
Abstract
A panel of six different murine hybridoma clones secreting IgG monoclonal antibodies (MAbs) specific for the human IgM Fc receptor (FcμR) was generated. All MAbs specifically precipitated a major protein of ∼60 kDa from membrane lysates of FcμR-bearing, but not FcμR-negative, cells as did IgM-ligands. Pre-incubation of membrane lysate of FcμR-bearing cells with these MAbs completely removed the ∼60 kDa IgM-reactive protein. By using recombinant human/mouse chimeric FcμR proteins, the epitope recognized by HM7 and HM10 MAbs was mapped to the Ig-like domain of human FcμR, whereas the other MAbs recognized the stalk region. Pre-incubation of FcμR+ cells with the Ig-like domain-specific MAbs, but not with others, markedly inhibited subsequent IgM-ligand binding. A similar, but much weaker, inhibition was also observed when the incubation order was reversed. When FcμR+ cells were simultaneously incubated with both IgM-ligands and MAbs, HM7 MAb efficiently competed with IgM for FcμR binding. Unlike control Jurkat cells, FcμR-bearing cells were resistant to apoptosis induced by agonistic IgM anti-Fas MAb (CH11); however, addition of the HM7 MAb inhibited the interaction of the Fc portion of CH11 MAb with FcμR, thereby promoting apoptosis of FcμR-bearing Jurkat cells. The variable regions of the HM7 MAb were composed of Ighv14-3, Ighd1-2, and Ighj2 for the γ2b heavy chain and Igk3-4 and Igkj2 for the κ light chain. These findings suggest that HM7 MAb efficiently blocks the ligand-binding activity of FcμR.
Introduction
R
In the present study, a panel of six different MAbs against human FcμR has been generated and two MAbs, HM7 and HM10, are described in detail with emphasis on their ability to block the interaction of IgM-ligand with FcμR.
Materials and Methods
FcμR-specific MAbs
Six different hybridoma clones secreting IgG MAbs specific for human FcμR (HM2 [γ3κ]; HM3, HM6, and HM7 [γ2bκ]; HM10 and HM14 [γ1κ]) were developed by hybridization of Ag8.653 plasmacytoma line with regional lymph node cells from BALB/c mice hyper-immunized with the BW5147 mouse thymoma line stably expressing human FcμR. These MAbs were selected based on their restricted reactivity with FcμR-bearing cells but not with pIgR-expressing FT-29, Fcα/μR+, or control BW5147 cells; the characterization of one of the MAbs (HM14) was previously described.(6) The secreted IgG MAbs were purified from the culture supernatants of single cell-derived hybridoma clones that were grown in media containing IgG-depleted fetal bovine sera (FBS) by protein G-coupled affinity columns. Some aliquots of the purified IgG MAbs and highly purified, human myeloma IgM protein were labeled with biotin. Protein concentration was determined by absorbance at 280 nm with an extinction coefficient of 1.4 as 1 mg/mL for both IgG and IgM.
Immunofluorescence analysis
To examine if receptor-specific MAbs block the IgM-ligand binding to FcμR, flow cytometric analysis was performed. Briefly, a mixture of BW5147 cells stably expressing both human FcμR and green fluorescein protein (FcμR+/GFP+) and wild-type control BW5147 cells was sequentially incubated with either anti-FcμR MAb or IgM at 10 μg/mL, then washed and incubated with either biotin-labeled IgM (4 μg/mL, ∼4 nM) or anti-FcμR MAb (1 μg/mL, ∼6 nM), respectively. The bound biotin-Igm or -MAbs were detected by the addition of phycoerythrin-labeled streptavidin (PE-SA, Southern Biotechnology Associates, Birmingham, AL) as previously described.(6) Alternatively, cells were simultaneously incubated with biotin IgM in the presence of various concentrations of anti-FcμR MAbs, washed, and developed with PE-SA. For epitope mapping, cells stably expressing a recombinant human/mouse chimeric FcμR protein (see below) were incubated with receptor-specific MAbs, washed, and developed with PE-labeled goat anti-mouse Ig antibody (Southern Biotechnology Associates). Stained cells were analyzed using an Accuri C6 flow cytometer and flow cytometric data were analyzed with FlowJo software (Tree Star, Ashland, OR).
Cell surface biotinylation and immunoprecipitation analysis
Cell surface proteins were labeled with sulfo-NHS-LC-biotin (Thermo Fisher Scientific, Pierce, Rockford, IL) as previously described.(6) Sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) analysis of immunoprecipitated materials from lysates of biotinylated plasma membrane proteins was essentially the same as that described previously.(6)
Chimeric FcμR
The Ig-like domain of human and mouse FcμRs was swapped by an overlapping polymerase chain reaction (PCR) strategy.(16) In brief, the cDNA corresponding to the Ig-like domain of each FcμR was amplified by PCR using PrimeStar HS DNA polymerase (Takara, Mountain View, CA) and a set of primers: (1) O-481F (5′-gaattcTAGAAGGGACAATGGACTTC-3′) and O-469R (5′-
Apoptosis assay
The apoptosis assay was essentially the same as that described previously.(6) In brief, FcμR+/GFP+ or GFP+ Jurkat cells (5×105 cells/mL) were cultured for 15 to 20 h in the absence or presence of the agonistic IgM anti-Fas MAb (CH11 clone, 10 ng/mL; Millipore, Billerica, MA), along with various concentrations of anti-FcμR MAb (HM7 or HM3). After washing, cells were stained with 7-aminoactinomycin D (7-AAD) and allophycocyanin (APC)-labeled annexin V for detecting apoptotic cells according to the manufacturer's recommendation (BD, Biosciences, San Jose, CA).
Sequencing of Ig heavy and κ light chain variable regions
Nucleotide sequence of variable regions of HM7 MAb was determined by reverse transcription PCR (RT-PCR). Briefly, total RNA was isolated from the HM7 hybridoma clone producing an FcμR-blocking MAb of IgG2bκ isotype and was converted to first-strand cDNA using SuperScript II reverse transcriptase (Invitrogen) and a primer O-454R (5′-ggacagggmtccakagttcc-3′) or O-456R (5′-gttggtgcagcatcagc-3′) corresponding with the constant region of murine γ heavy and κ light chains, respectively. The resultant first-strand cDNAs were used as template DNAs for amplification of cDNA encoding Ig heavy (VH) and light chain (Vκ) variable regions with a set of primers: (1) O-453F (5′-aggtsmarctgcagsagtcwgg-3′, in which S=C or G, M=A or C, R=A or G, and W=A or T)(17) and O-454R for the Ig VH; and (2) O-455F (5′-ccagatgtgtgatgacccagactcca-3′)(18) and O-456R for the Ig Vκ region. The amplified products of the expected size were subcloned into the ZeroBlunt TOPO vector before sequencing analysis and then with the IMGT/V-Quest program.(19)
Results
FcμR-blocking MAbs, HM7, and HM10
A panel of six different MAbs (HM2 [γ3κ isotype], HM3, HM6 and HM7 [γ2bκ], HM10 and HM14 [γ1κ]) was generated from mice hyper-immunized with the mouse thymoma cell line BW5147 stably expressing human FcμR. The hybridomas were selected based on their reactivity by flow cytometric analysis with FcμR+ BW5147 cells and other FcμR-bearing cells (i.e., phorbol myristate acetate-activated 697 pre-B cell line and CLL cells), but not with control or Fcα/μR+ BW5147 cells and pIgR-bearing FT-29 cell line. These MAbs also did not react with myeloid cells or the corresponding cell lines expressing FcγRs, FcɛRI, and/or FcαR, consistent with the results with HM14 MAb.(6) To determine the biochemical nature of the proteins recognized by these MAbs, SDS-PAGE analysis of biotin-labeled cell surface proteins was performed. A major protein with an Mr of ∼60 kDa was specifically precipitated with all MAbs, as well as with IgM-ligands from membrane lysates of FcμR+ BW5147 cells but not from those of control BW5147 cells (Fig. 1A). The sequential immunoprecipitation analysis with MAbs and IgM-ligands revealed that pre-incubation of the membrane lysates of FcμR+ cells with MAbs completely removed the ∼60 kDa IgM-reactive protein, whereas the reverse did not efficiently remove the ∼60 kDa MAb-reactive protein. These findings confirm the FcμR specificity of these MAbs and suggest that these MAbs are better than IgM-ligands for the detection of FcμR.
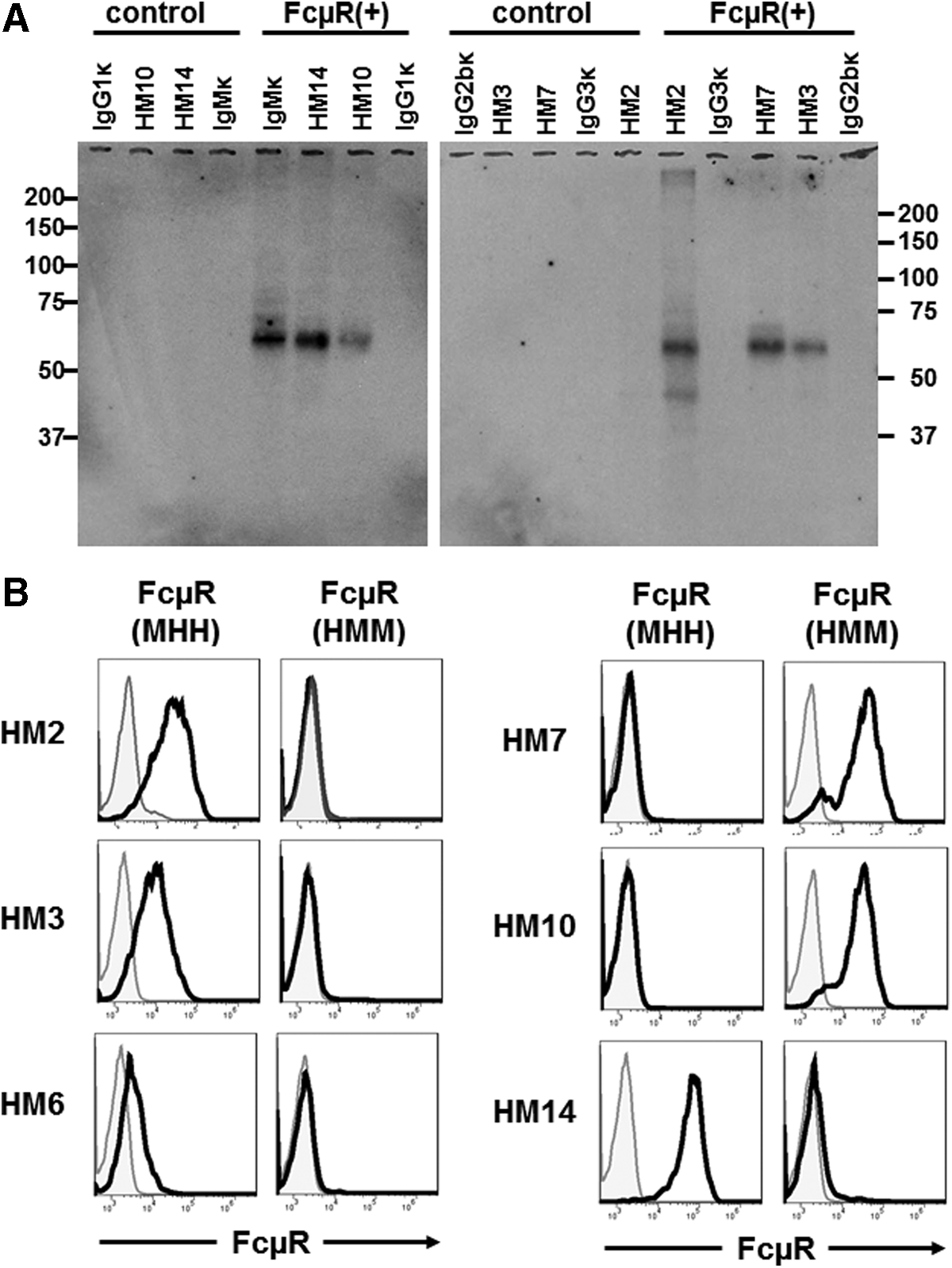
Biochemical characterization of proteins reactive with anti-FcμR MAbs and their epitope mapping. (
To map the epitope recognized by these MAbs, we made recombinant human and mouse FcμR fusion proteins by swapping Ig-like domains, as those MAbs did not cross-react with mouse FcμR. The acronym HMM indicates the FcμR consisting of the Ig-like domain of
To determine if any of these FcμR-specific MAbs can inhibit the interaction of FcμR with IgM-ligands, a mixture of FcμR+/GFP+ and control cells was sequentially incubated with receptor-specific MAbs and then with biotin-labeled IgM (plus PE-SA) before flow cytometric analysis. Pre-incubation with the four stalk region-specific MAbs did not affect the subsequent IgM binding to FcμR+/GFP+ cells, but HM7 and HM10, the two Ig-like domain-specific MAbs, could inhibit the IgM binding almost completely (Fig. 2A). Similar, but much weaker, inhibition by these two MAbs was also observed with the reverse order of incubation—that is, cells were first incubated with IgM and then with biotin-labeled MAbs (Fig. 2B). When cells were simultaneously incubated with ∼4 nM biotin-IgM in the presence of the two-blocking MAbs (5–75 nM), the HM7 MAb efficiently inhibited the binding of IgM to FcμR+/GFP+ cells in a dose-dependent manner, but the HM10 MAb did so only at the highest concentration (Fig. 2C). These findings suggest that two of the FcμR-specific MAbs (HM7 and HM10) recognize an epitope near the site for IgM-ligand binding and that the HM7 MAb efficiently blocks the ligand binding activity of FcμR.
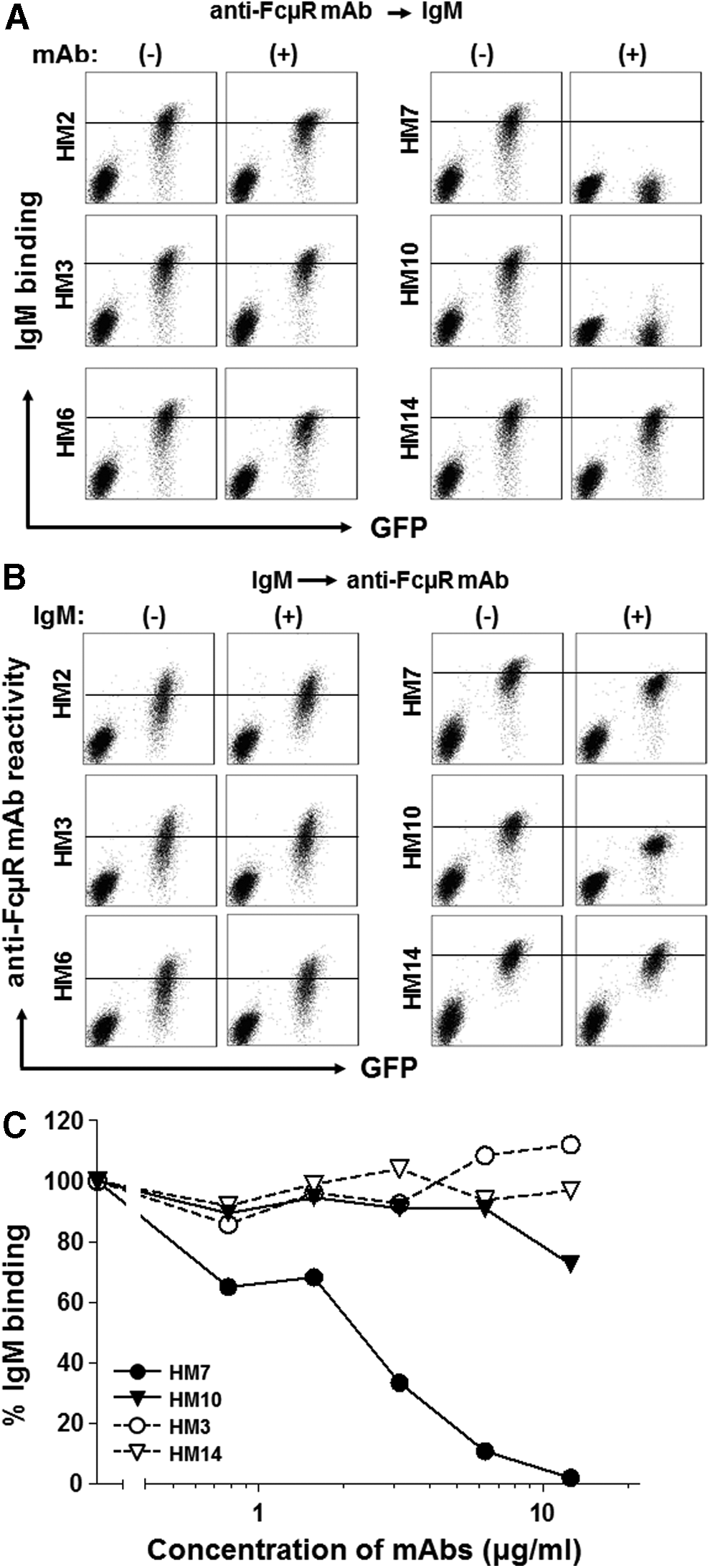
Binding characteristics of FcμR-specific MAbs. Effects of pre-incubation of anti-FcμR MAbs (
HM7 MAb allows FcμR-bearing Jurkat cells to become apoptotic with IgM anti-Fas MAb treatment
In previous studies by our group and others, apoptosis-prone Jurkat cells stably expressing FcμR were shown to be protected from Fas/CD95-mediated apoptosis when ligated with an agonistic IgM anti-Fas MAb (CH11 clone), but not when ligated with an agonistic IgG anti-Fas MAb or Fas ligand.(6,7,20,21) Since this apoptotic inhibition is a well-defined readout of the FcμR ligation, we employed this as a functional assay to determine whether the HM7 MAb, which blocks FcμR ligand binding activity, can inhibit the FcμR-mediated protection from apoptosis induced by the IgM anti-Fas MAb. As expected, the addition of the IgM anti-Fas MAb (10 pM) induced robust apoptosis of control GFP+ Jurkat cells, as determined by early and late apoptotic dyes Annexin V and 7-AAD, respectively (Fig. 3A). By contrast, FcμR+/GFP+ Jurkat cells were protected, consistent with the previous findings.(6,7,20,21) HM7 MAb alone did not induce apoptosis in either of the cell types; however, its inclusion in the assay resulted in an increase in the IgM anti-Fas MAb-mediated apoptosis of FcμR+/GFP+ cells in a dose-dependent manner (Fig. 3B). This observation suggests that the HM7 MAb blocks the binding of the Fc portion of the IgM anti-Fas MAb to FcμR, thus preventing the anti-apoptotic activity of FcμR+ cells in the IgM Fas MAb-mediated apoptosis assay. Addition of the non-blocking, isotype-matched HM3 anti-FcμR MAb did not affect the protection of FcμR+/GFP+ cells from the IgM anti-Fas MAb-induced apoptosis. The IgM anti-Fas MAb-mediated apoptosis of the control vector-only GFP+ cells was not affected by the addition of either HM7 or HM3 MAb. These findings demonstrate that the ability of the HM7 MAb to block FcμR ligand binding activity has functional consequences in that it also blocks the inhibitory activity of FcμR in IgM anti-Fas MAb-induced apoptosis of Jurkat cells.

Effect of HM7 MAb on FcμR-mediated protection of Jurkat cells from IgM anti-Fas MAb-induced apoptosis. (
VH and Vκ sequence of the HM7 MAb
Since the HM7 MAb might be a good candidate for immunotherapeutic purposes, we determined the nucleotide sequence of variable regions of the γ2b heavy and κ light chains by RT-PCR. HM7 was composed of Ighv14-3, Ighd1-2, and Ighj2 for the heavy chain and of Igkv3-4 and Igkj2 for the κ light chain (Fig. 4).

Nucleotide sequence of VH and Vκ regions of HM7 MAb. The nucleotide sequence of VH (upper) and Vκ (lower) regions of HM7 MAb and their translated amino acid sequence are indicated. Upper, Bold letters highlighted in gray, blue, and yellow correspond to the Ighv14-3, Ighd1-2, and Ighj2, respectively, and regular letters indicate the constant region of the γ2b heavy chain. The underlines indicate the sites corresponding to primers O-453F and O-454R. Lower, Bold letters highlighted in gray and yellow correspond to the Igkv3-4 and Igkj2, respectively, and regular letters indicate the constant region of the κ light chain. The underlines indicate the sites corresponding to primers O-455F and O-456R. The sequencing analysis was conducted with three independent clones. The sequences have been deposited in GenBank under the accession nos. KJ865881 for the VH and KJ865882 for the Vκ of HM7 MAb.
Discussion
The present study has focused on the characterization of a panel of six different IgG MAbs reactive with the surface component of human FcμR-bearing cells. All MAbs specifically immunoprecipitated a major protein of ∼60 kDa from membrane lysates of FcμR-bearing cells, but not FcμR-negative cells, as did IgM-ligands. Pre-incubation of membrane lysates of FcμR+ cells with MAbs completely removed the ∼60 kDa IgM-reactive protein. By using recombinant human/mouse chimeric FcμR proteins, the epitope recognized by two MAbs, HM7 (γ2bκ) and HM10 (γ1κ), was mapped to the Ig-like domain, whereas the other MAbs recognized the stalk region of human FcμR. Pre-incubation of FcμR+ cells with Ig-like domain-specific MAbs, but not with the others, markedly inhibited subsequent IgM-ligand binding. A similar, but much weaker, inhibition was also observed in the reverse order of incubation. When cells were simultaneously incubated with both IgM ligands and MAbs, the HM7 MAb efficiently competed the binding of IgM to FcμR-bearing cells. Although FcμR-bearing Jurkat cells, unlike control cells, were resistant to apoptosis induced by the agonistic IgM anti-Fas MAb (CH11), the addition of HM7 MAb inhibited the interaction of the Fc portion of IgM anti-Fas MAb to FcμR, thereby allowing the IgM Fas MAb-mediated apoptosis of FcμR+ cells to occur. The variable regions of the HM7 MAb were defined by cDNA sequencing as VH 14-3, DH 1-2, and JH 2 for the γ2b heavy chain and as Vκ 3-4 and Jκ 2 for the κ light chain. HM7 is thus an efficient blocking MAb for FcμR.
FcμR is the newest member of the FcR family and its cellular distribution is restricted to lymphocytes: B, T, and NK cells in humans(6,7) and B cells only in mice(8–10); hence it is quite distinct from the cellular distribution of FcRs for switched Ig isotypes (i.e., FcγRs, FcɛRs, FcαR). The precise function of FcμR remains elusive but the recent studies of Fcmr-deficient mice have suggested that FcμR plays a regulatory role in IgM homeostasis and humoral immune responses.(9,10,15) The ablation of Fcmr leads to increase in pre-immune serum IgM as well as natural autoantibodies of both IgM and IgG isotypes and to dysregulated antibody responses, especially to suboptimal doses of T cell-independent type 2 antigens. Notably, many abnormalities in Fcmr-deficient mice mirror those observed in mutant mice deficient in IgM secretion, which are able to express surface IgM and other Ig isotypes on B cells and to secrete all other classes of Igs except IgM, suggesting the critical role in normal B cell functions both for secreted IgM and for its interaction with FcμR.(22) Although FCMR deficiency has not yet been identified in humans, it seems likely that the phenotype will be much more complex and profound than the Fcmr deficiency in mice, because human FcμR is expressed by additional cell types, namely T and NK cells.
Enhanced levels of both the ∼60 kDa membrane-bound FcμR and the ∼40 kDa soluble FcμR were shown in patients with CLL compared with healthy donors.(12) Specific delivery of toxin-conjugated IgM Fc to the FcμR is now being explored for potential treatment for CLL patients.(11,23) HM7 and/or other MAbs may be another alternative for this purpose. Intriguingly, it has recently been shown that administration of a recombinant fusion protein consisting of human FcμR ectodomain and IgG Fc as a soluble FcμR ameliorates the myelin oligodendrocyte glycoprotein-induced experimental autoimmune encephalomyelitis in mice, although the mechanism underlying this effect is unclear.(24) In this regard, various forms of immune-mediated peripheral neuropathy are known to be associated with monoclonal IgM antibodies against myelin-associated glycoprotein, gangliosides, or related glycolipids.(25–27) While it is a generally accepted concept that IgM autoantibodies are usually protective but IgG autoantibodies are pathologic in autoimmune disorders, these demyelinating neuropathies are a clear example of disease association with IgM autoantibodies. Another intriguing observation is that patients with Alzheimer's disease have significantly elevated numbers of T cells with autochthonously bound IgM,(28) but whether this cytophilic IgM is bound to T cells via the Fc or Fab region remains to be determined.
Footnotes
Acknowledgments
We would like to thank Ms. Tomoko Motohashi and Mr. Dewitt Jones for their excellent technical assistance, Dr. Peter D. Burrows for critical reading, and Dr. Martin Weigert and Dr. Shozo Izui for suggestions regarding VH and Vκ sequencing. This work was supported in part by the National Institute of Health, National Institute of Allergy and Infectious Diseases (grants R56AI82249 and R21AI94624 to HK).
Author Disclosure Statement
The authors have no financial interests to disclose.