
Abstract
Octopamine (OA) is one of the biogenic monoamines in the housefly, which acts as an important neurohormone in the physiological process of this pest. In this study, a new hapten of OA was synthesized via aldol condensation. With the hapten, monoclonal antibodies (MAb) were generated and their characterizations were investigated. An indirect competitive enzyme-linked immunosorbent assay (icELISA) based on MAb 3C11-E3 was established, which required simple sample pre-treatments and had low cross-reactivity with OA structural analogise. The half maximal inhibition concentration (IC50) and the detected range (IC20-IC80) of the icELISA were 128 ng/mL and 12-1438 ng/mL, respectively. Average recoveries of OA ranged from 73 to 129% in the housefly.
Introduction
D
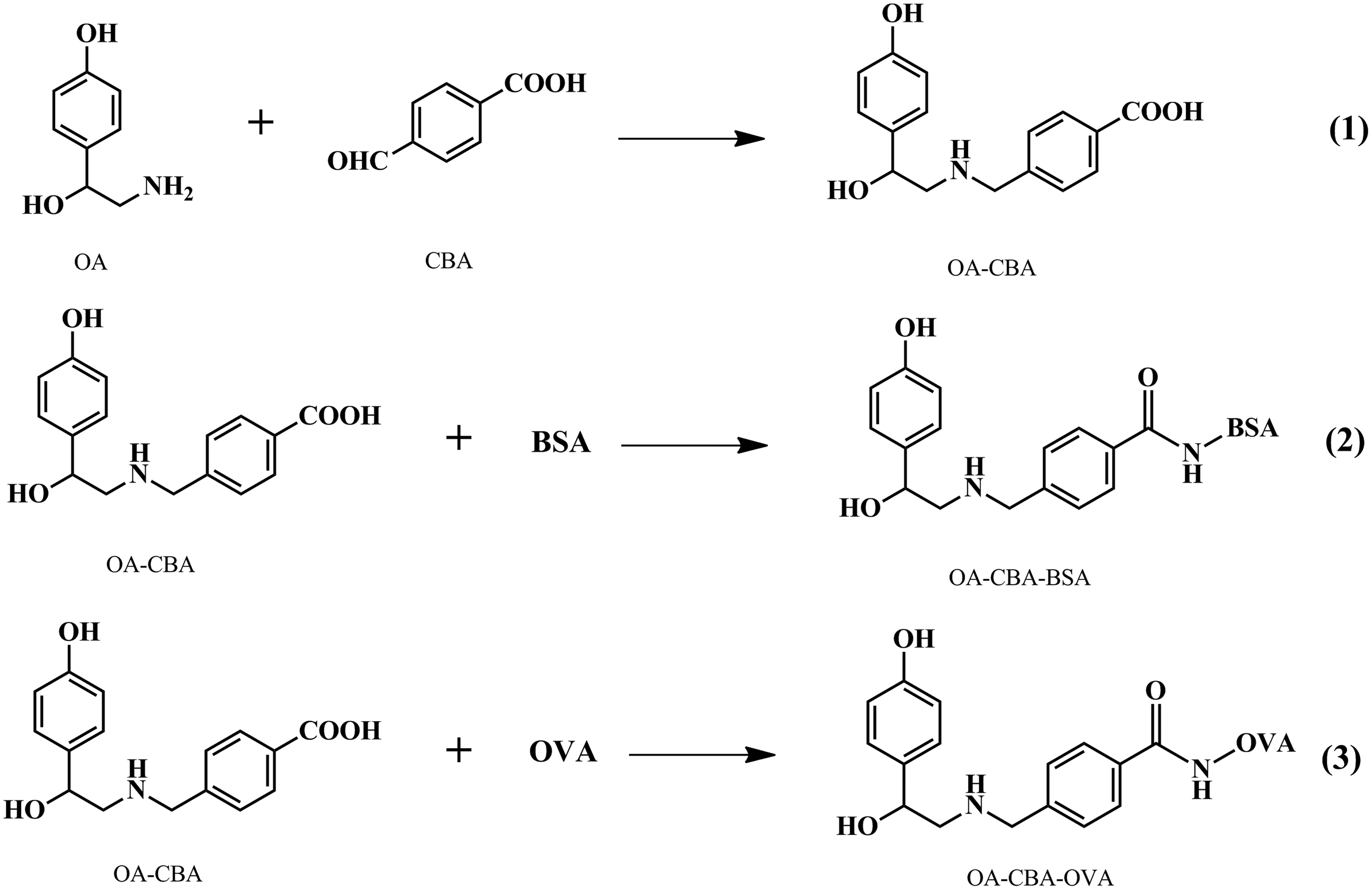
Reactions schemes used to synthesize the OA derivatives and conjugates. OA-CBA was used as the hapten; OA-CBA-BSA and OA-CBA-OVA were used as immunogen and coating antigen, respectively.
Nowadays several techniques have been used for OA analysis in agricultural insects. High-performance liquid chromatography (HPLC) is one of the most frequently used techniques.(2–6) Other instrumental analysis methods, such as gas chromatography conjugated negative ion chemical ionization mass spectrometry(7) and micellar electrokinetic capillary chromatography,(8) were also applied to detect OA in insect tissues. In addition, a radioenzymic assay was used by Davenport and Wright to measure OA in moth.(9) Barron and colleagues used a radioisotope assay to analyze the distribution and dynamics of OA in honeybees treated with 3 H-octopamine.(10)
Although the methods mentioned above are highly sensitive and accurate, they are expensive, time-consuming, and require complicated sample preparation. Enzyme-linked immunosorbent assay (ELISA) is one of the fastest, most sensitive, selective, and cost-effective analysis methods. It requires a relatively simple sample preparation. Both monoclonal antibodies (MAbs) and polyclonal antibodies (PAbs) against OA have been reported for immunohistological studies in insect tissues.(11–14) However, the MAbs and PAbs reported had relatively lower specificity to OA and have not been used for quantitative analysis. In the present study, a new MAb (3C11-E3) against OA was produced with a newly synthesized hapten. An indirect competitive ELISA (icELISA) based on 3C11-E3 was developed and optimized for the analysis of OA in the housefly.
Materials and Methods
Hapten synthesis and conjugate preparation
The hapten OA-CBA (4-((2-hydroxy-2-(4-hydroxyphenyl)ethylamino)methyl)benzoic acid) was synthesized via aldol condensation(15) (Fig. 1). Briefly, OA (200 mg) and NaOH (42 mg) dissolved in 1 mL methanol were added to a solution of 4-CBA (158 mg) and NaOH (42 mg) dissolved in 5 mL methanol. The mixture was incubated at 60°C for 4 h with stirring to obtain a yellow color. Thin-layer chromatography was used to monitor the reaction progress. After an ice bath, NaBH4 (50 mg) was added to the reaction product and the mixture was stirred for another 30 min. While the yellow color faded, concentrated HCl was added drop-wise to remove the unreacted NaBH4 and the solution was adjusted to pH 5.0. Acidification of the solution resulted in precipitation of a white solid, which was subsequently centrifuged, washed with methanol, and recrystallized. The crystal (OA-CBA) was dissolved in DMSO and analyzed with 1H-nuclear magnetic resonance (1H-NMR) spectra. The chemical shift was relative to 1H-NMR (300MHz, DMSO): 2.89 (m, CH2, 2H); 3.16 (s, OH, 1H); 4.25 (s, CH2, 2H); 4.91 (m, CH, 1H); 6.00 (s, OH, 1H); 6.73 (d, aromatic-H, 2H); 7.12 (d, aromatic-H, 2H); 7.67 (d, aromatic-H, 2H); 7.95 (d, aromatic-H, 2H); 9.48 (m, NH; COOH, 2H).
OA-CBA was conjugated with BSA and OVA (Fig. 1) via the active ester method.(16) To a solution of hapten OA-CBA (10 mg) and NHS (6 mg) dissolved in 1 mL DMF, 8.6 mg DCC were added. The mixture was gently stirred at room temperature for 1 h and then centrifuged at 10,000 g for 10 min. An aliquot of 250 μL and 255 μL of the supernatant was slowly added into BSA solution (30 mg in 2.0 mL carbonate buffer) and OVA solution (20 mg in 2.0 mL carbonate buffer) in 1 h, respectively. After the two solutions were stirred overnight at 4°C, the conjugate products OA-CBA–BSA and OA-CBA-OVA were dialyzed and then diluted to 1 mg/mL.
Method of immunization
The protocol of immunization was similar to that previously described.(17) Five female BALB/c mice, 6 weeks old, were initially injected with 100 μg of OA-CBA-BSA emulsified with an equal volume of complete Freund's adjuvant. Two subsequent injections were carried out at 2-week intervals using Freund's incomplete adjuvant. One week after the third injection, mice were eye-bled and sera were tested via ELISAs. The best-performing mouse was selected for hybridoma production and boosted with 100 μg of OA-CBA-BSA 3 days before fusion.
Antibody preparation
MAbs were prepared by fusing mouse spleen cells and murine SP2/0 myeloma cells according to the procedure previously described.(18) The hybridoma supernatants were tested by ELISAs for positive clones. The resulting clone was expanded in mice for ascites production and the MAb was purified by ammonium sulphate precipitation. The immunoglobulin isotype was determined with a mouse antibody isotyping kit. The titer was determined by indirect ELISA.
Establishment of icELISA
A 96-well microtiter plate was coated with 100 μL/well of OA-CBA-OVA in carbonate buffer at 37°C for 3 h. After being washed with 200 μL/well of PBST three times, 50 μL/well of analytes or standard in PBSTG were pipetted, followed by the addition of 50 μL MAb solution diluted in PBSTG. The plate was incubated at 37°C for 0.5 h and then washed with PBST as above. 100 μL of the goat anti-mouse IgG-peroxidase conjugate in PBSTG were added to each well followed by incubation at 37°C for 0.5 h. After being washed again, 100 μL/well of OPD substrate solution were added. The reaction was stopped by adding 50 μL of 2 M H2SO4. Absorbance was read at 492 nm. All data were analyzed by Origin 8.0 software.
Cross-reactivity studies
The cross-reactivity (CR) of the established icELISA assay was determined against several OA analogise, including adrenaline, 5-hydroxytryptamine, dopamine, norepinephrine, and tyrosine. CR was calculated as shown in Equation (1):
Spiking tests
Laboratory-maintained houseflies were used for recovery studies. An amount of 500 mg adult housefly sample was ground, and then divided into aliquots of 20 mg. OA stock solution was diluted to 20, 40, 80, 160, and 320 μg/mL in PBS, and then 10 μL of the standard solutions were added to the aliquots of ground sample, respectively, followed by gentle vortexing. A housefly sample without OA added was used as blank controls. After being dissolved in 1.0 mL of PBS, each sample was extracted at 4°C for 0.5 h, and subsequently centrifuged at 8000 rpm for 10 min. Each supernatant was detected with icELISA in triplicate.
icELISA analysis of housefly samples
Adult houseflies (resistant and sensitive strains to Imidacloprid, Malathion, Propoxur, Spinosad, and Beta-cypermethrin) were weighed and ground. The following extraction and detection of OA was conducted as the spiking tests.
Results and Discussion
Hapten synthesis and hapten-protein conjugation
OA is a small (MW 153) and simple molecule (Fig. 1), making it difficult to obtain specific and high affinity antibody as hapten. It presents in insects at a relatively low level, which represents a challenge for OA hapten design to obtain antibodies to develop a sensitive ELISA. Although we tried to synthesize hapten with glutaradehyde as previously described,(12) the antibodies produced have high affinity against glutaradehyde-OA, but low affinity against free OA. This hapten synthesization method only provoked antibody suitable for immunocytochemistry study, but not for ELISA to detect free OA. In the present study, hapten OA-CBA was synthesized via aldol condensation. It contained a terminal carboxyl acid for conjugation with carrier proteins. Then we conjugated OA-CBA with BSA and OVA via the active ester method as immunogen and coating antigen.
Monoclonal antibody generation and characterization
After the third immunization, blood was obtained from mice to determine titers and inhibition against OA. The mouse producing antisera of the best inhibition was then used to collect spleen cells for hybridoma cell production. Four clones secreting MAbs against OA were obtained after limiting dilution. One clone, designed as MAb 3C11-E3, with the best inhibition with OA was expanded for ascites production. The titer of the ascites was 1:16,000. MAb 3C11-E3 is an IgG1 isotype.
icELISA development and evaluation
Optimum concentrations of the coating antigen (0.25 μg/mL), purified MAb (50 ng/mL), and the IgG-HRP conjugate (1 μg/mL) for icELISA were screened by checkerboard titration. Figure 2 shows a standard curve of OA by the icELISA under the optimized conditions. The IC50 value of the icELISA was 128 ng/mL and calibration range (20–80% inhibition) was from 11 to 1438 ng/mL of OA. The specificity of MAb was evaluated with OA analogues (Table 1). The results showed high cross-reactivity with adrenaline (440%), whereas there was little with dopamine (0.4%) and norepinephrine (0.9%), and no cross-reactivity with 5-hydroxytryptamine and L-tyrosine. Although the cross-reactivity with adrenaline was high, adrenaline has not been found in invertebrates. So the high cross-reactivity would not affect the analysis of OA in houseflies. Compared with the reported MAb(11,13) (63% to epinephrine, 21% to tyramine, 8% to norepinephrine, dopamine and serotonin showed 0% inhibition), 3C11-E3 showed lower cross-reactivity with structural analogues, but polished with non-conjugated OA.

Typical icELISA standard curve of inhibition by OA based on MAb 3C11-E3.
CR, cross-reactivity.
The housefly samples were extracted at 0.5, 1.0, 2.0, and 4.0 h, respectively, to detected OA content for optimal recoveries. The icELISA data show that there was no difference in recoveries with an extraction time between 0.5 h and 4 h. So in the following analysis, we used the extraction time of 0.5 h. Table 2 shows that average recoveries of OA fortified in houseflies ranged from 73 to 129%. In addition, we fortified OA in German cockroach with the same concentrations; the average recoveries ranged from 82 to 104% by icELISA.
Data were averages of three determinations.
icELISA analysis of housefly samples
We used the established icELISA to determine OA content in houseflies of resistant or sensitive strains to Malathion, Propoxur, Imidacloprid, Spinosad, and Beta-cypermethrin, all of which are widely used insecticides, the latter three acting as insect neurotoxins. The result (Fig. 3) showed that OA was more in the sensitive strain of houseflies to Imidacloprid, Spinosad, and Beta-cypermethrin compared to resistant strain houseflies. But there was no difference in the two strains of houseflies to Malathion and Propoxur. In other words, the resistant strain houseflies may strengthen resistibility partly by reducing the content of OA. According to a previous report, OA could induce the increase of cyclic adenosine monophosphate (cAMP) content in insects, and then the cAMP would act as a second messenger to further regulate physiological responses.(19) The lower content of OA in a resistant strain housefly would control the cAMP level to make the housefly less sensitive to the treatment of some neurotoxins. We used the female and male houseflies for further studies, respectively. The results indicated that the contents of OA in the resistant strain (2.20 ng/mg in female housefly, 3.18 ng/mg in male housefly) were still lower than that in the sensitive strain (4.89 ng/mg in female housefly, 4.72 ng/mg in male housefly) after being treated with Spinosad. We also assayed the OA in the head and body of male and female houseflies, respectively. The conclusion was consistent with Roeder's research(1); more OA was present in housefly heads (13.58 ng/mg in female housefly, 15.86 ng/mg in male housefly) than in their bodies (5.50 ng/mg in female housefly, 5.16 ng/mg in male housefly).

Contents of OA in resistant and sensitive strain houseflies.
Conclusions
To our knowledge, this is the first report describing an OA hapten synthesization method via aldol condensation. The MAb produced by OA-CBA-BSA had higher affinity to OA than that previously published and was successfully used to develop an icELISA for the analysis of OA in the housefly. The IC50 value of icELISA was 128 ng/mL. The working range (20–80% inhibition) was from 11 to 1438 ng/mL. The MAb 3C11-E3 had weak cross-reactivity with other metabolic products in insects. Spiking tests of OA in housefly samples showed that the developed icELISA was a quick, reliable, and sensitive method.
Footnotes
Acknowledgments
The authors would like to thank Professor Qingxiao Li (Department of Molecular Biosciences and Bioengineering, University of Hawaii) for valuable suggestions.
Author Disclosure Statement
The authors have no financial interests to disclose.