
Abstract
The αβ T cell antigen receptor (TCR) endows T lymphocytes with immune specificity and controls their effector functions. Each person possesses a vast repertoire of TCRs that is generated by the well-studied processes of somatic recombination and thymic selection. While many antibodies specific for TCRβ variable domains are available, antibodies specific for human TCRα are rare. We now report a novel monoclonal antibody, 7F18, which binds to human TCRα constant region, with specificity for a denatured epitope that can be visualized by SDS-PAGE followed by Western blot. Both immature and mature TCR α-chain products can be visualized, making 7F18 potentially applicable to various biochemical assays of multiprotein complex assembly and maturation. This new monoclonal antibody provides a tool that can potentially facilitate the biochemical analysis of comprehensive populations of human αβ TCR complexes that need not be limited to small subsets of the repertoire.
Introduction
A
To better understand TCR function, extensive efforts have focused on the genetic, structural, biophysical, and biochemical characterization of the receptor complex. Assembly of the multiprotein complex begins with the generation of nascent TCR/CD3 polypeptides in the rough endoplasmic reticulum (ER). The heterodimers TCR αβ, CD3 ɛγ, and CD3 ɛδ are folded separately and then assembled together, with final addition of ζζ homodimer to form αβ/ɛγ/ɛγ/ζζ.(11–13) Upon completion of the assembly process, the TCR/CD3 complex undergoes conformational maturation and glycan modification as it progresses through the Golgi apparatus along the secretory pathway.(14) A marker for progression of complex maturation is provided by TCRα molecular weight, which before multi-subunit assembly is lower and displays incomplete glycosylation, and upon assembly and complex maturation appears relatively higher as glycan processing is completed.(15) Recent data suggest that final, surface-expressed TCR/CD3 complexes might form monovalent, bivalent, and higher-order complexes,(16–18) but complete biochemical analysis of such complexes has not been performed, and their relative representation among human TCRs has not been determined.
Much of this information was acquired using antibody-dependent assays. One monoclonal antibody (MAb) clone has been of outstanding utility in this regard: H28-710, specific for the constant domain of mouse TCR alpha-chain.(15,19–22) Prior to the current work, characterization of a MAb of equivalent function with specificity for human TCR alpha-chain had not been published. Most MAbs that bind human TCRs are specific for one of many different Vβ gene products, while MAbs that are specific for human Vα-chains have been reported less frequently. We report the generation of a MAb (7F18) whose specificity for Cα is designed to detect all human TCR alpha-chains by SDS-PAGE followed by Western blot (WB). This MAb recognizes an epitope present within the constant region of human alpha-chains, an epitope that is available in both immature/unassembled and mature/assembled protein products. We propose that by facilitating the detection of TCR alpha-chains by WB, 7F18 MAb represents a useful tool that is likely to contribute to resolving outstanding questions regarding the biochemistry and multiprotein assembly of human αβ TCRs.
Materials and Methods
Antigen design and MAb generation
We submitted the peptide sequence QLRDSKSSDK to Abmart, Inc. (Shanghai, China), which passed their in-house, bioinformatic predictive algorithm criteria as possessing adequate antigenicity for MAb development. By contractual agreement, Abmart synthesized the peptide and used it to immunize mice and generate a panel of MAb-secreting candidate hybridomas that our laboratory screened for the desired assay functionality and specificity. Abmart's role was limited to MAb generation, and the company retained no intellectual property rights that could limit or otherwise control dissemination of the MAb to the scientific community.
Cells
Jurkat, JRT3, JRT3.huCD8ab.LC13, and COS1 cell lines were maintained by weekly passage in either RPMI-1640 (Jurkat, JRT3, JRT3.huCD8ab.LC13) or DMEM (COS1) containing 10% HyClone fetal bovine serum (ThermoScientific, Waltham, MA). The JRT3.huCD8ab.LC13 cell line was kindly provided by Zhenjun Chen and James McCluskey (University of Melbourne, Australia).(23) RPMI-cultured cells were incubated in 5% CO2 atmosphere, while DMEM-cultured cells were incubated in 10% CO2 at 37°C. Human peripheral blood mononuclear cells (PBMC) were Ficoll-purified from leukoreduction system chambers as described,(24) in accordance with Mayo Clinic's Institutional Review Board regulations.
Lysates
Cells were lysed in one of two buffers as indicated: (1) digitonin lysis buffer: 0.05 M Tris, 0.15 M NaCl, with freshly added protease inhibitors (P2714 and A8456l, Sigma-Aldrich, St. Louis, MO) and 1% digitonin (ultrapure grade, Calbiochem, San Diego, CA); or (2) mild lysis buffer (MLB): 0.15 M NaCl, 0.25 M Hepes, 0.005 M EDTA, 0.0005 M CaCl2, 10% glycerol (G5516-1L, Sigma-Aldrich), 0.2% Tween-20 (170-6606-MSDS, Bio-Rad, Hercules, CA), 0.2% NP-40 (56741, Fluka, Seelze, Germany).(25) Lysis was performed at a concentration up to 50×106 cells/mL of lysis buffer. Nuclei and cellular debris were removed through centrifugation such that postnuclear lysates were used in all experiments.
Size exclusion chromatography
The Superose 6 column (GE Healthcare, Little Chalfont, United Kingdom) was equilibrated in running buffer (0.1% Brij-58 (Sigma-Aldrich), 50 mM Tris (pH 7.4), 150 mM NaCl) as part of the Akta FPLC system (GE Healthcare) operated in a chromatography refrigerator at 10–16°C. Samples were injected over the Superose 6 column in a sample volume of 2 mL and run at a flow rate of 0.75 mL/min. Elution profiles for the samples were monitored by UV absorbance at 280 nm. Fourteen 1 mL fractions were collected beginning at the fraction containing the void volume (5.75–6.75 mL=fraction 1) and spanning all fractions that could contain cellular protein. Following fractionation, 1 mL of acetone was added to each 1 mL fraction, and acetone-precipitation of protein occurred overnight at −20C.
Immunoprecipitation and pull-down assays
TCR complexes from digitonin-buffer lysates of Jurkat or human PBMC were used. For immunoprecipitation, SEC-fractionation of digitonin cell lysate was used to enrich for fully assembled, mature TCR/CD3 complexes (fractions 6–8, Superose 6 column). Anti-human CD3-epsilon (UCHT1) was used to immunoprecipitate by standard methods, and captured complexes were subjected to reducing SDS-PAGE and Western blotting, as indicated. Anti-CD3-zeta serum 448 was kindly provided by Balbino Alarcón (CBMSO, Universidad Autónoma de Madrid, Spain).(26)
For GST-Nck-SH3.1 pull-down assays, published methods were used.(24–27) Briefly, lysate from JRT3.huCD8ab.LC13 cells was incubated on ice for 30 min in the presence of 10 μg/mL of either APA1/1 (anti-CD3-epsilon, intracellular tail) or OKT3 (anti-human CD3-epsilon extracellular domain), followed by incubation with GST-Nck-SH3.1 beads overnight at 4°C. Captured TCR/CD3 complexes were subjected to reducing SDS-PAGE and subsequent Western blot analysis with 7F18.
Western blot analysis
Following acetone precipitation, samples were subjected to either reducing or non-reducing SDS-PAGE (10% acrylamide/bisacrylamide, Bio-Rad) and transferred to nitrocellulose membranes (Bio-Rad) by standard methods. Membranes were blotted with a primary MAb (7F18 or others, as indicated) and secondary antibody (Anti-Mouse IgG-HRP or Anti-Rabbit IgG-HRP, Jackson Immunoresearch, West Grove, PA), followed by substrate addition (Amersham ECL Prime Western Blot Detecting Reagent, GE Healthcare). Bands were either visualized by exposing the blots to photographic film (Amersham Hyperfilm ECL, GE Healthcare), followed by film development and image scanning; or digital image capture utilizing a FluorChem M FM0231 Gel Dock imagine system (ProteinSimple, Santa Clara, CA). Images were analyzed and figures produced using Adobe Photoshop software (San Jose, CA), but no image alteration was imposed using the software. Pixel quantification was performed after importing unmanipulated TIF WB images into ImageJ software (NIH), capturing relevant WB bands within the field of view, and measuring the quantity of pixels detected. A ratio was determined between TCR alpha-chain and CD3 zeta-chain to estimate their relative detection.
Transient transfection
Expression plasmids (pCI-NEO, Promega, Madison, WI) encoding eGFP, TCR-LC13α, or TCR-LC13β were cloned. The human TCR LC13 alpha and beta genes were obtained from retroviral expression constructs kindly provided by Zhenjun Chen and James McCluskey (University of Melbourne, Australia).(23) For transient transfection, COS1 cells were suspended in 250 μL RPMI 1640 containing 5% FBS and 5% FCS (no antibiotics) and mixed with plasmid DNA (30 μg re-suspended in 50 μL RPMI 1640 containing 10 mM HEPES). Electroporation was done in a BTX Electro Square Porator T820 (BTX Corp., San Diego, CA) using standard settings: low voltage, pulse length=10, pulses=1, 4 mm gap cuvettes, voltage=300.
Results and Discussion
By targeting an epitope within Cα, we wished to generate a new MAb that would be specific for all human TCR α-chains. We hoped the MAb would function to bind denatured protein subjected to SDS-PAGE/Western blot (WB), and thus prove as useful for human TCRα analysis as the MAb H28-710 has proven for mouse TCRα analysis.(20–22) An anti-human TCRα MAb of such utility would be a renewable resource, with characterizable properties that would not be expected to change batch-to-batch, as is often seen with polyclonal antisera. We initially analyzed sequence homology between 10 different human TCRα cDNAs, plus human TCRCA consensus sequence, publicly available from PubMed (GenBank accession nos. AF043174, AF043177, AJ004871, AY232282, DQ269212, DQ341450, EU599571, EU862326, EU863816, FJ795363, GQ179994, X02883). All 11 cDNA sequences were in vitro-translated into amino acid sequences using MacVector software, and all sequences were aligned to identify Cα amino acid sequences of 100% agreement between the different sources (data not shown). Sequences passing this criterion were considered highly accurate, and were subsequently assessed for homology to human TCRCB1, TCRCB2, and mouse Tcrca (amino acid sequences extrapolated based on GenBank accession nos. M12887, M12888, and M64239, respectively). Sets of 10 consecutive amino acid sequences that were present in human TCRα but not in the other TCR proteins were identified. Finally, these sequences were subjected to protein BLAST search hosted by NCBI, resulting in one 10 amino acid sequence that was considered to be of high potential for antigen development, QLRDSKSSDK. This sequence, normally expressed in an N-terminal region of Cα (Fig. 1), was used by Abmart, as antigen to generate a panel of hybridomas that we tested for human TCRα specificity.
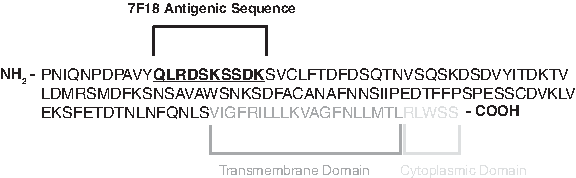
Schematic of human TCRα constant region and pertinent antigenic sequence for 7F18 MAb. 7F18 MAb was generated to detect an epitope within the N-terminal portion of the constant sequence in order to confer TCR clonotype-independent detection of human TCRα. Note that only C-domain sequence is represented, while V-region amino acids (not shown) are contiguous from the N-terminus of the full TCRα protein length.
Each of the candidate MAb clones was first tested by subjecting Jurkat T cell lysates to reducing SDS-PAGE/WB analysis. We observed that several of the MAb clones recognized a protein with apparent molecular weight (MW) matching that expected of human TCRα (∼40–45 kDa, mature form(14)) present within the Jurkat lysates (Fig. 2A, MAbs 2D13 and 7F18; data from similar MAbs not shown). Other MAb clones did not appear to detect TCRα (Fig. 2A, MAb 2M21; data from similar MAbs not shown). Due to strong assay signal, the mouse IgG2b kappa clone 7F18 was selected and further tested for specificity against human TCR.
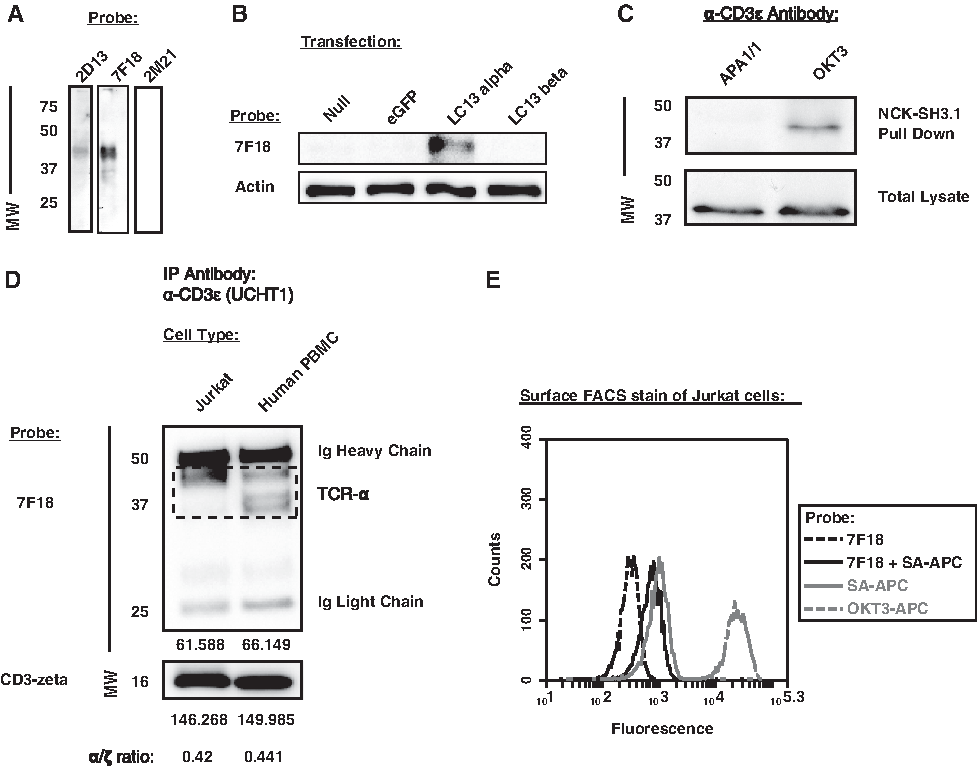
7F18 MAb detects human TCRα by SDS-PAGE followed by Western blot. (
To assess the specificity of 7F18 for human TCRα chain, pCI-NEO-based expression plasmids were cloned (empty vector, eGFP, LC13α, LC13β; LC13 is a well-characterized human anti-viral public TCR(23)) and used for transient transfection of COS1 cells. Upon cell lysis, Western blot showed that MAb 7F18 specifically detected the human LC13 TCR alpha-chain but not LC13β or other negative control transfectants (Fig. 2B).
Next we asked whether 7F18 could detect TCR alpha-chains that were co-associated with CD3 complexes that have adopted a stimulation-induced conformation.(11,27–30) We employed a pull-down assay used to observe a CD3 conformation that can be induced by binding the extracellular domain of CD3-epsilon with specific antibodies. The stimulation-induced conformation is detected when bead-coupled GST-Nck-SH3.1 fusion protein binds to a proline-rich sequence (PRS) in CD3ɛ that is exposed upon MAb binding to the extracellular domain. This specific interaction is blocked by MAb APA1/1, which also binds the PRS of CD3-epsilon.(30) Using JRT3.huCD8ab.LC13 lysate, we found that OKT3 (anti-CD3-epsilon, extracellular domain) induced complexes that could be captured by the GST-Nck-SH3.1 pull-down and probed with 7F18; however, in the presence of APA1/1 MAb, no such complexes were observed by 7F18 Western blot (Fig. 2C). We conclude that MAb 7F18 is able to bind TCR alpha-chains associated with CD3 complexes that have adopted the PRS-exposed conformation.
We wished to estimate the extent to which 7F18 appeared capable of binding to most (or all) human TCR alpha-chains. To do this, we compared detection of human TCRα protein using MAb 7F18 when TCR complexes originated from monoclonal (Jurkat) vs polyclonal (human PBMC) sources. We used size-exclusion chromatography (SEC) to obtain fully assembled TCR/CD3 complexes (fractions 6–8, Superose 6 column [further described below]). Such complexes were subjected to immunoprecipitation with the anti-human CD3ɛ MAb UCHT1, followed by reducing SDS-PAGE/WB analysis with 7F18 probe. We found that from either cell source, MAb UCHT1 captured a similar amount of co-associated TCRα relative to CD3ζ (Fig. 2D). This observation was based on using semi-quantitative analysis of pixels to estimate the level of TCRα protein detected by 7F18 present within the expected molecular weight range (Fig. 2D, dashed box; polyclonal PBMCs can express TCR alpha-chains of various lengths within this MW region). Because monoclonal (Jurkat) and polyclonal (PBMC) sources of complexes showed a similar ratio of TCRα/CD3ζ, we propose that 7F18 can bind to most (or all) human TCR alpha-chains. In summary, we conclude that MAb 7F18 is useful for detecting human TCRα by WB, although MAb 7F18 was not useful for probing cell surface TCRs on live cells by FACS (Fig. 2E).
To visualize maturation and multiprotein assembly of human TCR alpha-chain, we used SEC followed by reducing SDS-PAGE/WB to provide a two-dimensional analysis that first separated proteins and multiprotein complexes by native MW (SEC), and then by reduced, denatured MW (reducing SDS-PAGE/WB). Human TCRα was assessed by WB with 7F18, initially comparing Jurkat and JRT3 cell lines, the latter being a mutant derivative of Jurkat in which TCRβ expression is lost and TCR/CD3 assembly is inhibited.(31) For Jurkat cells, we observed that the TCRα band at 40–45 kDa displayed peak elution in early fractions (higher native MW), indicative that the protein is assembled into multiprotein complexes, as is expected for TCR/CD3 (Fig. 3, upper panel). However, for JRT3 cells, a different major 7F18 band appeared in later fractions (lower native MW), where unassembled proteins are expected. Furthermore, that band was also of lower reduced, denatured MW, matching that expected of hypo-glycosylated, immature, unassembled TCRα(15) (Fig. 3, lower panel). These observations are consistent with a model in which human TCRα assembly and glycoprotein maturation matches that which has been reported for mouse TCRα. In this model, Jurkat TCRα-containing complexes elute from SEC in fractions containing fully assembled TCR/CD3 complexes in which TCRα has undergone glycoprotein maturation; in contrast, without TCRβ, JRT3 cells contain many unassociated alpha-chains that neither complete subunit assembly nor finish glycan addition and processing.
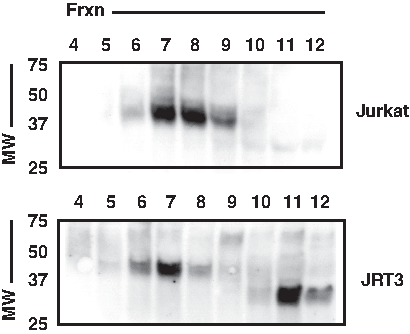
Both mature/assembled and immature/unassembled forms of TCRα are detected by MAb 7F18. A 2D analysis of TCR-containing cell lysates was performed using size-exclusion chromatography (SEC) followed by reducing SDS-PAGE/WB. Lysates of 20×106 Jurkat or JRT3 cells were subjected to the analysis, culminating in WB using MAb 7F18. In Jurkats, where normal TCR/CD3 expression occurs, the major band matching the expected MW of mature TCRα was found in high native molecular weight fractions (SEC fractions 6–9), indicative of multiprotein assembly. The major band's reduced, denatured migration matched that predicted of mature TCRα glycoprotein. In contrast, in JRT3 cells, which lack TCRβ, a different major band appeared of lower native MW (SEC fractions 11–12) and lower reduced, denatured MW, consistent with the migration expected of immature, unassembled TCRα.
Finally, since properly folded TCRs are expressed as disulfide-linked αβ heterodimers, we analyzed human PBMC lysates under both reducing and non-reducing conditions. Whereas mature, reduced human TCRα migrates at ∼40–45 kDa by SDS-PAGE, under non-reducing conditions disulfide-linked αβ migrates ∼80 kDa. Using SEC fractionation followed by SDS-PAGE/WB, we found that 7F18 specifically detected human TCRα at ∼40–45 kDa under reducing conditions, and at ∼80 kDa under non-reducing conditions (Fig. 4, lanes 6–8 show major band), as predicted. Primary PBMCs also possessed a low amount of TCRα that was smaller than the major band in terms of both native and denatured MW (Fig. 4, SEC fractions “12” in both blots). The existence of this minor band is consistent with the idea that it represents an immature glycoprotein (thus its lower MW by SDS-PAGE) that has not been assembled into a multiprotein complex (thus its apparent MW by SDS-PAGE remains constant, regardless of reducing/non-reducing conditions). We conclude that the MAb 7F18 specifically detects the human TCRα constant region by SDS-PAGE/WB, and it appears capable of detecting both mature/assembled and immature/unassembled TCRα protein.
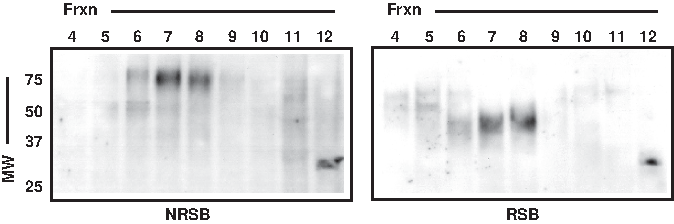
Human TCRα can be detected by MAb 7F18 when SEC-fractionated primary human T cell lysates are prepared in either reducing sample buffer (RSB) or non-reducing sample buffer (NRSB). Human PBMC lysate prepared from 50×106 cells was subjected to SEC followed by SDS-PAGE/WB analysis with MAb 7F18. When non-reducing conditions were maintained at the SDS-PAGE step, the major 7F18 band was detected at ∼80 kDa, as predicted for TCRα that is disulfide-linked to TCRβ (left). However, when reducing conditions were introduced at the SDS-PAGE step, the major 7F18 band was detected at ∼40–45 kDa, as predicted for reduced TCRα (right).
Concluding Remarks
We report the generation of MAb 7F18 specific for the constant region of human TCR alpha-chain. The present data show that 7F18 is capable of detecting human TCRα by both traditional SDS-PAGE/WB, as well as 2D analysis (SEC followed by SDS-PAGE/WB), under reducing or non-reducing conditions. Previous anti-human TCR MAb efforts have concentrated on specificity for the variable regions composing the TCR antigen-binding domains. As such, anti-V region MAbs were not elicited to display pan-reactivity with all human αβ TCRs, and thus provide limited use for human TCRα biochemical characterization. However, the specificity observed with 7F18 for human TCRα constant region makes it suitable for applying important biochemical analyses to TCR clones of any specificity. We propose that 7F18 will likely enable human TCR analyses similar in scope to those that MAb H28-710 has provided for mouse TCR studies.
Footnotes
Acknowledgments
This work was funded by the Mayo Clinic (AGS, DG). We thank Zhenjun Chen and James McCluskey (University of Melbourne, Australia), for kindly providing JRT3.huCD8ab.LC13 cells and retroviral vectors containing LC13 sequences,(23) from which we separately cloned alpha and beta sequences for use in transient transfection experiments. We also thank Balbino Alarcón (CBMSO, Universidad Autónoma de Madrid, Spain) for providing serum 448 (anti-CD3-zeta),(26) and Thomas Beito (Mayo MAb Core Facility) for determining the isotype of 7F18.
Author Disclosure Statement
The authors intend to make MAb 7F18 available to interested commercial entities by non-exclusive agreement.