
Abstract
Recently we reported the development of a highly specific murine monoclonal antibody (ERG MAb 9FY) against the ERG oncoprotein. ERG is expressed in over half of all prostate cancers (CaP) as a result of specific gene fusions involving ERG and the androgen regulated TMPRSS2 promoter. ERG MAb 9FY has been extensively used in the evaluations of CaP. Increasing use of ERG MAb in CaP has prompted us to characterize the precise ERG epitope it binds to and to define the molecular basis of its specificity to ERG. The 9FY antibody binds to an epitope formed by amino acid residues 42-66 of the ERG protein. To determine the key residues involved in 9FY binding, experiments were carried out using a combination of approaches including overlapping peptides, alanine scanning mutagenesis, ELISA, and immunoblot assays. Analysis of both overlapping and variant peptides harboring truncations of amino acids revealed that a minimal epitope of eight residues (RVPQQDWL) is sufficient for binding to the 9FY antibody. In order to further identify key residues that mediate the binding of the antibody to ERG protein, a 14-residue peptide (P23) with optimal reactivity was subjected to alanine scanning mutagenesis. Alterations to residues QQDW were found to eliminate binding to the antibody, while residues (R50 and L57) were found to contribute to the binding of the antibody. Further experiments showed that peptide P23 competed effectively with ERG protein for binding 9FY. On the other hand, peptides with alanine substitutions for residues Q53 and W56 (P27 and P30, respectively) failed to interfere with binding. These data provide new information about a minimal epitope (RVPQQDWL) within amino acid residues 42-66 of the ERG protein that is recognized by MAb 9FY, which may aid in the diagnosis and also development of antibody based therapeutics against prostate and other cancers showing ERG overexpression.
Introduction
H
A major goal in CaP, as with other cancers, is to define protein and antibody markers that may facilitate early detection, distinguish indolent from aggressive diseases, define treatment strategies, and allow follow-up of patients. The prostate-specific antigen (PSA) assay that is routinely used in the clinical screening of CaP has been shown to lack specificity.(2) Hence, additional biomarkers are clearly needed in this arena. The prevalence of ERG overexpression, resulting from the fusion of ERG coding sequences to the androgen-regulated gene transmembrane protease, serine 2 (TMPRSS2) promoter, has provided an impetus to develop monoclonal/polyclonal antibodies to the ERG protein for diagnostic and/or therapeutic purposes.(4–6) On this basis, mouse monoclonal antibodies (MAb) against ERG were developed in our laboratory. One of the ERG MAb clones, 9FY, recognized an epitope formed by the amino acid sequence GQTSKMSPRVPQQDW LSQPPARVTI, which corresponds to residue positions 42-66 in the ERG protein.(4) The 9FY monoclonal antibody was found to be highly specific for the detection of ERG protein in cell culture–based experiments and in human prostate cancer specimens.(4) The isotype analysis showed that the ERG MAb 9FY is IgG1 (k) and does not recognize other members of ETS family. In addition to 9FY, antibodies against ERG have also been developed by several commercial vendors including Epitomics, Santa Cruz and Origene, using peptides and recombinant proteins as immunogens.
Studies from our laboratory have shown that the 9FY antibody can be successfully used for detecting ERG protein in prostate cancer specimens by immunohistochemistry (IHC).(4,5) Similar observations were also reported for a rabbit monoclonal antibody raised against the C-terminal peptide of ERG as an immunogen.(6,7) Recent analysis of 180 whole mount prostate sections from age and pathologic stage matched from patients revealed that there is a striking difference in ERG expression in African American (AA) and Caucasian American (CA) patients.(8) A markedly greater frequency of ERG oncoprotein expression was noted between the index tumors in CA patients (63.3%) compared to AA patients (28.6%). Much lower frequencies (10–27%) of ERG alterations have been reported in studies from China, Japan, and India.(9–11)
Despite the development of murine and rabbit monoclonal/polyclonal antibodies against ERG protein, currently there is no information available about the precise epitopes recognized by these antibodies. This scenario has prompted us to undertake the characterization of the epitope formed by amino acids 42-66 of the ERG protein and identify the minimal sequences that are sufficient to mediate binding of the 9FY antibody by pep scan,(12) alanine scanning, ELISA, and immunoblot methods. The results presented in this study have enabled us to define a minimal epitope within amino acid residues 42-66 of the ERG protein and to identify key residues involved in the interaction between MAb 9FY and the ERG protein.
Materials and Methods
Cell culture
Human embryonic kidney (HEK) 293 cells were obtained from American Type Culture Collection (ATCC; Manassas, VA). Cells were cultured in Dulbecco's modified eagle medium (DMEM, ATCC) supplemented with 10% fetal bovine serum (FBS, ATCC) under humidified conditions at 37°C with 5% CO2. Medium changes occurred every other day, and cells were passaged as needed, when they reached confluency.
Transfection of expression plasmid DNA in cells
HEK293 cells were plated at a concentration of 4×106 cells in 15 mL of complete media (DMEM supplemented with 10% FBS) in 150 mm tissue culture dishes (Falcon, Corning, NY). Cells were incubated overnight at 37°C, so that they would be 50% confluent at the time of transfection. The next day cells were transfected with TMPRSS2-ERG plasmid DNA (4 μg) using Lipofectamine 2000 (Invitrogen, Grand Island, NY), as per the manufacturer's protocols. Cells were harvested 48 h post-transfection using cell scrapers, centrifuged at 4°C to pellet the cells, washed twice with PBS, and lysed.
Recombinant protein and peptides
Recombinant full-length ERG protein, produced in mammalian cells upon transfection of expression plasmid DNA, was purchased from Origene (Rockville, MD). Peptides were synthesized by a commercial vendor (Infinity Biotech, Aston, PA).
Enzyme-linked immunosorbent assay
ELISA procedures were carried out in NUNC 96-well flat-bottom Maxisorp plates (Thermo Scientific, Rockford, IL). Plates were coated with 500 ng/well of peptide, using 100 μL coating buffer (50 mM NaHCO3, pH 9.6). The reactions were carried out in duplicate. The plates were covered with microplate sealers (Pierce/Thermo Scientific, Rockford, IL) and incubated at 4°C overnight. The next day plates were washed four times with wash buffer (1X PBS+Tween-20, KD Medical, Columbia, MD) and blocked with 200 μL blocking buffer (StartingBlock, Thermo Scientific), covered, and incubated for 1 h at room temperature (RT). Plates were washed once with wash buffer, incubated with 100 μL MAb 9FY (licensed to Biocare Medical, Concord, CA) or other commercially available antibody (1 μg/mL serially diluted), and covered for 1 h at 37°C. Plates were again washed four times with wash buffer and then incubated with 100 μL of an HRP-conjugated secondary antibody against the primary antibody, diluted as per manufacturer's protocols (KPL, Inc., Gaithersburg, MD), and covered for 1 h at 37°C. Plates were washed four times with wash buffer, 100 μL of K-Blue Aqueous TMB substrate (Neogen, Lexington, KY) was added to the plates, and incubated uncovered for 30 min at RT. Sulphuric acid (100 μL of 2 N) was added to the plates post-incubation to stop the reaction. Plates were immediately read at 450 nm to measure absorbance. All dilutions of reagents were performed in ELISA diluent (20% NGS in 1X PBS with 0.1% Triton-X 100). For competition studies, the antibody (1 μg/mL, serially diluted) was first pre-incubated with the specific peptide (10 μg/mL) for 30 min at RT and then added to microtiter plates.
Western blot analysis
Cell pellets from transfection procedures were resuspended in 500 μL Mammalian Protein Extract Reagent (M-PER, Pierce/Thermo Scientific) containing protease inhibitor and phosphatase inhibitor cocktails and incubated on ice for 15 min. Suspensions were then sonicated using a water bath sonicator at 15 s pulses for 4 min at 35% amplitude. The lysate was centrifuged for 15 min at 4°C at 10,000 g. The clear supernatants were transferred to fresh centrifuge tubes and protein concentration was determined using Protein Assay Reagent (Bio-Rad, Hercules, CA). Five μg of protein were loaded onto 4–12% NuPAGE Bis/Tris Gels (Invitrogen) and run at 130 V for 75 min. The bands were transferred onto PVDF membranes at 25 V for 90 min. Membranes were blocked in blocking buffer (Li-CoR, Lincoln, NE) for 1 h at RT. Ten ng of each peptide were pre-incubated with MAb 9FY (1 ng/mL) or Epitomics-5115 (1 ng/mL) for 30 min at RT. The membranes were then incubated overnight with the peptide-MAb mixture at 4°C. The next day, the membranes were washed in Tris-buffered saline+Tween-20 (TBST) and then incubated with a secondary antibody against the primary antibody, diluted 1:20,000, for 1 h at RT. The membranes were again washed and imaged on the Odyssey Imaging System (LI-COR, Lincoln, NE).
Results
Identification of minimal epitope bound by ERG MAb 9FY antibody using overlap peptides
ERG MAb 9FY recognizes an epitope within a peptide (Fig. 1A) corresponding to amino acids 42-66 of the ERG protein (NP_891548.1). In order to characterize the minimal epitope within amino acids 42-66 sufficient for binding of the 9FY antibody, a series of five overlapping peptides (P1-P5) were synthesized. The 12 amino acid peptides (except peptide P5, which contains 13 amino acids) were designed with an overlap of nine residues between consecutive peptides (Fig. 1A).
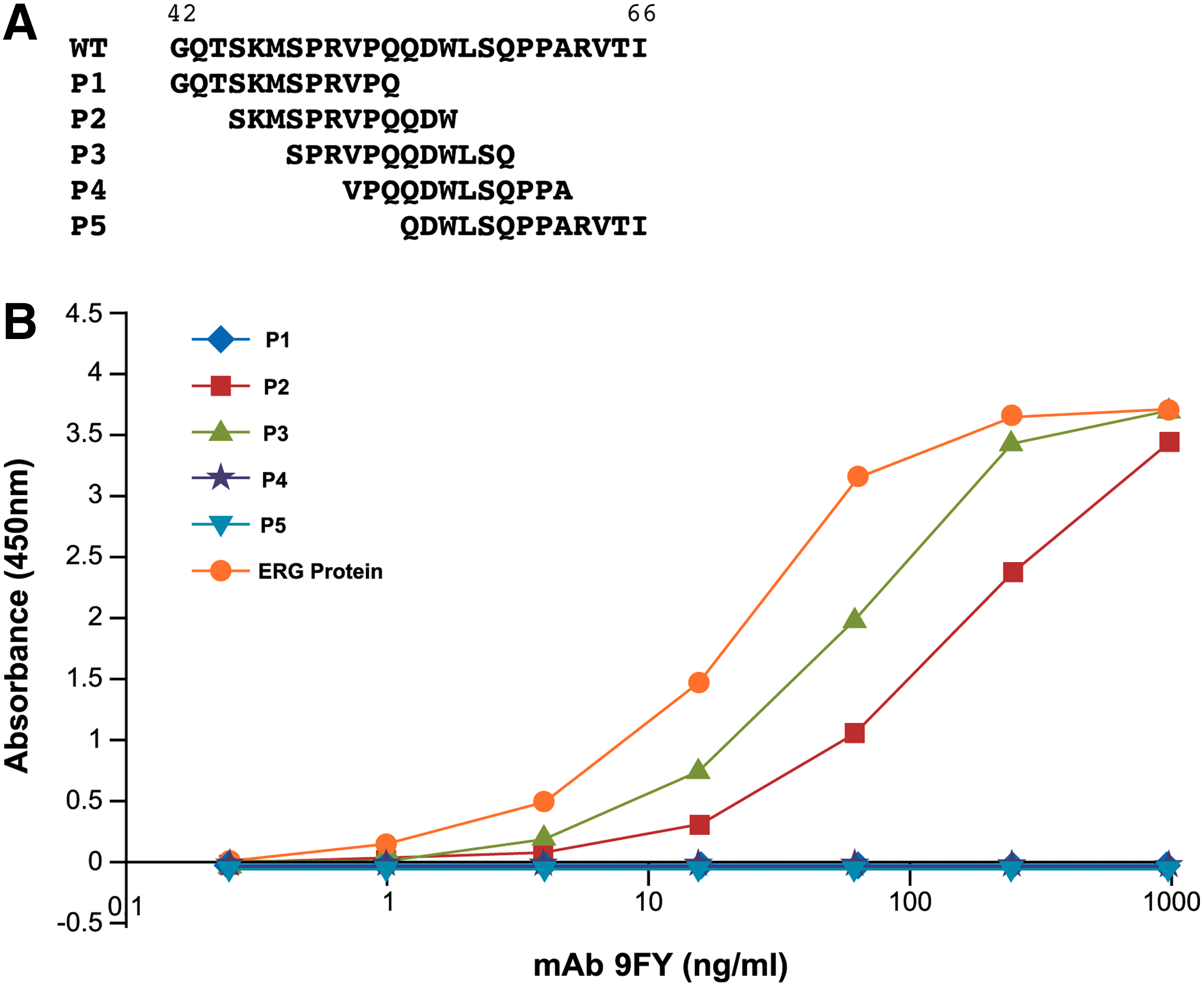
Identification of peptide sequences reactive to 9FY antibody using overlapping peptide approach. (
The peptides were evaluated for their ability to bind ERG MAb 9FY through ELISA. Recombinant ERG protein derived from mammalian cells, used as a positive control, exhibited the highest reactivity to 9FY. Peptide P1 (residues 42-53), representing the N-terminus, failed to bind MAb 9FY, while peptides P2 (residues 45-56) and P3 (residues 48-59) showed strong binding to MAb 9FY. Peptides P4 (residues 51-62) and P5 (residues 54-66) again showed no reactivity to 9FY, similar to peptide P1 (Fig. 1B). These results indicate that the 9FY antibody recognition sequences may be located in a stretch of 15 amino acids, represented by SKMSPR VPQQDWLSQ, starting from residue positions 45-59 of ERG.
Analysis of variant peptides for delineating a minimal epitope for interaction with ERG MAb 9FY
Based on the results from the peptide overlap analysis, variant peptides containing a truncation of residues were synthesized (P6-P22), as shown in Figure 2A. Peptide P6 lacks residues both at the N- and C-termini from peptides P2 and P3. Peptides P7-P22 consisted of amino acids spanning residues 46-59, in which incremental truncations were carried out. These peptides were evaluated for their reactivity to the 9FY antibody by ELISA. While peptide P6 (KMSPRVPQQDW, residues 46-56) showed slightly less binding to 9FY in comparison to P2 and P3, truncated peptide P7 (MSPRVPQQDW) showed a highly reduced binding. Interestingly, peptide P21 (RVPQQDWL) exhibited a high reactivity to 9FY. On the other hand, peptide P22 (PRVPQQDWLS) showed a decreased binding to 9FY (Fig. 2B). The remaining peptides were not reactive against the 9FY antibody. These data suggest that an epitope containing a minimum of eight residues is sufficient for binding to MAb 9FY. This is supported by the lack of reactivity observed with peptide P11 containing seven residues (RVPQQDW) and suggests that residues downstream of W56 may play a role in the binding of the peptide to the antibody.
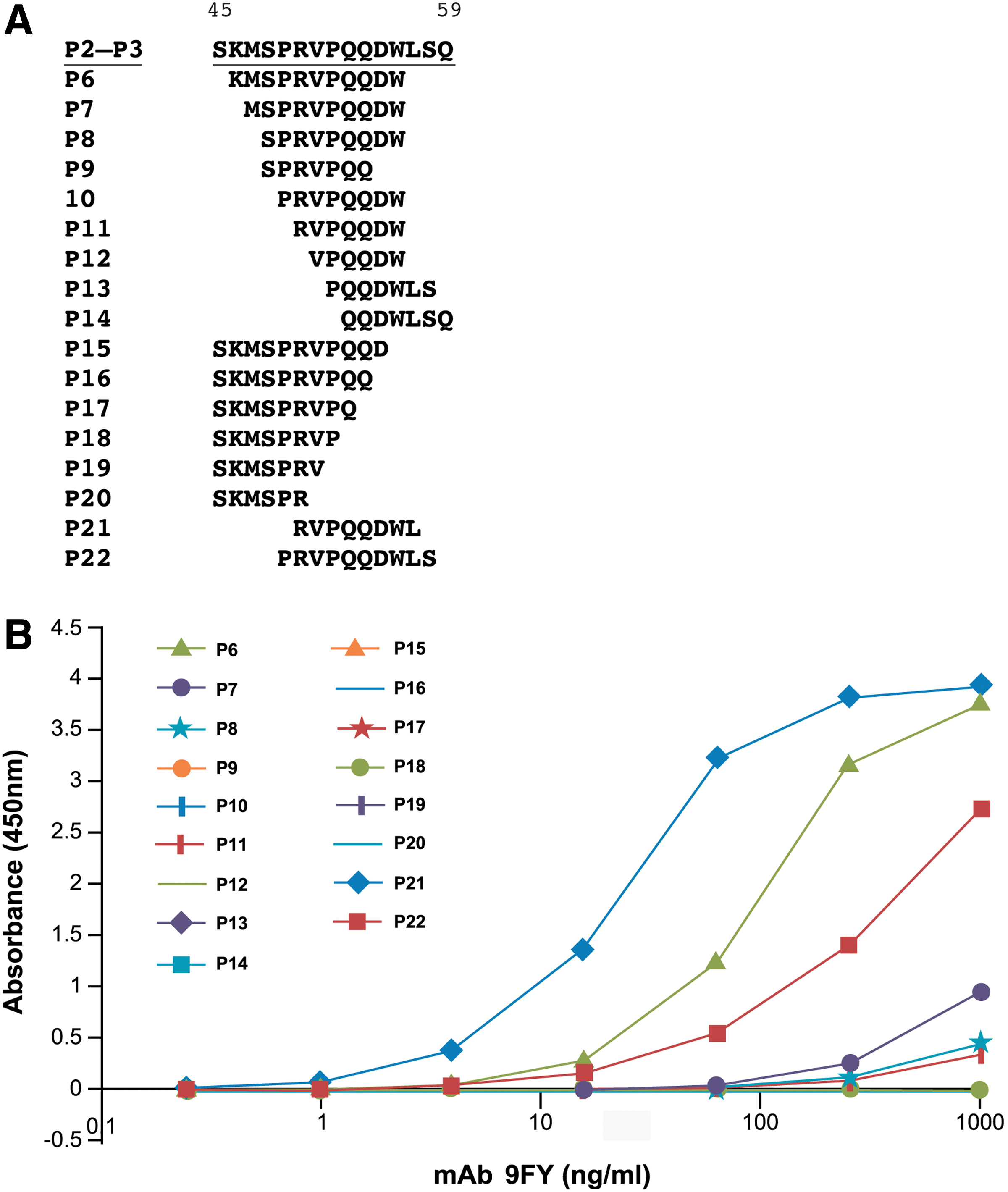
Delineation of a minimal epitope recognized by MAb 9FY. (
Identification of key residues involved in ERG MAb 9FY antibody binding through alanine scanning
In order to analyze the epitope at the level of the residue, we utilized an alanine scanning approach in which each amino acid was individually substituted with an alanine residue. Alanine scanning is routinely used to assess the contribution of side chains of individual amino acids in a protein.(13,14) Peptide P23, which had exhibited high reactivity to 9FY, was used as a template for alanine mutagenesis studies. The peptides used for analyses (P23-P37) are shown in Figure 3A. ELISA was again used for the evaluation of these peptides binding to ERG MAb 9FY. The effect of alanine substitution on the reactivity of peptides with 9FY is shown in Figure 3B. Notably, substitution of alanine individually for residues Q53, Q54, D55, and W56 (corresponding to peptides P30, P29, P28, and P27, respectively) completely eliminated the binding of these peptides to the antibody. Therefore, it is likely that these four residues are part of a core epitope recognized by 9FY. In addition, it was noted that residues flanking the core epitope at the upstream (residue R50, peptide P33) and downstream (residue L57, peptide P26) position showed binding at a reduced level in comparison to P23 containing the wild-type sequence. This suggests that flanking residues may play an accessory role in the binding and stabilization of the peptide-antibody complex.
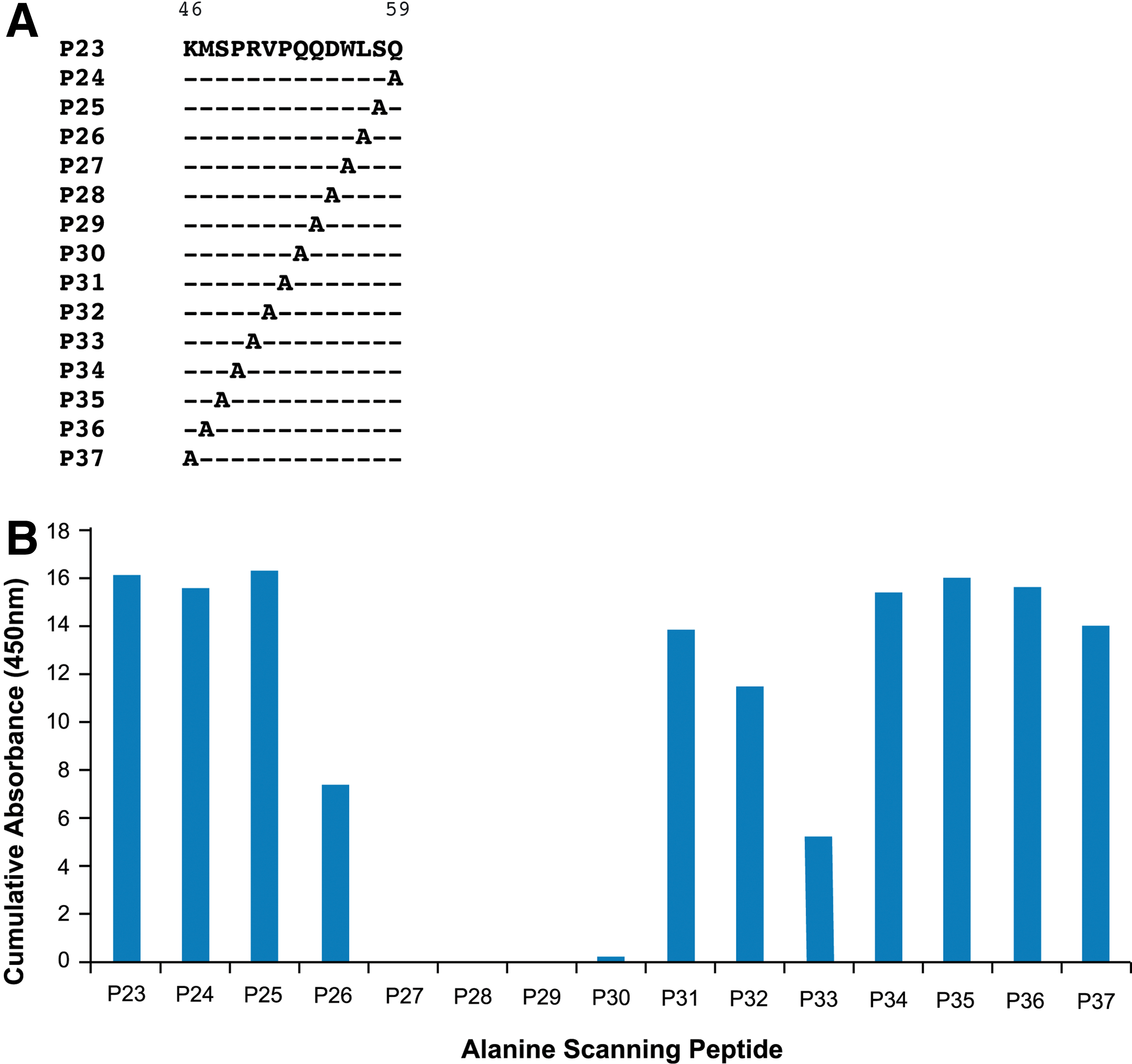
Fine mapping of the residues involved in MAb 9FY binding to peptides. (
Competition studies involving peptides with 9FY antibody binding to ERG protein
Based on the results described above, we tested select peptides for their ability to compete for binding between 9FY and ERG protein. The binding of MAb 9FY to ERG was tested by ELISA using a recombinant ERG protein generated through an expression vector using mammalian cells (Origene, Rockville, MD). Binding was observed in a dose-dependent manner (Fig. 4A). A similar pattern of binding was also observed for the rabbit monoclonal antibody Epitomics-5115 (Epitomics/Abcam, Burlingame, CA), which was raised using a peptide sequence derived from the C-terminus of ERG. In order to assess the specificity of binding, we used a competitive ELISA analysis in which MAb 9FY was pre-incubated with a peptide containing the wild-type epitope sequence (P23) or peptides containing mutated residues (P27 and P30). Peptide P23 competed efficiently for MAb 9FY binding to the ERG protein coated on microtiter wells. Peptides containing an alanine substitution at residue positions Q53 (P30) and W56 (P27) failed to block the binding (Fig. 4B). These results suggest that the epitope recognized by 9FY successfully competed with the binding of 9FY to ERG. The specificity of the epitope was also demonstrated using the Epitomics-5115 rabbit monoclonal antibody against ERG. We reasoned that the peptide recognized by the 9FY epitope is unlikely to be recognized by the Epitomics epitope and therefore may not compete for binding ERG protein. As expected, peptide P23 failed to block the binding of Epitomics-5115 to ERG (Fig. 4C). In addition, we used an immunoblot assay to further evaluate peptide competition. As shown in Figure 4D, peptide P23 effectively interfered with MAb 9FY binding to ERG on the immunoblot. On the other hand, peptides P27 and P30, which did not have reactivity to the 9FY antibody, also failed to prevent the antibody from binding to ERG protein bound to the membrane. These results suggest that peptide P23, which exhibits optimal binding to 9FY antibody, also hinders its reactivity to ERG protein bound to immunoblots by competing for the same site in the antibody.
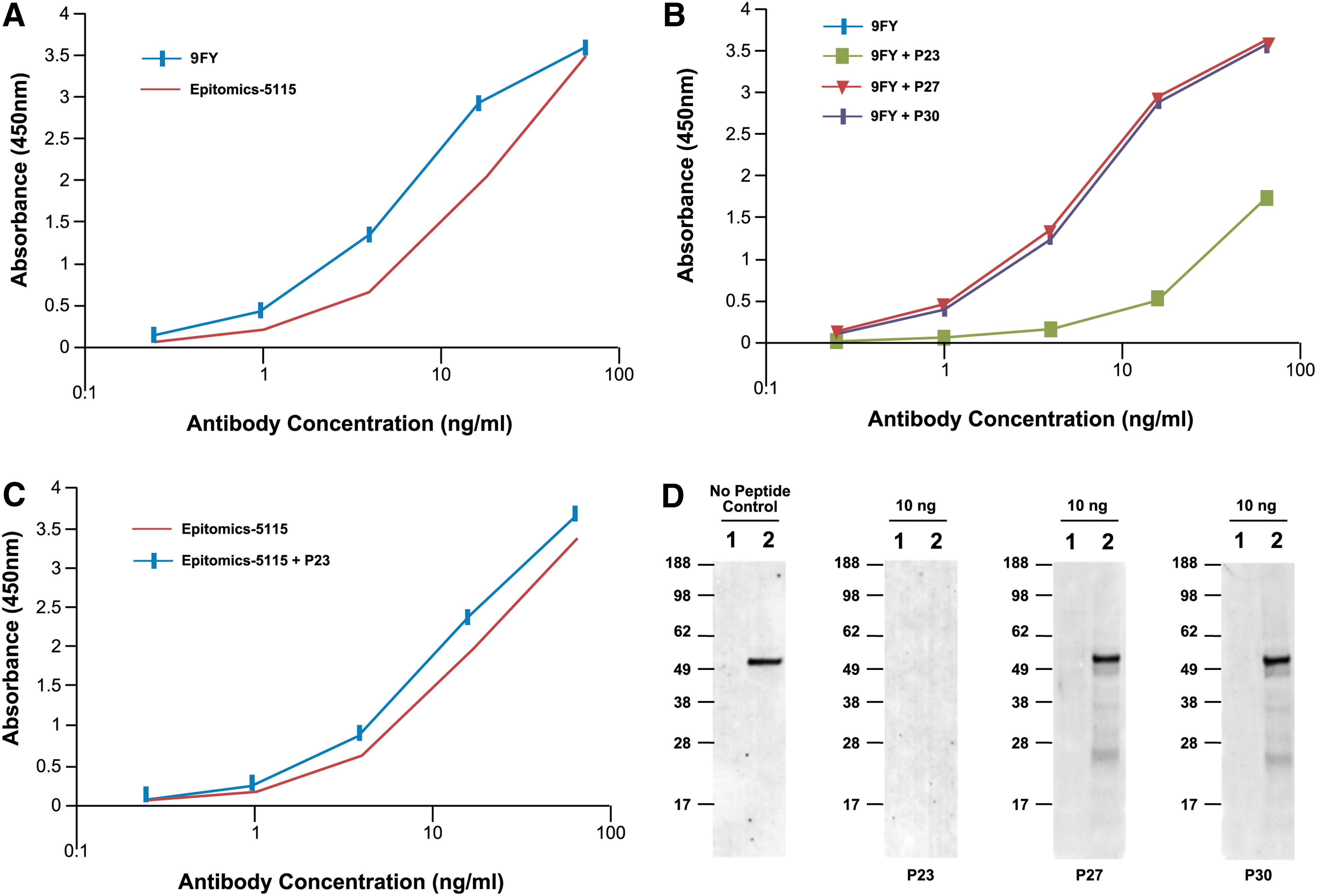
Peptides as competitors to define binding specificity of MAb 9FY to epitope in ERG. (
Discussion
It has been well established that the host immune system plays a vital role in protecting humans from infectious agents and other diseases including cancer. Besides the innate immune system, the body also has an adaptive arm of immunity. The latter comprises humoral and cellular immune responses for protection. This was demonstrated in humans with respect to diverse infectious agents. Specifically, antibodies, elicited in response to proteins from pathogens as part of the humoral response, have been explored as diagnostic markers for identifying individuals with infection, as well as for therapeutic purposes such as passive immunization.(15) This was soon followed by the observation that the immune system also generates antibodies against cancer antigens, which are also known as self antigens. Studies have reported that antibodies to self proteins have been shown to be present in cancers of diverse organs.(16) Based on this, it is suggested that antibodies can also be used to treat cancer. This is further accelerated by the ability to generate monoclonal antibodies against cancer antigens and provides an additional approach in the diagnosis and treatment of cancer.
A major discovery in prostate cancer research is the finding of the intrachromosomal rearrangement resulting in the overexpression of ERG protein.(17,18) The TMPRSS2-ERG gene fusion is present in over half of CA patients. Hence, the detection of ERG expression in the prostate tissues could be used as a surrogate marker for prostate cancer detection. The currently available assays utilize only a few markers for this purpose, using urine (PCA3 and TMPRSS2-ERG, ERG), biopsy tissues (ERG, AMACR, p63 and high molecular weight cytokeratin proteins, TMPRSS2-ERG fusions, and Oncotype DX-prostate) and serum (NADIA ProsVue) as the source for analysis. This served as the basis of our efforts towards the generation of a monoclonal antibody against the ERG protein. The search for immunogenic domains based on antigenicity and hydrophilicity, and further exclusion of homologous regions based on the CLUSTALW alignment of amino acid sequences of ERG and other members of the ETS family, identified an antigenic sequence specific for ERG. The reactivity of MAb 9FY to ERG protein immobilized on a nitrocellulose membrane suggests that 9FY likely recognizes a linear epitope. Interestingly, 9FY also binds to native ERG protein, as has been demonstrated by immunofluorescence and immunoprecipitation assays. This suggests that the epitope in the native ERG protein is likely to be located on the surface, since it is accessible to the 9FY antibody. While there is no structural information available for ERG based on X-ray crystallography, homology modeling studies suggest that the N-terminal region is a disordered domain.(19)
In this study, our goal was to delineate a minimal epitope within amino acids 42-66 of ERG that mediates interaction between the 9FY antibody and the ERG protein. Towards this, we have utilized a pep scan approach in which peptides with an overlap of nine amino acids were generated. Using this method, we showed that peptides P2 and P3 reacted with the 9FY antibody. These results then led us to generate additional peptides (P6-P22) with amino acid truncations to identify a minimal epitope. The comparison of the amino acid sequences of peptides P2 and P3 shows that residues SKM in P2 and LSQ in P3 contribute to the binding of peptides to the antibody. This interpretation is consistent with the data obtained using peptides P6 and P23. The results from an analysis of the variant peptides indicate that a minimal peptide of eight amino acids (P21) in length exhibits high reactivity to MAb 9FY in comparison to a peptide with ten amino acids (P22). It is likely that a reduced reactivity of peptide P22 may be due to the structural/steric features of the peptide. However, a high reactivity noted with P23 indicates that such features are unlikely to exist in the context of a peptide with 14 amino acids.
In addition to identifying the minimal epitope, it was also important to evaluate the contribution of specific residues in the interaction between MAb 9FY and ERG protein. Peptide P23 contains residues K, R, Q, D, W, and L, which have been reported as potential residues for antibody-antigen interactions.(20) To address this, we utilized an alanine scanning approach, as the substitution of alanine would eliminate the side chain at the amino acid level. Peptides P24-P37 containing the alanine substitutions were evaluated by ELISA. The substitution of alanine for Q53, Q54, D55, and W56 individually eliminated the reactivity of the peptide to the 9FY antibody. This suggests that these residues are important for binding and are likely involved in direct interaction with the binding pocket of the antibody. Based on this data, we have designated QQDW as the core residues of the epitope. We can envisage two scenarios regarding a role for the core residues: (1) residues 53-56 individually are involved in the interaction with the antibody, and (2) the contiguous four amino acids provide a binding site for the antibody. Hence, substitution of alanine for any one of the residues leads to the disruption of the binding site. Our experiments also suggest that flanking residues (R50 and L57) on either side of the core residues also play a role, as their alterations also resulted in a reduced level of reactivity to the antibody. It is tempting to suggest that the flanking residues may play a structural role in terms of stabilizing the antigen-antibody complex. Further, the specificity of binding of MAb 9FY to the target protein/peptide was also demonstrated by a competition assay. Competition studies (ELISA and Western blot) showed that peptides corresponding to the epitope efficiently competed with the binding of 9FY to ERG protein immobilized on nitrocellulose or coated on microtiter wells. These results suggest that these peptides share structural determinants or features with native ERG protein.
The identification of a minimal epitope within residues 42-66 of the ERG protein that is recognized by the 9FY antibody is critical for assessing the specificity of the antibody. It should be noted that information about the precise epitope of various commercially available monoclonal antibodies these antibodies is lacking. The immunogens used for generating the antibodies include full-length ERG protein, truncated protein, or synthetic peptides. In addition, the peptide sequence used for generating 9FY (Fig. 5A) was subjected to BLAST analysis. While high homology was noted among all the isoforms of ERG, FLI1 also showed homology in the region comprising the epitope (Fig. 5B). Based on the data from alanine scanning mutagenesis, we suggest that the lack of reactivity of 9FY antibody to FLI1 may be due to the differences in the residues corresponding to R50, D55, and L57 in ERG.

Molecular basis of specificity of MAb 9FY to ERG. (
Conclusions
Overall the data presented here allow us to draw several conclusions: (1) the location of the 9FY binding epitope in ERG indicates that 9FY would recognize a majority of TMPRSS2-ERG variants in the context of prostate cancer; (2) there is no polymorphism reported in the amino acids comprising the epitope; (3) MAb 9FY binds a linear epitope with a minimum of at least 8 amino acids (RVPQQDWL) in the ERG protein; (4) residues Q53, Q54, D55, and W56 are important for recognition by the antibody; (5) the 9FY antibody is highly specific to ERG due to a unique primary amino acid sequence of the epitope. Further studies are needed addressing the therapeutic value of the monoclonal antibodies or modified versions thereof. These attributes qualify 9FY antibody as a valuable reagent in the diagnosis and treatment of prostate cancer.
Footnotes
Acknowledgments
The authors would like to thank Mr. Stephen Doyle for his help with the figures.
Research reported in this publication was partly supported by NCI-EDRN (IAA no. ACN 12011-001-0) and also by the funds awarded to the Center for Prostate Disease Research, a program of the Henry M. Jackson Foundation for the Advancement of Military Medicine Program Fund HU0001-10-2-0002 to D.G.M.
Author Disclosure Statement
The authors have no financial interests to disclose. The opinions or assertions contained herein are the private ones of the author/speaker and are not to be construed as official or reflecting the views of the Department of the Army, Department of Defense, the Uniformed Services University of the Health Sciences, or the U.S. Government.