
Abstract
Hemophilia A is an X-linked disorder affecting 1 in 10,000 males. The disease is caused by a defect or mutation of factor 8 or 9. Human factor 8 gene (hFVIII) is a relatively large gene consisting of 26 exons and approximately 2,351 amino acids with a length of 9 Kb mRNA. Expression of hFVIII in mammalian milk is becoming a widespread strategy for high-level production of hFVIII because of the most complex post-translational modifications. The aim of this study was the cloning and expression of hFVIII in mammary glands of two transgenic mice. To obtain a recombinant plasmid, first a plasmid carrying an FVIII gene fragment (pCMV6-hFVIII) was digested by EcoRI-SalI restriction enzymes and then the fragment was purified from agarose gel and inserted into a pUCWAP7 vector carrying a tissue-specific promoter (mWAP 4.1 kbp). After that, it was isolated by agarose gel and transferred into the murine zygotes by standard microinjection methods. Methods for expression of recombinant FVIII RT-PCR and ELISA were studied. The results show the successful expression of factor FVIII gene and its product in the mouse mammary glands.
Introduction
H
In the past, plasma-derived therapeutics has been used to treat hemophiliacs but recently, concerns over HIV, hepatitis, and most recently prions have made finding other sources for this therapeutics particularly attractive. Factor VIII production using recombinant DNA technology in cell culture has been explored with varying degrees of success. Unfortunately, the available amounts do not meet worldwide demand, especially for prophylaxis, and the obtained rhFVIII shows considerable variability in post-translational modifications.(7) On the other hand, the use of transgenic animals as bioreactors to produce pharmaceutical biologics is maturing with several derived proteins in or about to enter clinical trials. The transgenic animals are not only cost-effective bioreactors, but with the complex secretory cell types and organs of the mammalian organism, researchers can perform much more complicated protein modifications than simply cultured cells.(8) By employing targeted gene transfer (e.g., using mammary gland-specific regulatory sequences fused with hFVIII gene), it is possible to govern the expression to occur exclusively in the mammary gland and hence the gene product is being ultimately secreted in milk.(7) Animals automatically supplement their body fluids with fresh nutrients, remove waste products, reliably regulate their internal temperature and pH, and are resistant to pathogens. The mammary gland probably is the most promising target tissue because it produces large amounts of protein in a temperature-regulated fluid that may be collected daily in a non-invasive fashion.(10)
Materials and Methods
DNA constructs
The pUCWAP7 construct was assembled by altering the Kpn I site of pUCWAP6(12) by introducing EcoRI and SalI sites immediately downstream of the 4.1 kbp WAP (whey acidic protein) promoter and ahead of the 1.7 kbp of mouse WAP 3′ UTR containing the coding of the polyadenylation signal to produce pUCWAP7. The mWAP is a tissue-specific promoter and contains genetic codes, which direct the expression of the foreign protein in the mammary gland during lactation. The WAP clone (pUC Not I+) containing ∼4.1 kb of WAP 5′ promoter region, and ∼1.7 kb of WAP 3′ UTR, including the flanking 3′ end, was modified from a 7.2 kb genomic WAP fragment. For pUCWAP7-hFVIII construction, pUCWAP7 plasmid was restricted by EcoRI and ligated by t4-ligase with an EcoRI-SalI fragment of pCMV6-XL4/nM_000132 containing the full factor VIII gene (Origene, Rockville, MD). Construct DNA was diluted to 5 μg/mL in microinjection buffer (10 mM Tris-HCl, 0.1 mM EDTA) to produce monogenic lines or mixed in equal molar amounts for co-microinjection at a final concentration of 5 μg/mL.
Generation of transgenic mice
Generation of transgenic mice was performed using the general method of Hogan and colleagues(10) for embryo collection and Canseco and colleagues(11) for embryo microinjection. Briefly, female CD-1 mice and BALB/c mice approximately 25–29 days of age were superovulated with intraperitoneal injections of 10 IU pregnant equine serum gonadotropin followed by 5 IU human chorionic gonadotropin (hCG) 48 h later and placed with males. Zygotes from mated mice were collected in M2 medium 21 h after hCG and microinjected with 1–4 pL of DNA solution at 5 μg/mL. Surviving embryos were transferred to oviducts of pseudo-pregnant females (30–100 embryos per recipient) with the number transferred reflective of survivability studies done in culture for each preparation of DNA.
Identification of transgenic mice
Total DNA was isolated from newborn murine tail biopsies. As negative control, DNA isolated from non-transgenic mice was used. PCR amplification was performed using 100 ng of DNA derived from tail biopsies with hFVIII gene-specific primers: 5′-GTAGAC AGC TGT CCA GAG GAA-3′ and 5′-GAT CTG ATT TAG TTG GCC CAT C-3′. The reaction mixture underwent initial denaturation process at 94°C for 5 min, followed by 30 cycles at 94°C for 30 s, 64°C for 30 s, and 72°C for 35 s. The final extension was performed at 72°C for 10 min in a thermocycler (PaqLab, Erlangen, Germany). The PCR fragments were run in 2% agarose gel and were visualized by ethidium bromide staining.
Collection and processing of mouse milk
Milk from transgenic and control mice was directly collected into 1.7 mL centrifuge tubes by vacuum aspiration, with 200 μL of milk removed, added to 200 μL of milk buffer (50 mM Tris and 200 mM EDTA [pH 8.5]) and frozen at −90°C. Milk samples were defatted by centrifugation at 15,000 g in a bench-top centrifuge for 20 min at 4°C. After skimming, the resulting clarified milk (prepared whey) was stored at −90°C until analysis.
ELISA
Secretion of rhFVIII into the transgenic milk was determined quantitatively using an ELISA kit (Asserachrom FVIII:Ag, Diagnostica Stago, Paris, France) according to the manufacturer's manual.
Coagulation assay
The clotting assay for the hfVIII level was performed on a thromboScreen 400c optical coagulometer using the reagents kit Factor VIII test (NPO Renam, RF) according to the kit manufacturer's protocol.
Reverse transcription PCR
Total RNA was extracted from tissue samples of mammary glands from transgenic mice on day 35 of first lactation according to the manufacturers' protocol. cDNA was synthesized from the isolated RNA, pre-treated with RNase-free DNase, by AMV-reverse transcriptase. The synthesized hFVIII-cDNA was subsequently analyzed by amplification of two fragments, using two specific primer pairs, namely p1-F (5′-AGT CCT GAA GCT AGA TCT CTC TCC-3′), p1-R (5′-ATA AGA ATG CGG CCG CAA TAT GGA GAG AGA TCT AGC-3′), p2-F (5′-ATT TGG CGG GAG GAA T GC CTT ATT GGC G-3′), and p2-R (5′-ACA CCT CGA GTC AGT AGA GGT CCT GTG CCT CG-3′) from the hFVIII coding region.
Results
Analysis of transgenic animals
PCR analysis of potentially transgenic mice, which developed from embryos microinjected with pUCWAP7-hFVIII constructs and hFVIII-specific PCR product of 578 bp, was obtained (Fig. 1).

Electrophoresis of PCR products for the hFVIII transgene integration into genome of mice. Lane 1, DNA marker (100 bp DNA ladder); lanes 2 and 4, transgenic mice (female CD-1 and BALB/c, size of hFVIII-specific PCR product is 578 bp); lane 3, DNA template (negative control); lane 5, non-transgenic mice.
The results of integration rates of pUCWAP7-hFVIII construct in two mice are summarized in Table 1. The results show that pUCWAP7-hFVIII constructs frequently integrated into the embryonic genomes of mice. Furthermore, mendelian transmission was observed in 10 of the 17 CD-1 mice that were tested.
Concentration of rhFVIII in mice milk
ELISA results identified that several lines secreted detectable amounts of rhFVIII into their milk and the concentrations of rhFVIII detected depended on generation and lactation day in the milk of transgenic female mice ranged between 5 and 130 μg/mL. Also, no measurable concentrations in non-transgenic mouse milk were detected.
Activity measurement of hFVIII
The highest activity of rhFVIII was detected in transgenic BALB/c females from the F2 generation on second lactation (475 mU/mL), which corresponds to 47.5% of the level of normal human plasma. On the other hand, the coagulation activity of concentrated samples was five times higher than that of transgenic BALB/c females from F3 generation.
RT-PCR analysis
The presence of the rhFVIII transcript in the selected transgenic female mice were shown by the PCR amplification of two fragments from FVIII cDNA, reversely transcribed from transgenic mouse mRNA. As the electrophoresis pattern of the RT-PCR products indicates, bands of the appropriate size support the transcription of hFVIII in the transgenic mice (Fig. 2).
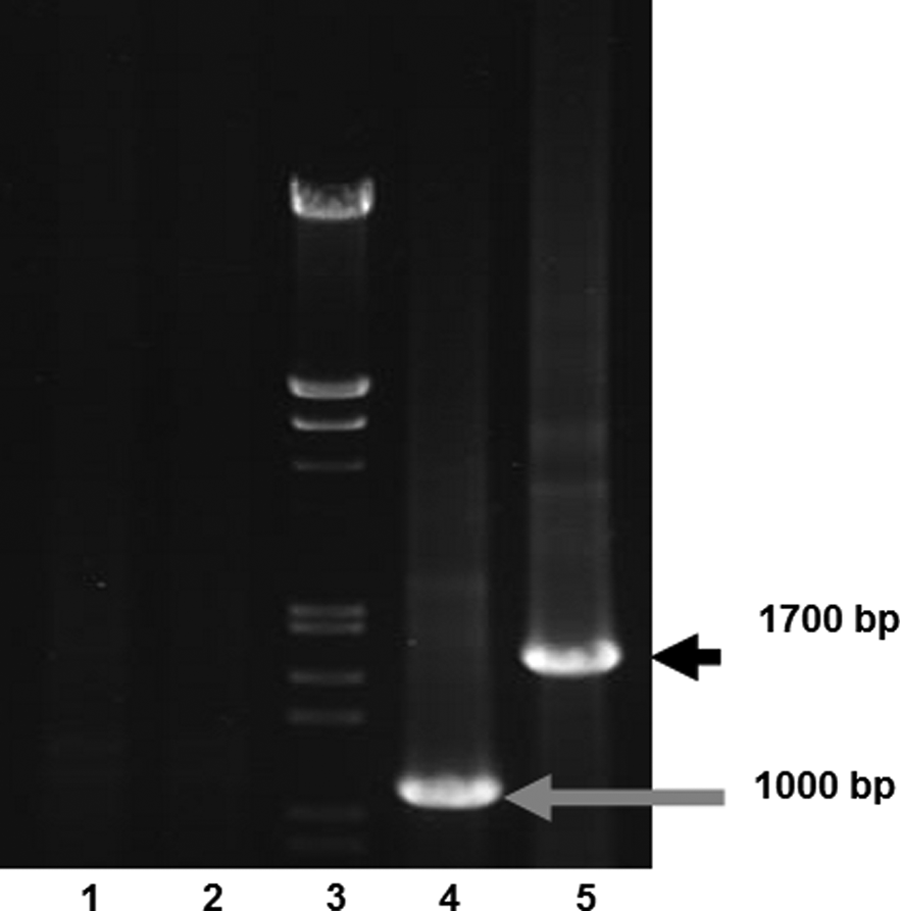
Electrophoresis of PCR products amplified from the reverse transcribed total RNA. In each case, the prepared RNA was treated with RNase-free DNase. Lane 1, non-transgenic mice using p1-F/p1-R primers; lane 2, transgenic mice using total RNA pre-treated with RNase-free DNase as template and p1-F/p1-R primers; lane 3, DNA size marker (λ-phage DNA digested with HindIII/EcoRI); lane 4, transgenic mice using p2-F/p2-R primers; lane 5, transgenic mice using p1-F/p1-R primers.
Discussion
Treatment of hemophilia A by replacement therapy is done by infusion of either human plasma derived FVIII or recombinant FVIII. But FVIII replacement remains a very expensive therapeutic with the average patient using up to $100,000 worth of hFVII concentrate per year. In spite of production of hFVIII in cultured cells, investigations on the overproduction of hFVIII in transgenic animals are in progress in several laboratories throughout the world.(8–10) In recent years it has been shown that establishing the use of live bioreactors for the expression of human genes in the mammary gland of transgenic animals is one of the most cost-effective ways for the production of valuable recombinant factor VIII.(9) Here we have reported a successful production of biologically active hFVIII in our laboratory.
Wild-type FVIII is unstable under most conditions. In vivo this instability is a characteristic of the way in which its activity is regulated. Under cell culture conditions, judicious gene splicing combined with careful control of the growth media can allow production of stable FVIII. Cell culture, however, has relatively low productivity and scale-up to industrial-level production is difficult. In the milk of transgenic animals, high production is possible but the generation of high specific activity protein is difficult and costly. The complex milk environment includes proteases that degrade the FVIII and caseins that sequester it and complicate its purification. All of these complications combine to make the production of large amounts of high-specific activity FVIII a goal that is thus far unattained. The production of recombinant proteins in the mammary glands of transgenic animals is dependent on gene promoters used in transgene constructs. Most of the studies have been carried out with the ovine β-lactoglobulin, bovine α-lactalbumin, caprine β-casein, or mouse whey acidic protein (WAP) promoter.(13) The mouse WAP promoter has been used in basic biological studies, as well as in the synthesis of pharmacologically active human proteins, to direct the expression of heterologous genes to alveolar epithelial cells.(14,15) In this study, two lines of transgenic mice from different mice were developed to express the hFVIII gene in the mammalian glands under control of milk mWAP promoter. Our PCR results confirmed the mWAP-hFVIII transmission in several generations of transgenic females. The founding females secreted hFVIII in their milk. Our results confirm the production of a large amount of recombinant biologically active hFVIII in the mammary glands of transgenic mice.
The lactation specificity of the regulatory regions used in the control of mammary gene expression in transgenic animals is very important in case of expression of foreign biologically active proteins, because these proteins may exhibit their biological functions in the animal if secreted prior to the tight junction formation in the mammary epithelial cells.(10) The effects of expressing growth factors, hormones, oncogenes, ECM components, and receptors on mammary gland development and differentiation have been well documented.(17) The results show the transgenic mice expressing recombinant human protein into mammary gland, under mouse WAP promoter, exhibiting defects in lactation and impaired mammary gland development.(18) The WAP promoter was shown to be less efficient than the β-lactoglobulin promoter at driving the overexpression of recombinant human protein into milk.(19) However, in our transgenic mice we confirmed high levels of tissue-specific expression of hFVIII in the mammary glands. On the other hand, hFVIII has been produced in the milk of transgenic animals. Thus far hFVIII have expressed in the milk of transgenic swine and transgenic sheep.(8,9) Both research groups had very low specific activity, with 0.33 U/μg obtained in pigs and 0.005 U/μg obtained in sheep. Antigen analysis via ELISA showed FVIII levels of approximately 5 μg/mL obtained in pigs with Niemann detecting only trace amounts in transgenic sheep milk. The factor VIII in plasma typically has an activity of 5 U/μg with 200 ng/mL being a typical FVIII concentration. When FVIII is expressed in milk, it will suffer from the same problems as that observed in cell culture along with several other hindrances unique to the milk environment. Unfortunately, it is secreted into milk with low specific activity due in part to the labile, heterodimeric structure that results from furin processing of its B domain. Recently, in one study, it was expressed biologically in active rFVIII in the milk of transgenic mice through targeted bioengineering.(20) The transgenic mice had >450-fold higher IU mL (-1) than previously reported in cell culture for rFVIII and exhibited similar binding to plasma-derived VWF as therapeutic-grade rFVIII. Also interestingly, intravenous infusion of this transgenic mouse corrected the bleeding phenotype of hemophilia A mice. In total, a higher rhFVIII concentration in our transgenic mice was detected as compared to previously reported rhFVIII concentration in mammary glands of other transgenic animals. Furthermore, transgenic mice offer rapid evaluation of both synthesis rate and functionality due to their short generation intervals and ease of genetic manipulation and multiple offspring per litter, thus allowing production of a biologic before investigation in larger animals. However, a high variability between generations and individual transgenic mice in rhFVIII concentration and biological activity is observed, and for commercial use an individual evaluation and selection of transgenic animals are required.
Footnotes
Acknowledgments
This research was supported with funds from the Shahr-e-Qods Branch of Islamic Azad University.
Author Disclosure Statement
The authors have no financial interests to disclose.