, a purinergic P2X/2Y receptor antagonist, caused a significant reduction of the shear stress stimulation-mediated increases in ecNOS immunohistochemical and ecNOS mRNA expression.
These findings suggest that shear stress stimulation produces a significant release of ATP from LEC, which activates the purinergic P2X/2Y receptor, thereby facilitating ecNOS mRNA and the protein expression. The conclusion is shown in the graphical abstract (Fig. 7). Thus, it is clarified that lymph flow releases NO in cooperation with accelerating ecNOS mRNA and the protein expression in the LEC at physiological condition. In the other words, the lymph flow-mediated shear stress stimulation on the LEC regulates the NO production and release with negative feedback system.
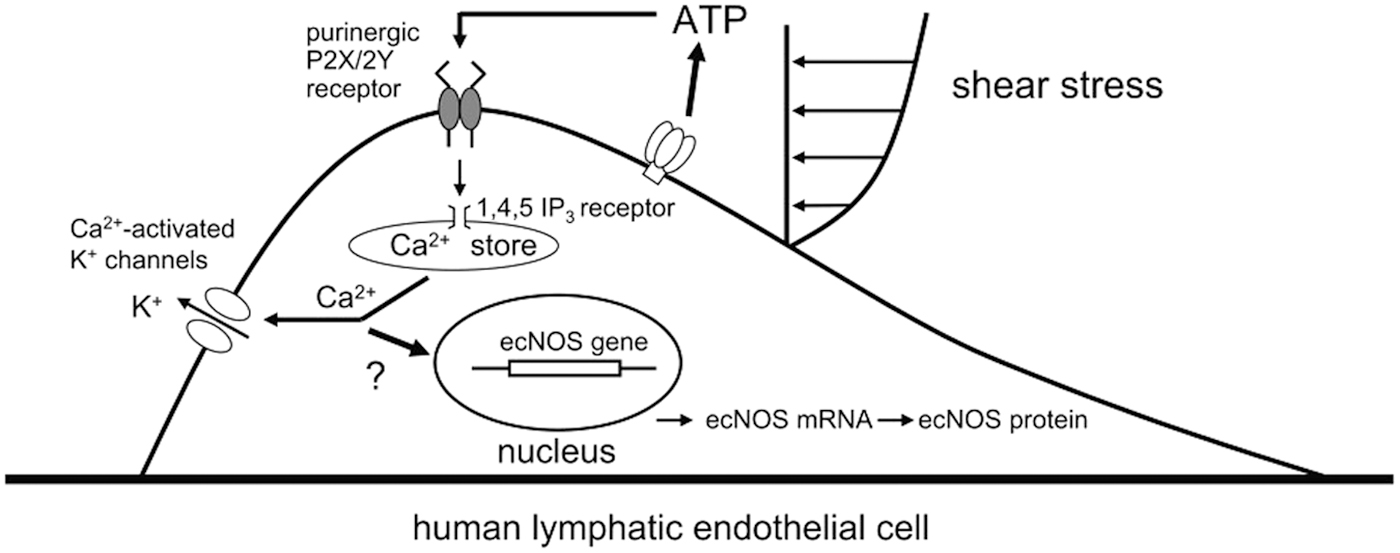
Schematic diagram outlining the physiological roles of shear stress and ATP in the regulation of ecNOS mRNA and the protein expression in LEC. 1,4,5 IP3, inositol 1,4,5-trihosphate. With permission from The American Journal of Physiology, 2010. 47
Collaboration with Aminergic Nerve Activity and Lymph Flow-Mediated NO Release in the Control of Lymph Production and Transport
Once one lymphangion is distended by inflowed lymph volume, it contracts through the activation of stretch-induced contraction of lymphatic smooth muscles. According to the contraction of the lymphangion, lymph fluid in the lymphangion is pushed forward to the next one. Thus, each lymphangion operates as a separate individual pump. The lymphatic pump activity is defined as active propulsion of lymph produced by rhythmical spontaneous contractions of lymphatic smooth muscles in each lymphangion. 6
It is well known that sympathetic aminergic nerve fibers innervate in bovine and human mesenteric lymph vessels.6,48–50 Especially, high density of aminergic sympathetic nerve fibers is innervated into the walls of bovine mesenteric lymph vessels. 48 The norepinephrine released from the nerve fibers regulates the frequency of heart-like spontaneous contractions. Thus, the released norepinephrine decreased the frequency when the pacemaker site of spontaneous contraction locates around valvular region. In contrast, the norepinephrine increased the frequency when it distributes at intervalvular region. 10 In addition, in physiological condition, the activation of aminergic nerve fibers induced the movement of pacemaker site of spontaneous contraction from the valvular region at downstream to intervalvular one through the activation of β-adrenergic receptors with noradrenaline on the smooth muscles in the pacemaker site (Fig. 8).
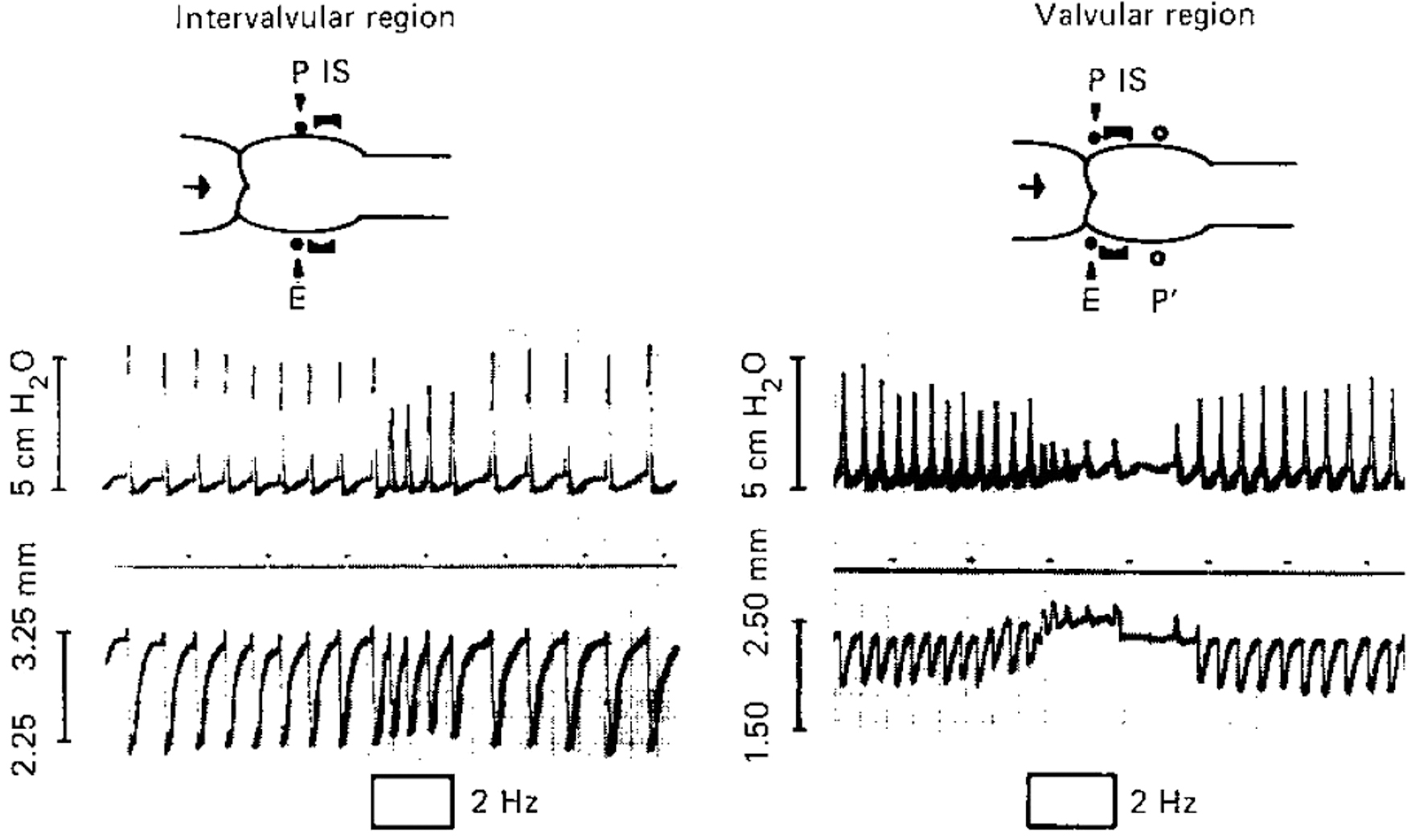
Representative two kinds of responses of isolated bovine mesenteric lymphangion with active pump activity to electrical stimulation (2 Hz, 0.5 ms, 50 V; selective parameter for the stimulation of aminergic nerve fibers in the wall). P shows the pacemaker site of active pump activity; P′ is a new pacemaker site after the electrical stimulation (intervalvular region). IS and E demonstrate the recording position of the outer diameter of lymph vessel with a diameter gauge and the position of electrical stimulation, respectively. The pacemaker site with the valvular and intervalvular regions shows peristaltic and pendular-type movement of active pump activity on the lymphangion, respectively. With permission from Ohhashi 10 with Karger. 10
The movement of pacemaker from the valvular to intervalvular region produced the change in the contraction style, from the peristaltic movement to pendular-type movement combined with several lymphangion. On the result, the pendular-type movement may contribute to absorb strongly the fluid and protein through lymphatic capillaries from the interstitial space, resulting in increase of lymph production and lymph transport. 10 The large amount of lymph transport may produce higher lymph flow in the upstream lymphangion, which accelerates large amount of NO release by the shear stress stimulation, resulting keeping much volume of lymph in the upstream lymphangion. When the pacemaker site returns at the valvular region with decreasing aminergic nerve activity, the stored lymph volume will delivery efficiently into the upstream lymphangion. Thus, the collaboration of aminergic nerve activity and lymph flow-mediated NO release may be useful for large amount of lymph transport.
Conclusion
In the lymphatic system, lymph flow plays an important role in regulation of lymph transport through the release of endogenous NO from the endothelial cells and activation of ecNOS mRNA and the protein expression in physiological condition. Thus, in the mesenteric lymph vessels, the lymph flow-mediated NO release inhibits the pacemaker activity of active lymph pump, resulting in increasing lymph volume of one lymphangion. In addition, the activation of aminergic nerve fibers in the mesenteric lymph vessels moves the pacemaker site from the valvular to intervalvular region, resulting in change in the heart-like spontaneous contraction from peristaltic to pendular-type movement.
The change in the spontaneous contraction may accelerate lymph production. The increasing volume in the lymphangion produced with NO release or increase in aminergic nerve activity stretches the smooth muscles in the wall, thereby producing stretch-mediated larger contraction of the smooth muscle cells, which contribute to be more efficient lymph transport. In physiological condition, the collaboration of aminergic nerve activity and lymph flow-mediated NO release may play key roles of lymph transport.
Footnotes
Ethics
Animal experiments in the present study were performed with the approval of Shinshu University Animal Care and Use Committee.
Consent for Publication
All authors approved the final version of the article and accepted for the publication of this article.
Data Availability Statement
All relevant data are available from the corresponding author on request.
Acknowledgments
We thank Ms. Yumiko Yokoyama and Ms. Maki Kaidoh in Shinshu University School of Medicine for their help to conduct the experiments.
Authors' Contributions
T.O. and Y.K. designed the experiments, analyzed the data, and wrote the article. Y.K., M.H., T.-W.A., and D.M. performed the experiments, analyzed the data, and revised the article.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
The Department of Innovation of Medical and Health Sciences Research at Shinshu University School of Medicine has been established and supported financially by the donation (2014-present) of BOURBON, Co., Ltd., Kashiwazaki, Niigata, Japan, and Aizawa Hospital, Matsumoto, Nagano, Japan. The authors declare that this study received funding from BOURBON Co. Ltd. The funder was not involved in the study design, collection, analysis, interpretation of data, the writing of this article, or the decision to submit it for publication.