
Abstract
A new single-chain variable fragment (scFv) antibody library was generated and human serum albumin (HSA)-specific clones were characterized to investigate the usefulness of porcine antibodies. Phage libraries were developed from pigs immunized with the model antigen HSA. The library size was 1.5×107 for kappa (VL) and 1.4×107 for lambda fragments. Eight HSA-specific clones from the kappa library and one clone from the lambda library were isolated using affinity selection. The binding specificity of these clones was confirmed using a phage enzyme-linked immunosorbent assay (ELISA). The scFvs were expressed in Escherichia coli and purified from the periplasm fraction for further investigation. Based on the results of ELISA and Western blot analysis, four scFv clones with high activity and high yield were selected and purified. The purified scFvs from four of the nine clones exhibited an approximate K D of 10−8 M. This is the first report describing isolation of HSA-specific porcine scFv antibodies from an antibody phage library and characterization of their binding properties.
Introduction
A
Phage display technology, first reported by Smith,(2) is a powerful tool for generating the high-affinity binders and has been widely used to isolate MAb fragments, such as single-chain variable fragments (scFv) and antigen-binding fragments, from various animals, including camels,(3) rabbits,(4) and chickens.(5) Using this method, antibodies are expressed as fusion proteins with phage coat proteins and displayed on the surface of the phage. Once the antibody phage library is constructed from various genetic sources, it is possible to obtain specific antibody phages harboring the genes of interest through biopanning.
Pigs (Sus scrofa) have long been bred for meat, but recently their value as experimental animals for medical research has been reviewed.(6,7) The use of pig tissue in xenotransplantation has also been examined.(8–10) There are a few reports regarding the production of pig antibodies. Almagro and colleagues conducted comparative analyses of the VH gene in horses (Equus caballus), sheep (Ovis aries), cattle (Bos taurus), and pigs. They found that pig antibody genes are the most similar to human VH3.(11) Expression analysis of porcine antibodies has indicated that only a limited number of genes are expressed,(12) including five of the VH3 family members, which cover 80% of expressed antibodies, a single VL kappa family member (IGKV2), and two VL lambda family members (IGLV3 and IGLV8).(13) Pig antibodies are produced from a small number of genes compared with those in humans and mice, with the immunological diversity of porcine antibodies attained by hypermutations.(14)
Porcine scFv antibodies binding to a specific antigen using phage display have not been investigated in detail. In this study, we cloned several porcine scFv antibodies that specifically bound to human serum albumin (HSA) using phage display and analyzed the properties of the obtained scFvs.
Materials and Methods
Immunization of pigs
A pig was immunized with 12 mg of HSA (Sigma-Aldrich, Tokyo, Japan) and Freund's complete adjuvant (Difco, Franklin Lakes, NJ) via intramuscular injection. Two weeks post-immunization, a booster containing 12 mg of HSA and incomplete Freund's adjuvant (Difco) was administered. After confirming sufficient antibody titer using an enzyme-linked immunosorbent assay (ELISA), a blood sample (200 mL) was obtained and 6.3×108 peripheral blood mononuclear cells (PBMCs) containing B cells were collected by Ficoll-1077 density gradient separation (Wako Chemicals, Osaka, Japan).
Phage library construction
Total RNA was extracted from the collected PBMCs using RNAiso plus (Takara Bio, Shiga, Japan). First strand cDNA was synthesized with Prime Script II (Takara Bio) using oligo dT primers, according to the manufacturer's protocols. The VH-specific primers (VH-B and VH-F) for PCR amplification were designed to cover a large portion of the porcine VH genes based on sequence information from the IMGT database (International ImMunoGeneTics information system;
Primer pairs were used for the first- (
The heavy and light chain fragments were amplified using KOD-plus neo DNA polymerase (Toyobo, Osaka, Japan) using PCR primers (0.3 μM, Table 1). The PCR was performed in a volume of 50 μL and involved an initial denaturation step at 94°C for 2 min, then 25 cycles of 98°C for 10 s, 55°C for 30 s, and 68°C for 15 s. A second PCR was used to add restriction sites to the amplicons from the initial PCR. The primers we used were specific (Table 1B), and thermal cycling conditions were the same as those used in the first round of PCR. A third round of PCR amplification was used to connect the DNA fragments corresponding to the heavy and light chains with GS linker DNA (Table 1C). The PCR was conducted using Gene Taq DNA polymerase (Nippon Gene Co., Tokyo, Japan) in the presence of 4.5 nM VH and VL fragments and linker DNA with 0.45 μM VH-B-Sfi and VLκ-F-Not or VLλ-F-Not primers (Table 1B). The PCR thermal cycling profile involved an initial denaturation step at 94°C for 2 min, then 20 cycles of 94°C for 30 s, 60°C for 30 s, 72°C for 30 s, and a final extension step at 72°C for 5 min. The scFv gene amplicons generated by the third round PCR were separated by agarose gel electrophoresis and purified with a QIAquick Gel Extraction Kit (Qiagen, Hilden, Germany). Subsequently, scFv gene amplicons were digested with SfiI and NotI (New England Biolabs, Beverly, MA) restriction enzymes, then ligated into pKSTV-01 (Fig. 1). The vector containing the scFv gene fragment was purified using phenol/chloroform, precipitated with ethanol, and dissolved in distilled water. The vectors were transformed into Escherichia coli TG-1 electrocompetent cells (Lucigen, Madison, WI). Generated library sizes were evaluated by counting the colonies formed from an aliquot of transformed cells; they were 1.5×107 for VL kappa and 1.4×107 for lambda.
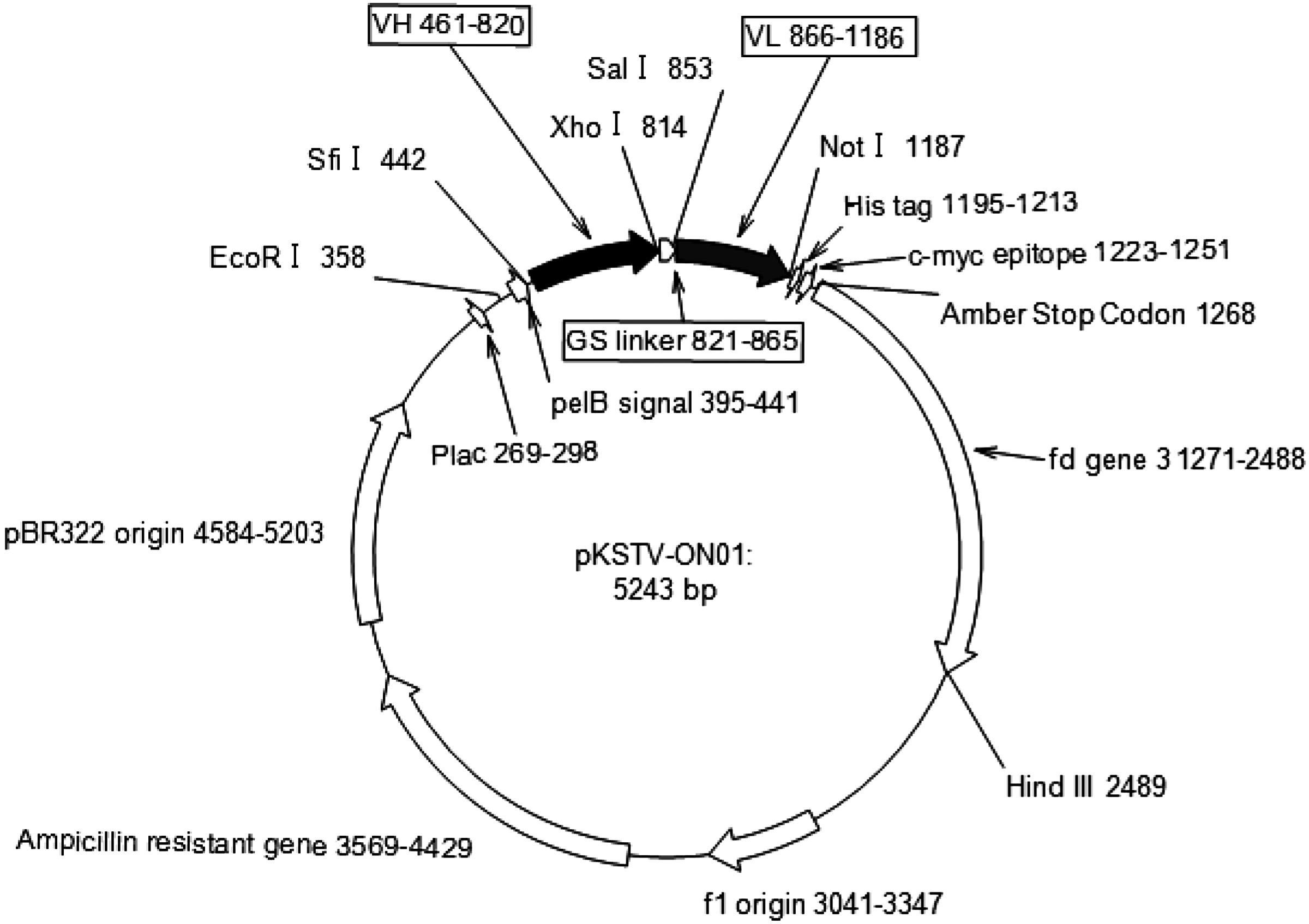
Genetic map of pKSTV-01 phagemid vector. This vector was constructed based on pTV118N from our laboratory. VH and VL depicted in the box indicate typical human VH and VL genes in the vector.
A scFv phage display library was prepared as previously reported.(15) TG-1 cells transformed with phagemid vector were grown to log phase in 2TYAG medium (2TY medium [1.6% tryptone, 1% yeast extract, 86 mM NaCl] containing 2% glucose and 100 μg/mL ampicillin), and then infected with M13K07 helper phage (Invitrogen, Carlsbad, CA) for 30 min at 37°C without shaking. Following centrifugation (1000 g, 10 min), the pellet was suspended in 2TYAK (2TY medium containing 100 μg/mL ampicillin and 50 μg/mL kanamycin) and incubated overnight at 30°C with shaking. The culture was centrifuged (6000 g, 15 min), and the obtained supernatant was mixed with a one-fifth volume of 2.5 M NaCl and 20% (w/v) polyethylene glycol 6000 (PEG 6000). Following incubation for 6 h at 4°C, the precipitated phages were collected by centrifugation, dissolved in 1 mL of phosphate-buffered saline (PBS), and used for biopanning.
Selection of specific phages that bound to HSA
Wells of an ELISA plate (Maxisorp, Nalge Nunc, New York, NY) were coated with 100 μL of HSA (20 μg/mL) dissolved in 0.1 M NaHCO3 (pH 8.3) at 4°C overnight. Wells were washed with PBS, blocked with 0.5% gelatin in PBS at room temperature for 2 h, and then washed three times with PBS containing 0.1% Tween-20 (PBST). Phage solution (100 μL), containing 0.5×1011 colony forming units (cfu) of phages suspended in PBS supplemented with 0.5% gelatin, was added to wells and incubated at room temperature for 1 h with shaking. Wells were washed 10 times with PBST, and bound phages were eluted in 300 μL of 0.1 M glycine-HCl buffer (pH 2.2) for 10 min. The eluate was neutralized with 26 μL of 1 M Tris-HCl buffer (pH 9.1) to a final pH of approximately 7.0. Phages were then infected into fresh E. coli TG-1 cells and used for a second round of panning. Wells were coated with HSA (10 μg/mL) and the phage solution in 1% skim milk. The E. coli TG-1 cells were infected with eluted phages, spread on 2TY agar plates (2TY medium containing 1.5% agar, 2% glucose, and 100 μg/mL ampicillin) and cultured at 30°C overnight. Colonies were picked, transferred to 1 mL of 2TYAG medium, and incubated at 30°C overnight. The overnight culture was diluted 100-fold with 2TYAG and cultivated to the log phase of growth. M13K07 helper phages were added to cultures at a multiplicity of infection of 10, and the cultures were incubated at 37°C for 30 min without shaking. After centrifugation (1000 g, 10 min), the pellets were collected, resuspended in 2TYAK, and incubated at 30°C overnight. After further centrifugation (13,000 g, 30 min), supernatants were collected and assayed immediately or after precipitation with PEG 6000.
ELISA
The wells of an ELISA plate were coated with 50 μL of HSA (1 μg/mL) dissolved in 0.1 M NaHCO3 (pH 8.3), and incubated at 4°C overnight. Wells were washed with PBS and then blocked with 0.5% gelatin in PBS (PBSG) at room temperature for 2 h. After washing three times with PBST, 50 μL of phage solution in PBSG were added to the wells. Following a 1 h incubation with shaking, wells were washed five times with PBST. A horseradish peroxidase (HRP)-conjugated anti-M13 antibody (50 μL; GE Healthcare, Waukesha, WI) was diluted 1:5000 in PBS containing 1% gelatin, added to wells, and incubated for 1 h with shaking. Wells were washed five times with PBST, and 50 μL of 3,3′,5,5′-tetramethylbenzidine solution (One-step Ultra, Thermo Fisher Scientific, Rockford, IL) were added. After 5 min, the reaction was terminated by adding H2SO4. The absorbance at 450 nm was measured using a microplate reader (Tecan Group, Männedorf, Switzerland).
Bacterial expression of scFv
Cultures of E. coli HB2151 infected with individual phage clones were incubated on 2TYAG agar plates at 30°C overnight. Colonies were transferred into 2TYAG liquid medium and cultured at 30°C overnight. The culture was diluted 1:100 in 2TYAG and grown until the optical density was 0.5 at 600 nm. After centrifugation, the collected pellets were suspended in 2TY medium containing 100 μg/mL ampicillin and 1 mM isopropyl-β-D-thiogalactopyranoside and incubated at 30°C for 6 h. After removing the supernatant by centrifugation, the pellets were suspended in cold (4°C) buffer (10 mM Tris-HCl, 10 mM EDTA, 20% (w/v) sucrose [pH 8.0]) and incubated on ice for 10 min. After centrifugation (20,000 g, 15 min), the collected pellet was suspended in cold water (4°C), kept on ice for 10 min, and centrifuged again (20,000 g, 15 min), and the supernatant containing the periplasm fraction was collected. Pellets were resuspended in PBS and sonicated for 5 min using a Sonifier 450 (Branson, CT). After centrifuging (20,000 g, 15 min), the supernatant containing the cytoplasmic fraction was collected.
Western blot analysis
The cytoplasmic and periplasmic fractions were subjected to sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE; 12% TGX gel; Bio-Rad Laboratories, Hercules, CA) and transferred to polyvinylidene difluoride (PVDF) membranes (Hybond-P, GE Healthcare) using a semi-dry membrane transfer system (GE Healthcare). After blocking with 5% skim milk in PBS, scFv was detected using an HRP-conjugated anti-Myc-Tag MAb (HRP-Direct, Medical and Biological Laboratories Co., Nagoya, Japan). An ECL Plus Western Blotting Detection System and Image Quant 350 (both from GE Healthcare) were used to detect positive signals following a 5-min exposure time.
Binding affinity analysis
Periplasmic fractions of E. coli HB2151 were subjected to affinity chromatography on HisTalon Superflow Cartridges (Takara Bio), according to protocols provided by the manufacturer. Purified scFv was dialyzed against PBS and used for binding affinity analysis on a Biacore T100 system (GE Healthcare). We used HBS-EP buffer (0.01 M HEPES [pH 7.4], 0.15 M NaCl, 3 mM EDTA, 0.005% Surfactant P20) at a flow rate of 50 μL/min and 25°C. HSA was immobilized to a CM5 sensor chip up to 251 response units (RU) using an amine coupling protocol. A scFv was diluted in HBS-EP buffer at different concentrations and injected to monitor association and dissociation reactions for 2 min per reaction. A nscFv-immobilized chip was regenerated with 0.1 M glycine buffer (pH 2.0) for 15 s. Affinity kinetic parameters were calculated using evaluation software (GE Healthcare) assuming a 1:1 binding model.
Results
Porcine scFv phage display library
We constructed a scFv phage display library using B cells from pigs immunized with HSA. The genes encoding the variable region of the heavy chain (VH) and the variable region of light chain (VL) were amplified using specific primer sets (Table 1). The library sizes were 1.5×107 for VL kappa and 1.4×107 for VL lambda.
Isolation and analysis of HSA-specific scFv phage clones
Phages (n=96) from each library were cloned and screened for binding capacity using ELISAs. Clones from the VL kappa (n=10) and VL lambda (n=3) libraries showed specific binding to HSA. DNA sequence analysis of scFv genes in these clones indicated nine variations in the scFv sequence, designated B7, C10, D4, D8, E1, E4, E11, G3, and H6. The binding specificity of these scFv phages is shown in Figure 2. Eight of the nine clones specifically bound to HSA, while one clone (H6) bound to both porcine serum albumin and HSA.
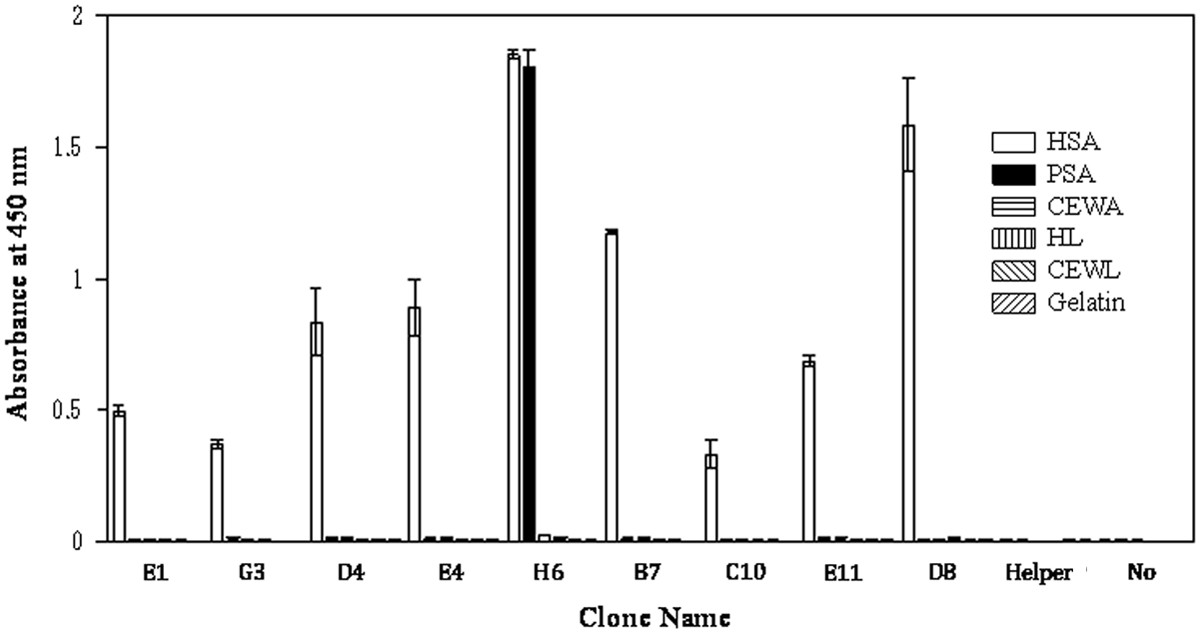
Binding specificities of isolated phage clones. Isolated phage clones were screened by ELISA. HSA, human serum albumin; PSA, porcine serum albumin; CEWA, chicken egg white albumin; HL, human lysozyme; and CEWL, chicken egg white lysozyme. “Helper,” helper phages added to wells. “No,” no phages were added to wells.
Amino acid sequences of isolated scFvs
The amino acid sequences of scFv clones are summarized in Figure 3. The sequence of the GS linker was GGGGSGGGGSGSTGS, which differs from the normal GS linker sequence (GGGGSGGGGSGGGGS) because of the restriction enzyme sites at the end of the GS linker. Seven of the clones, with the exception of H6 and D8, shared almost identical VH sequences. The three complementarity determining regions of VH were identical. Analysis of VH DNA sequences showed that they were most similar to IGHV1S1 (IMGT accession no. AF064686); the VH gene for D8 was most similar to IGHV1S2 (AF064688). The JH DNA sequence in our clones was most similar to that for IGHJ1 (AJ554305); we observed a single amino acid substitution (Pro to Ala in VH-FR4) in the JH segment of D8.

Amino acid sequences of anti-HSA scFvs. Amino acid sequences of scFvs isolated from the library were aligned with 4D5 scFv sequence. Dots indicate unchanged amino acids. Bars indicate deletions.
The D8 clone contained a VL lambda chain, while the other eight clones had a VL kappa chain. Among the VL sequences, with the exception of D8 and H6, nucleotide identities were high (>85%), with the IGKV1 gene being common among these clones. The VL sequences of D8 and H6 were very different from the other clones with low sequence identities (60 and 40%, respectively), and were also very different from each other (38% identity).
Expression of scFv clones in E. coli
For the E1, G3, D4, E4, H6, and E11 clones, scFvs were expressed in the periplasmic and cytosolic fractions (Fig. 4). For the B7 and D8 clones, scFvs were only expressed in the periplasmic fraction at low levels. The purity of purified scFvs was verified by SDS-PAGE (data not shown). Prominent bands were observed at approximately 27 kDa, corresponding to the molecular weight calculated from amino acid sequences. The yield of scFv was around 1000 μg/L from clones E4 and D4, 220 μg/L from E11, and 110 μg/L from H6.

Detection of scFv expressed in bacteria. E1, G3, D4, E4, H6, B7, C10, E11, and D8 porcine scFv clones were expressed in E. coli HB2151. Supernatant (S), periplasm (P), and cytoplasm (C) fractions were collected and subjected to SDS-PAGE. Proteins were transferred to PVDF membrane and detected with an anti-myc-tag antibody.
scFv affinity analysis
The binding affinities of scFvs were measured using a CM5 sensor chip immobilized with HSA in a Biacore T100 system (Fig. 5). The association (ka) and dissociation (kd) rate constants were estimated, and a dissociation equilibrium constant (K D ) was calculated. The K D values of the scFvs were between 9–21 nM (Table 2).

Affinity analysis of scFv. The binding affinity of four porcine scFvs were analyzed with Biacore T100. The SPR response curves are indicated for D4 (
Association and dissociation rate constants were measured with a Biacore T-100 using a CM5 sensor chip immobilized with HSA at 25°C.
Discussion
In this study we generated HSA-specific porcine antibodies using scFv phage display libraries. We found that nine porcine antibodies specifically bound to HSA; seven of the clones (D4, E4, E11, E1, G3, B7, C10) contained the same VH sequences but different VL sequences. This suggests that the VH domain plays a crucial role in antigen binding for these scFvs.
Although it seems that the VL domain did not significantly contribute to antigen specificity (Fig. 2) or affinity (Table 2) among scFvs harboring the same VH (D4, E4, E11, E1, G3, B7, C10), it did affect the production of scFvs in E. coli. Clones E4 and D4 produced the most scFv in the periplasmic fraction, yielding 1000 μg of protein/L of bacterial culture. The amount of scFv produced by E11 was about 20% of the level produced by either E4 or D4. This would suggest that VL affects the stability and/or folding of scFvs, thereby altering their yield during bacterial expression.
We compared sequences of porcine VH with those in human antibodies. The VH segment of porcine antibodies generated in this study was homologous to human IGHV3-23. This was consistent with a previous report demonstrating high homology between human VH3 antibody and a pig antibody.(12) The repertoire of VH genes present in pigs is small, consisting of only one subgroup (IGHV1), compared with the seven subgroups (IGHV1–7) in humans. The low number of porcine antibody sequences makes it easier to construct an antibody library and to predict mutations that are responsible for conferring antigen specificity and affinity.
In conclusion, we have described the properties of HSA-specific porcine scFv antibodies selected with a phage display library. The obtained clones demonstrated high affinity and low cross-reactivity. This study will provide a basis for further porcine antibody research.
Footnotes
Author Disclosure Statement
The authors have no financial interests to disclose.